
Sanguinarine Exhibits Antiviral Activity against Porcine Reproductive and Respiratory Syndrome Virus via Multisite Inhibition Mechanisms

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus, and Reagents
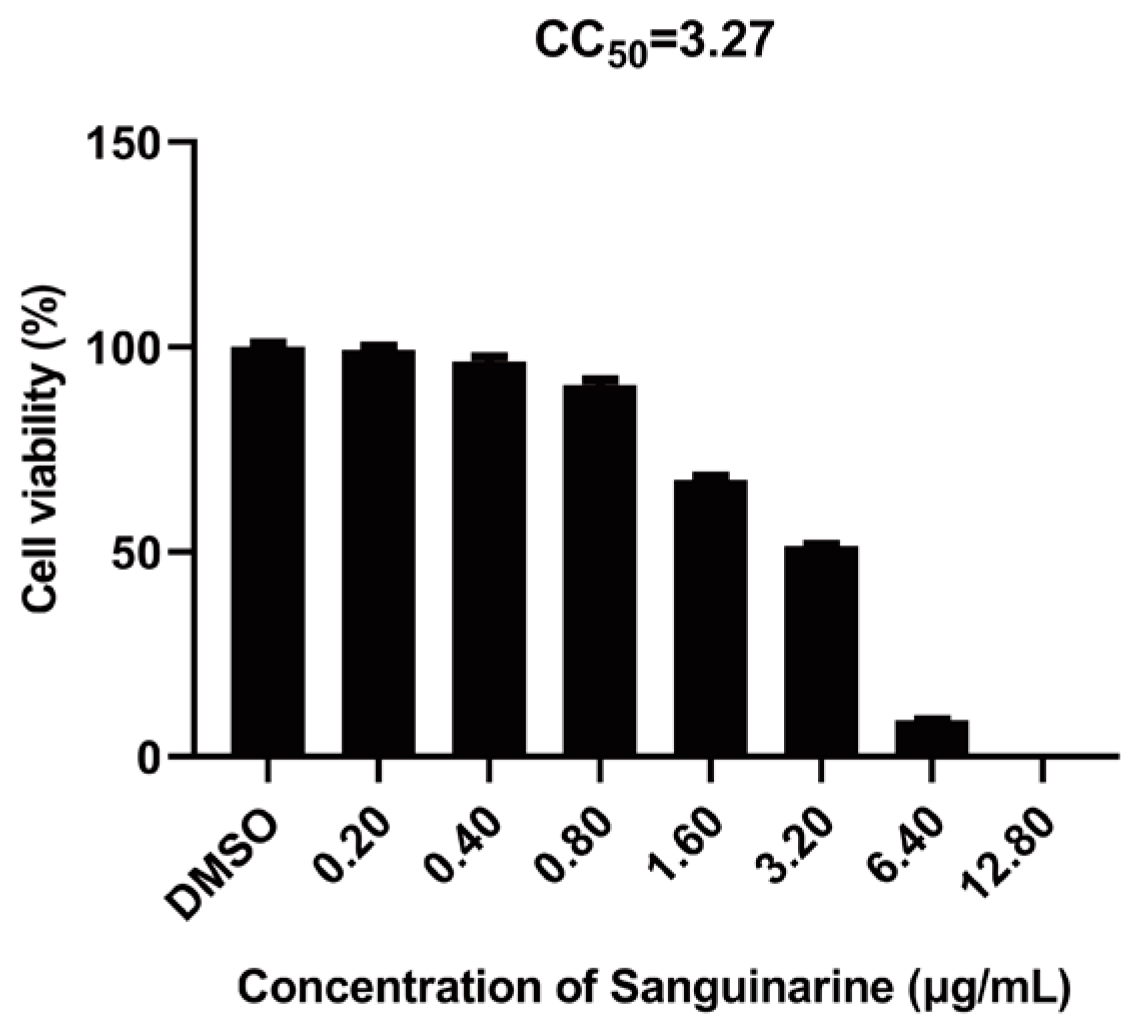
2.2. Cytotoxicity Assay
2.3. The 50% Tissue Culture Infectious Dose (TCID50) Assay
2.4. Western Blot Assay
2.5. RNA Extraction
2.6. Real-Time Reverse-Transcriptase Quantitative PCR (RT-qPCR)
2.7. Immunofluorescence Assay
2.8. Plaque Assay
2.9. Antiviral Assay
2.10. Attachment Assay
2.11. Internalization Assay
2.12. Replication Assay
2.13. Release Assay
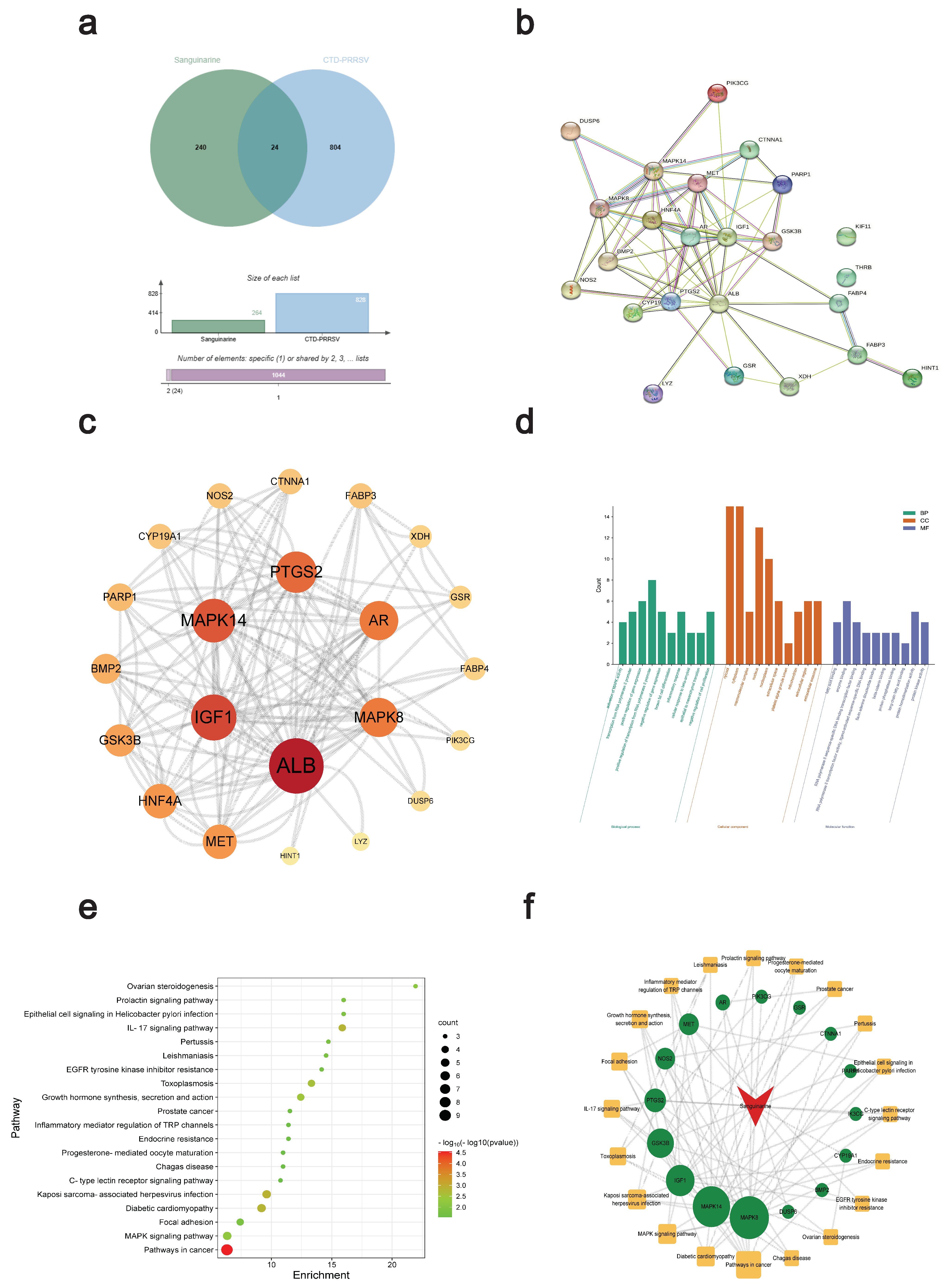
2.14. Anti-PRRSV Target Screening
2.15. PPI Network
2.16. GO and KEGG Pathway Analysis
2.17. Molecular Docking
2.18. Statistical Analysis
3. Results
3.1. Sanguinarine Exhibits Anti-PRRSV Activity
3.2. Sanguinarine Dose-Dependently Inhibits PRRSV Proliferation
3.3. Sanguinarine Restrains PRRSV Infection via Multisite Inhibition Mechanisms
3.4. Network Pharmacology and Molecular Docking Analysis of Potential Anti-PRRSV Targets of Sanguinarine
3.5. Combination with Chelerythrine Improves Antiviral Activity of Sanguinarine
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, Y.F.; Liu, Y.; Yang, J.; Li, W.Z.; Yu, X.X.; Wang, S.Y.; Li, L.A.; Yu, H. Recombination between NADC34-like and QYYZ-like strain of porcine reproductive and respiratory syndrome virus with high pathogenicity for piglets in China. Transbound. Emerg. Dis. 2022, 69, e3202–e3207. [Google Scholar] [CrossRef]
- Montaner-Tarbes, S.; Del Portillo, H.A.; Montoya, M.; Fraile, L. Key Gaps in the Knowledge of the Porcine Respiratory Reproductive Syndrome Virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.J.; Zhou, E.M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Zhang, P.; Zhang, Z.; Youn, J.Y.; Wang, C.; Zhang, H.; Cai, H. Traditional Chinese Medicine (TCM) in the treatment of COVID-19 and other viral infections: Efficacies and mechanisms. Pharmacol. Ther. 2021, 225, 107843. [Google Scholar] [CrossRef]
- Ge, M.; Xiao, Y.; Chen, H.; Luo, F.; Du, G.; Zeng, F. Multiple antiviral approaches of (-)-epigallocatechin-3-gallate (EGCG) against porcine reproductive and respiratory syndrome virus infection in vitro. Antiviral Res. 2018, 158, 52–62. [Google Scholar] [CrossRef]
- Sun, P.; Sun, N.; Yin, W.; Sun, Y.; Fan, K.; Guo, J.; Khan, A.; He, Y.; Li, H. Matrine inhibits IL-1β secretion in primary porcine alveolar macrophages through the MyD88/NF-κB pathway and NLRP3 inflammasome. Vet. Res. 2019, 50, 53. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Zhu, Z.; Wang, X.; Chen, Y.; Liu, X. Pyrithione inhibits porcine reproductive and respiratory syndrome virus replication through interfering with NF-κB and heparanase. Vet. Microbiol. 2017, 201, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Mackraj, I.; Govender, T.; Gathiram, P. Sanguinarine. Cardiovasc. Ther. 2008, 26, 75–83. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Dai, P.; Bao, H.; Liang, P.; Wang, W.; Xing, A.; Sun, J. Anti-inflammatory and neuroprotective effects of sanguinarine following cerebral ischemia in rats. Exp. Ther. Med. 2017, 13, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaziano, R.; Moroni, G.; Buè, C.; Miele, M.T.; Sinibaldi-Vallebona, P.; Pica, F. Antitumor effects of the benzophenanthridine alkaloid sanguinarine: Evidence and perspectives. World J. Gastrointest. Oncol. 2016, 8, 30–39. [Google Scholar] [CrossRef]
- Qing, Z.X.; Cheng, P.; Liu, X.B.; Liu, Y.S.; Zeng, J.G. Systematic identification of alkaloids in Macleaya microcarpa fruits by liquid chromatography tandem mass spectrometry combined with the isoquinoline alkaloids biosynthetic pathway. J. Pharm. Biomed. Anal. 2015, 103, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Lu, X.; Liu, B.; Yan, H.; Feng, J. Anti-TMV activity and mode of action of three alkaloids isolated from Chelidonium majus. Pest Manag. Sci. 2021, 77, 510–517. [Google Scholar] [CrossRef]
- Jena, A.B.; Kanungo, N.; Chainy, G.B.N.; Devaraji, V.; Das, S.K.; Dandapat, J. A Computational Insight on the Inhibitory Potential of 8-Hydroxydihydrosanguinarine (8-HDS), a Pyridone Containing Analog of Sanguinarine, against SARS CoV2. Chem. Biodivers. 2022, 19, e202200266. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fang, L.; Liu, S.; Zhao, F.; Jiang, Y.; He, K.; Chen, H.; Xiao, S. The genomic diversity of Chinese porcine reproductive and respiratory syndrome virus isolates from 1996 to 2009. Vet. Microbiol. 2010, 146, 226–237. [Google Scholar] [CrossRef]
- Huang, P.; Xia, L.; Zhou, L.; Liu, W.; Wang, P.; Qing, Z.; Zeng, J. Influence of different elicitors on BIA production in Macleaya cordata. Sci. Rep. 2021, 11, 619. [Google Scholar] [CrossRef]
- Zhong, M.; Wang, Y.H.; Wang, L.; Long, R.Q.; Chen, C.L. Preparation and application of magnetic molecularly imprinted polymers for the isolation of chelerythrine from Macleaya cordata. J. Sep. Sci. 2018, 41, 3318–3327. [Google Scholar] [CrossRef]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.M. Antiviral Strategies against PRRSV Infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [Green Version]
- Li, W.L.; Zheng, H.C.; Bukuru, J.; De Kimpe, N. Natural medicines used in the traditional Chinese medical system for therapy of diabetes mellitus. J. Ethnopharmacol. 2004, 92, 1–21. [Google Scholar] [CrossRef]
- Bello-Onaghise, G.; Wang, G.; Han, X.; Nsabimana, E.; Cui, W.; Yu, F.; Zhang, Y.; Wang, L.; Li, Z.; Cai, X.; et al. Antiviral Strategies of Chinese Herbal Medicine against PRRSV Infection. Front. Microbiol. 2020, 11, 1756. [Google Scholar] [CrossRef]
- Laines-Hidalgo, J.I.; Muñoz-Sánchez, J.A.; Loza-Müller, L.; Vázquez-Flota, F. An Update of the Sanguinarine and Benzophenanthridine Alkaloids’ Biosynthesis and Their Applications. Molecules 2022, 27, 1378. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, Q.; Chen, Y.; Duan, M.; Tian, G.; Deng, X.; Sun, Y.; Zhou, T.; Zhang, G.; Chen, W.; et al. Inhibition of proanthocyanidin A2 on porcine reproductive and respiratory syndrome virus replication in vitro. PLoS ONE 2018, 13, e0193309. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Lee, C. Sasa quelpaertensis Nakai extract suppresses porcine reproductive and respiratory syndrome virus replication and modulates virus-induced cytokine production. Arch. Virol. 2015, 160, 1977–1988. [Google Scholar] [CrossRef]
- Huang, C.Y.; Huang, Y.J.; Zhang, Z.Y.; Liu, Y.S.; Liu, Z.Y. Metabolism and Tissue Distribution of Chelerythrine and Effects of Macleaya Cordata Extracts on Liver NAD(P)H Quinone Oxidoreductase. Front. Vet. Sci. 2021, 8, 659771. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.X.; Chen, M.; Liu, Y.S.; Shi, Q.; Yang, B.; Zhang, H.C.; Cheng, P.; Tang, Q.; Liu, Z.Y.; Zeng, J.G. Pharmacokinetics of sanguinarine, chelerythrine, and their metabolites in broiler chickens following oral and intravenous administration. J. Vet. Pharmacol. Ther. 2019, 42, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Rawling, M.D.M.; Daniel, L.; Davies Simon, J. Preliminary assessment of dietary supplementation of Sangrovitp on red tilapia (Oreochromis niloticus) growth performance and health. Aquaculture 2009, 294, 1a2. [Google Scholar] [CrossRef]
- Chen, J.; Kang, B.; Zhao, Y.; Yao, K.; Fu, C. Effects of natural dietary supplementation with Macleaya cordata extract containing sanguinarine on growth performance and gut health of early-weaned piglets. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1666–1674. [Google Scholar] [CrossRef]
- Juskiewicz, J.; Gruzauskas, R.; Zdunczyk, Z.; Semaskaite, A.; Jankowski, J.; Totilas, Z.; Jarule, V.; Sasyte, V.; Zdunczyk, P.; Raceviciute-Stupeliene, A.; et al. Effects of dietary addition of Macleaya cordata alkaloid extract on growth performance, caecal indices and breast meat fatty acids profile in male broilers. J. Anim. Physiol. Anim. Nutr. 2011, 95, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M. In vitro Antibacterial Efficacy of Non-Antibiotic Growth Promoters in Poultry Industry. J. Poult. Sci. 2020, 57, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.B.; Chen, B.; Yao, S.Z. Rapid determination of protopine, allocryptopine, sanguinarine and chelerythrine in fruits of Macleaya cordata by microwave-assisted solvent extraction and HPLC-ESI/MS. Phytochem. Anal. 2006, 17, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Liu, Y.C.; Huang, J.L.; Liu, X.B.; Qing, Z.X.; Zeng, J.G.; Liu, Z.Y. Medicinal plants of the genus Macleaya (Macleaya cordata, Macleaya microcarpa): A review of their phytochemistry, pharmacology, and toxicology. Phytother. Res. 2018, 32, 19–48. [Google Scholar] [CrossRef]
- Huigens, R.W., III; Morrison, K.C.; Hicklin, R.W.; Flood, T.A., Jr.; Richter, M.F.; Hergenrother, P.J. A ring-distortion strategy to construct stereochemically complex and structurally diverse compounds from natural products. Nat. Chem. 2013, 5, 195–202. [Google Scholar] [CrossRef]
- Morrison, K.C.; Hergenrother, P.J. Natural products as starting points for the synthesis of complex and diverse compounds. Nat. Prod. Rep. 2014, 31, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasker, S.Z.; Cowfer, A.E.; Hergenrother, P.J. Preparation of Structurally Diverse Compounds from the Natural Product Lycorine. Org. Lett. 2018, 20, 5894–5898. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| nsp9 | ACCCTAGGACCTGTGAAC | GGCGAGTAACTTAGGAGATG |
| ORF7 | GCAATTGTGTCTGTCGTC | CTTATCCTCCCTGAATCTGAC |
| 5′UTR | GCATTTGTATTGTCAGGAGC | AGCAGTGCAACTCCGGAAG |
| 5′UF | GACGTATAGGTGTTGGCTC |
| Names of Target Genes |
|---|
| BMP2, AR, ALB, CYP19A1, LYZ, GSR, KIF11, DUSP6, NOS2, PTGS2, CTNNA1, THRB, MAPK14, HNF4A, MET, MAPK8, PARP1, GSK3B, IGF1, PIK3CG, XDH, HINT1, FABP4, FABP3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, Q.; Duan, K.; Cheng, Y.; Xu, S.; Xiao, S.; Fang, L. Sanguinarine Exhibits Antiviral Activity against Porcine Reproductive and Respiratory Syndrome Virus via Multisite Inhibition Mechanisms. Viruses 2023, 15, 688. https://doi.org/10.3390/v15030688
Ke Q, Duan K, Cheng Y, Xu S, Xiao S, Fang L. Sanguinarine Exhibits Antiviral Activity against Porcine Reproductive and Respiratory Syndrome Virus via Multisite Inhibition Mechanisms. Viruses. 2023; 15(3):688. https://doi.org/10.3390/v15030688
Chicago/Turabian StyleKe, Qiyun, Kaiqi Duan, Yan Cheng, Si Xu, Shaobo Xiao, and Liurong Fang. 2023. "Sanguinarine Exhibits Antiviral Activity against Porcine Reproductive and Respiratory Syndrome Virus via Multisite Inhibition Mechanisms" Viruses 15, no. 3: 688. https://doi.org/10.3390/v15030688