

Incidence and Epidemiology of Citrus Viroids in Greece: Role of Host and Cultivar in Epidemiological Characteristics

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Surveys, Citrus Hosts, Cultivars and Plant Tissue Sampling
2.2. Viroid Detection
2.3. Cloning, Sequence Analysis and Variability
2.4. Data Analyses and Processing
3. Results
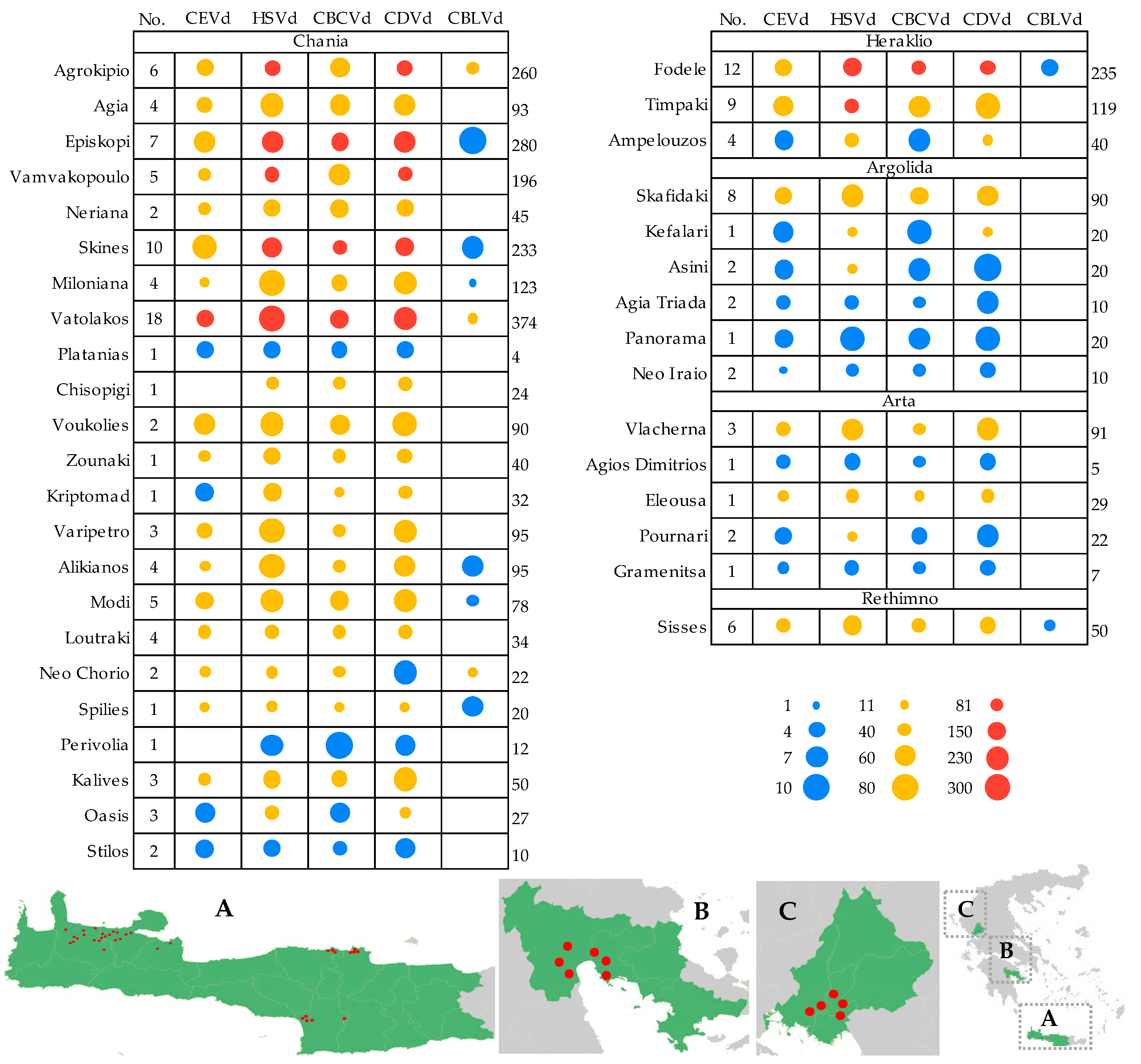
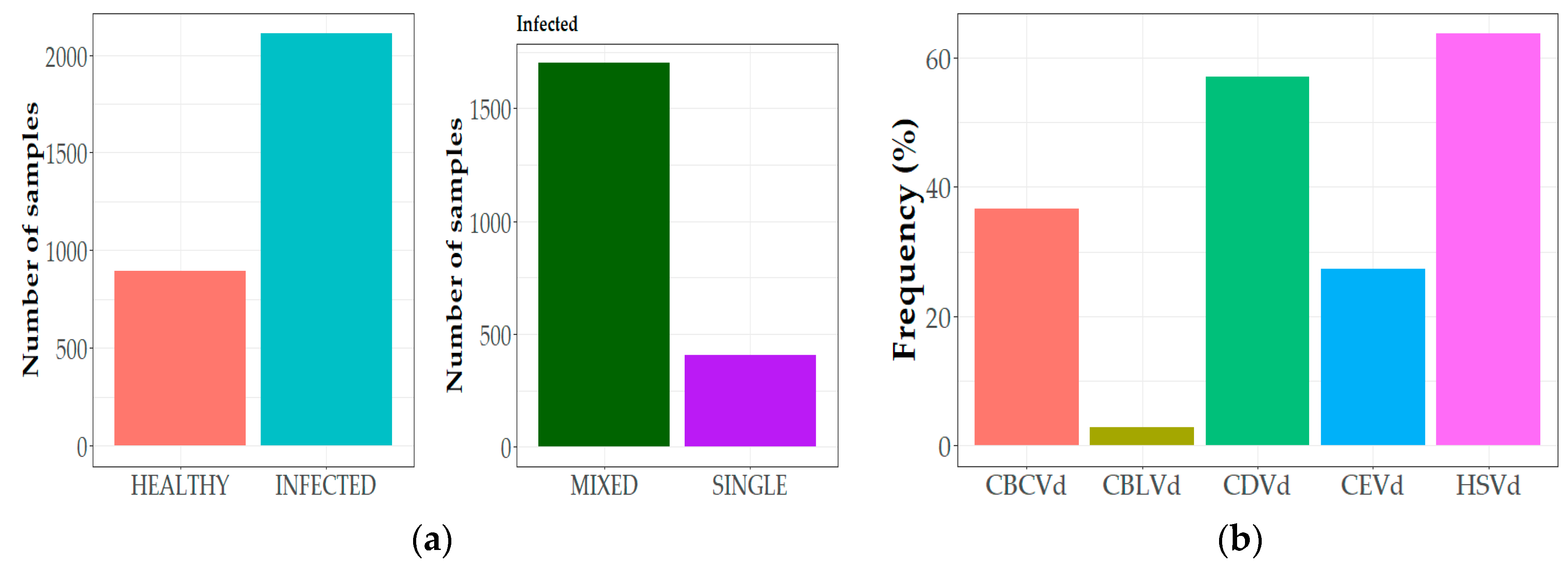
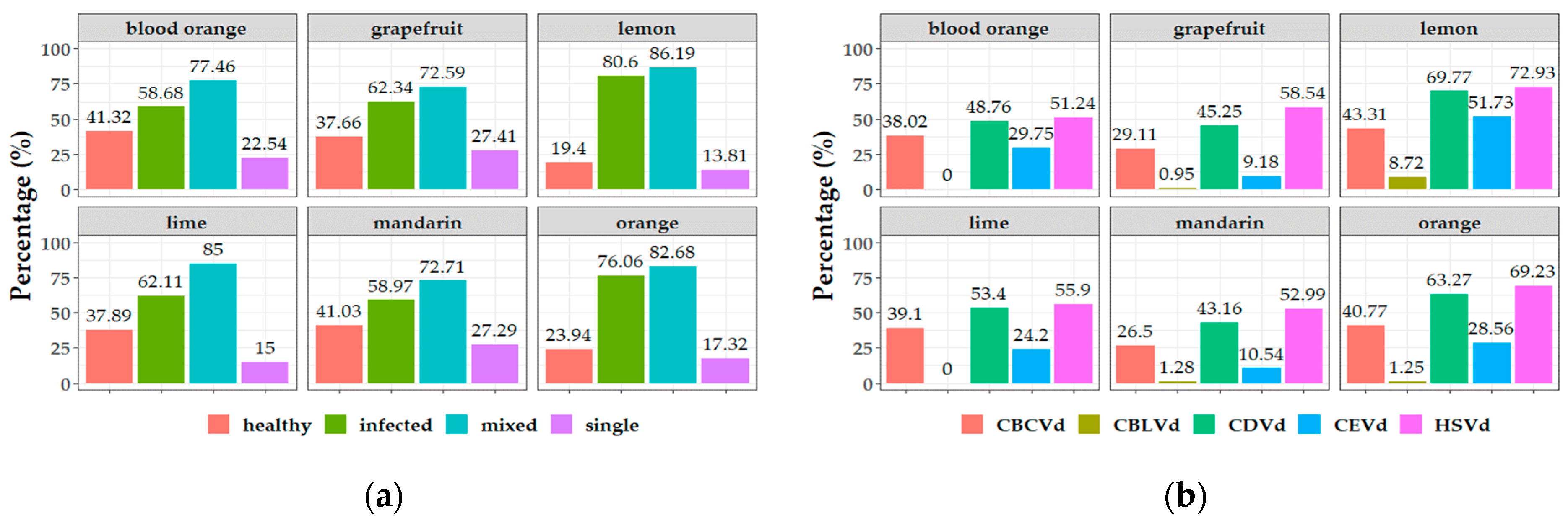
3.1. Incidence and Distribution of Citrus Viroids in Greece
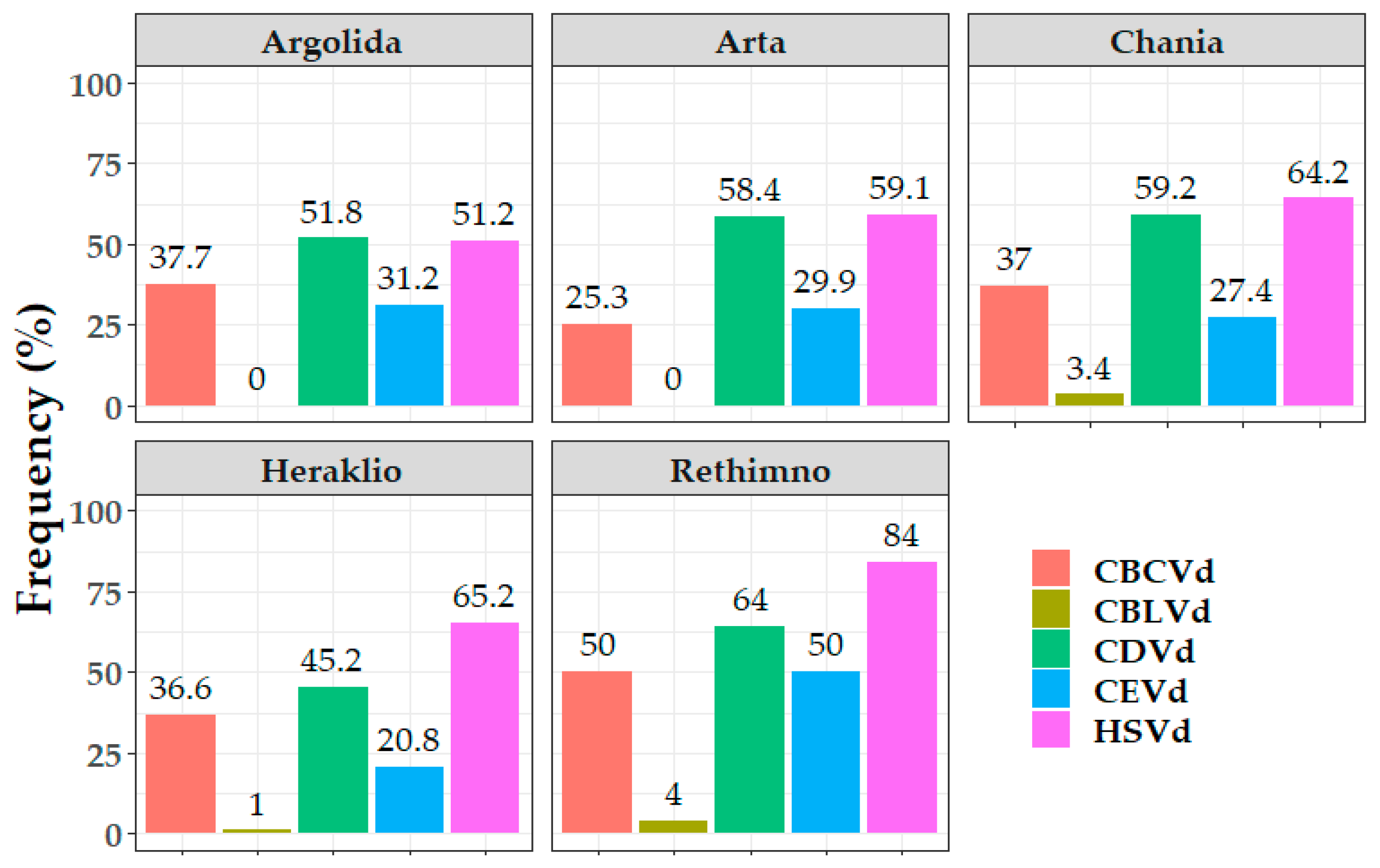
3.2. Epidemiological Characteristics of Citrus Viroids
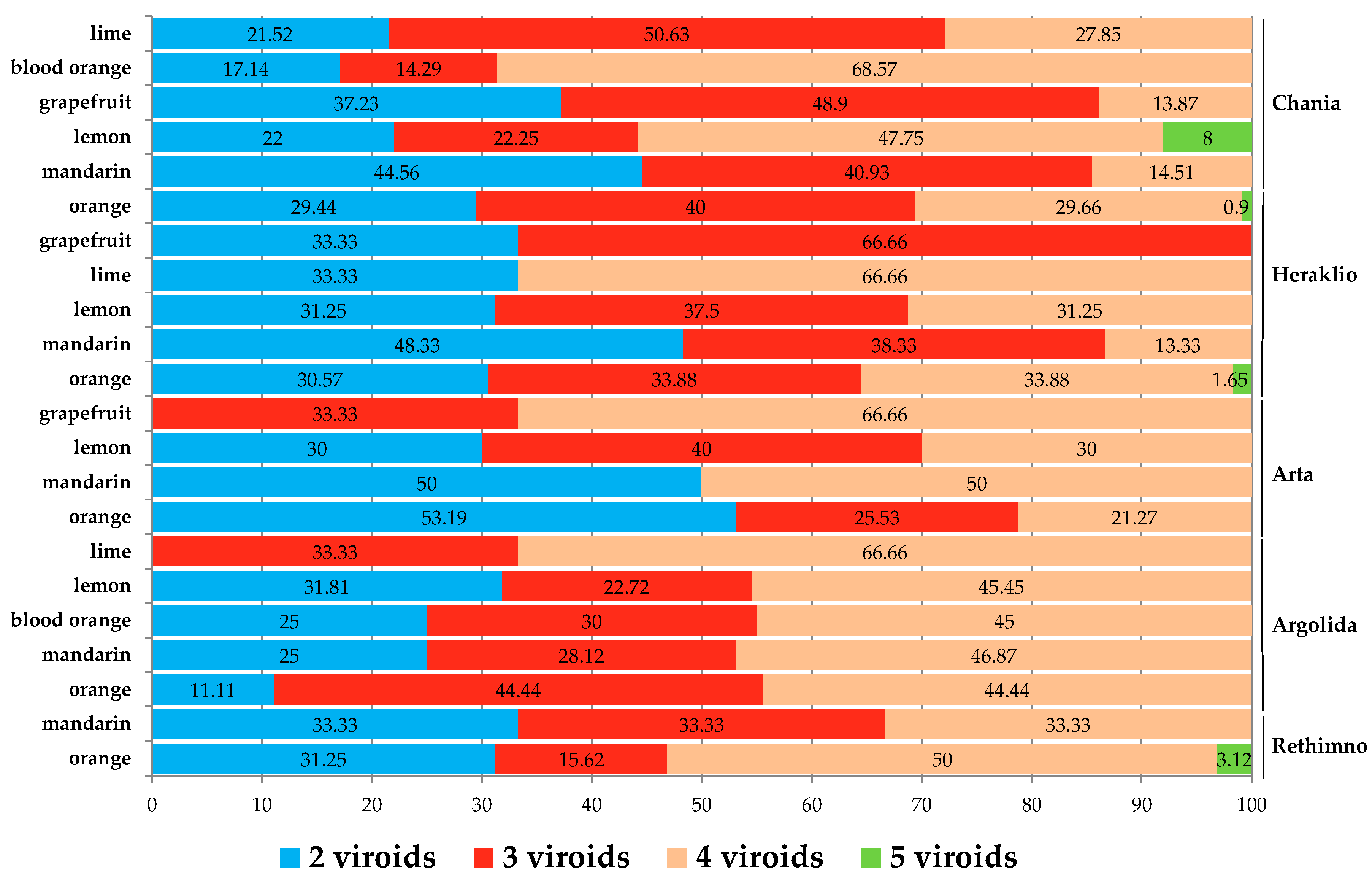
3.3. Potential Correlation between Pathogen Viroid and Host—Cultivar
3.4. Sequence Analysis of Viroid Isolates, Symptoms Description
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. FAOSTAT Database. 2020. Available online: https://www.fao.org/faostat/en/#data/FBS (accessed on 15 December 2022).
- Tennant, P.F.; Robinson, D.; Fisher, L.; Bennett, S.M.; Hutton, D.; Coates-Beckford, P.; McLaughlin, W. Diseases and pests of citrus (Citrus spp.). Tree For. Sci. Biotech. 2009, 3, 81–107. [Google Scholar]
- ELSTAT. 2019. Available online: http://www.statistics.gr/el/statistics/agr (accessed on 15 December 2022).
- Moreno, P.; Ambrós, S.; Albiach-Martí, M.R.; Guerri, J.; Peña, L. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant Pathol. 2008, 9, 251–268. [Google Scholar] [CrossRef]
- Dimou, D.; Drossopoulou, J.; Moschos, E.; Varveri, C.; Bem, F. First Report of Citrus tristeza virus in Greece. Plant Dis. 2002, 86, 329. [Google Scholar] [CrossRef]
- Malandraki, I.; Marouli, E.; Varveri, C. New isolates of Citrus tristeza virus naturally occurring in old lemon and mandarin trees in Greece. New Dis. Rep. 2011, 23, 2. [Google Scholar] [CrossRef] [Green Version]
- Owen, C.; Mathioudakis, M.; Gazivoda, A.; Gal, P.; Nol, N.; Kalliampakou, K.; Figas, A.; Bellan, A.; Iparaguirre, A.; Rubio, L.; et al. Evolution and Molecular Epidemiology of Citrus tristeza virus on Crete: Recent Introduction of a Severe Strain. J. Phytopathol. 2014, 162, 839–843. [Google Scholar] [CrossRef]
- Chatzivassiliou, E.K.; Nolasco, G. Detection of a new variant of Citrus tristeza virus in Greek citrus crops. Phytopathol. Mediterr. 2014, 53, 140–147. [Google Scholar] [CrossRef]
- Chambers, G.A.; Donovan, N.J.; Bodaghi, S.; Jelinek, S.M.; Vidalakis, G. A novel citrus viroid found in Australia, tentatively named citrus viroid VII. Arch. Virol. 2018, 163, 215–218. [Google Scholar] [CrossRef]
- Duran-Vila, N.; Roistacher, C.N.; Rivera-Bustamante, R.; Semancik, J.S. A Definition of Citrus Viroid Groups and Their Relationship to the Exocortis Disease. J. Gen. Virol. 1988, 69, 3069–3080. [Google Scholar] [CrossRef]
- Ito, T.; Ieki, H.; Ozaki, K.; Ito, T. Characterization of a new citrus viroid species tentatively termed Citrus viroid OS. Arch. Virol. 2001, 146, 975–982. [Google Scholar] [CrossRef]
- Serra, P.; Barbosa, C.; Daròs, J.; Flores, R.; Duran-Vila, N. Citrus viroid V: Molecular characterization and synergistic interactions with other members of the genus Apscaviroid. Virology 2008, 370, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Hadidi, A.; Randles, J.; Flores, R.; Palukaitis, P. (Eds.) Viroids and Satellites; Elsevier: Cambridge, MA, USA, 2017. [Google Scholar]
- Wang, J.; Boubourakas, I.N.; Voloudakis, A.E.; Agorastou, T.; Magripis, G.; Rucker, T.L.; Kyriakopoulou, P.E.; Vidalakis, G. Identification and characterization of known and novel viroid variants in the Greek national citrus germplasm collection: Threats to the industry. Eur. J. Plant Pathol. 2013, 137, 17–27. [Google Scholar] [CrossRef]
- Katsarou, K.; Chiumenti, M.; Kalantidis, K.; Mathioudakis, M.M. First Report of Citrus Viroids Infecting Persian (Tahiti) Lime in Greece. Plant Dis. 2020, 104, 998. [Google Scholar] [CrossRef]
- Mathioudakis, M.M.; Saponari, M.; Hasiów-Jaroszewska, B.; Elbeaino, T.; Koubouris, G. The Detection of viruses in olive cultivars in Greece, using a rapid and effective RNA extraction method, for certification of virus-tested propagation material. Phytopathol. Mediterr. 2020, 59, 203–211. [Google Scholar] [CrossRef]
- Ito, T.; Ieki, H.; Ozaki, K. Simultaneous detection of six citrus viroids and Apple stem grooving virus from citrus plants by multiplex reverse transcription polymerase chain reaction. J. Virol. Methods 2002, 106, 235–239. [Google Scholar] [CrossRef]
- Bernad, L.; Duran-Vila, N. A novel RT-PCR approach for detection and characterization of citrus viroids. Mol. Cell. Probes 2006, 20, 110–113. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- RStudio Team Rstudio. Integrated Development for R. Rstudio; PBC: Boston, MA, USA, 2020. [Google Scholar]
- Flores, R.; Randles, J.W.; Palukaitis, P. Viroids and Satellites. Part VI. Geographical Distribution of Viroids and Viroid Diseases; Academic Press: Boston, MA, USA, 2017; pp. 459–518. [Google Scholar]
- Lavagi, I.; Matoušek, J.; Vidalakis, G. Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Other Cocadviroids; Academic Press: Cambridge, MA, USA, 2017; pp. 275–287. [Google Scholar]
- Zhou, C.; da Graça, J.V.; Freitas-Astúa, J.; Vidalakis, G.; Duran-Vila, N.; Lavagi, I. The Genus Citrus; Citrus Viruses and Viroids; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Lin, C.-Y.; Wu, M.-L.; Shen, T.-L.; Yeh, H.-H.; Hung, T.-H. Multiplex detection, distribution, and genetic diversity of Hop stunt viroid and Citrus exocortis viroid infecting citrus in Taiwan. Virol. J. 2015, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-F.; Zhou, C.-Y.; Tang, K.-Z.; Lan, J.-Q.; Zhou, Y.; Li, Z.-A. Preliminary Studies on Species and Distribution of Citrus Viroids in China. Agric. Sci. China 2008, 7, 1097–1103. [Google Scholar] [CrossRef]
- Najar, A.; Hamdi, I.; Varsani, A.; Duran-Vila, N. Citrus Viroids in Tunisia: Prevalence and molecular characterization. J. Plant Pathol. 2017, 99, 787–792. [Google Scholar] [CrossRef]
- Ito, T.; Namba, N.; Ito, T. Distribution of citrus viroids and Apple stem grooving virus on citrus trees in Japan using multiplex reverse transcription polymerase chain reaction. J. General Plant Pathol. 2003, 69, 205–207. [Google Scholar]
- Malfitano, M.; Barone, M.; Duran-Vila, N.; Alioto, D. Indexing of viroids in citrus orchards of Campania, Southern Italy. J. Plant Pathol. 2005, 87, 115–121. [Google Scholar]
- Gillings; Broadbent, P.; Gollnow, B. Viroids in Australian Citrus: Relationship to Exocortis, Cachexia and Citrus Dwarfing. Funct. Plant Biol. 1991, 18, 559–570. [Google Scholar] [CrossRef]
- Villalobos, W.; Rivera, C.; Hammond, R.W. Occurrence of citrus viroids in Costa Rica. Rev. De Biol. Trop. 1997, 45, 983–987. [Google Scholar]
- Aviña-Padilla, K.; Zamora-Macorra, E.J.; Ochoa-Martínez, D.L.; Alcántar-Aguirre, F.C.; Hernández-Rosales, M.; Calderón-Zamora, L.; Hammond, R.W. Mexico: A Landscape of Viroid Origin and Epidemiological Relevance of Endemic Species. Cells 2022, 11, 3487. [Google Scholar] [CrossRef]
- Donovan, N.J.; Chambers, G.A.; Englezou, A.; Phanthavong, S.; Daly, A.; Wildman, O.; Holford, P.; Burgess, L.W. First report of citrus exocortis viroid, citrus bent leaf viroid, hop stunt viroid and citrus dwarfing viroid in Lao PDR. Australas. Plant Pathol. 2020, 49, 661–663. [Google Scholar] [CrossRef]
- Pagliano, G.; Umaña, R.; Pritsch, C.; Rivas, F.; Duran-Vila, N. Occurrence, prevalence and distribution of citrus viroids in Uruguay. J. Plant Pathol. 2013, 95, 631–635. [Google Scholar] [CrossRef]
- Murcia, N.; Bernad, L.; Caicedo, A.; Duran-Vila, N. Citrus viroids in Colombia. In Proceedings of the Seventeenth IOCV Conference, Adana, Turkey, 22–26 October 2007; UC Riverside: Riverside, CA, USA, 2010; pp. 158–166. [Google Scholar]
- Mohamed, M.E.; Hashemian, S.B.; Dafalla, G.; Bové, J.M.; Duran-Vila, N. Occurrence and identification of citrus viroids from Sudan. J. Plant Pathol. 2009, 91, 185–190. [Google Scholar]
- Al-Shariqi, R.; Al-Hammadi, M.; Al-Sadi, A. First report of Citrus bent leaf viroid in the United Arab Emirates. J. Plant Pathol. 2014, 95, 4–71. [Google Scholar] [CrossRef]
- Khoo, Y.; Iftikhar, Y.; Murugan, T.; Roslin, N.; Adawiyah, R.; Kong, L.; Vadamalai, G. First Report of Citrus Bent Leaf Viroid in Malaysia. J. Plant Pathol. 2017, 99, 293. [Google Scholar] [CrossRef]
- Mazhar, M.A.; Bagherian, S.A.A.; Ardakani, A.S.; Izadpanah, K. Nucleotide Sequence and Structural Features of Hop Stunt Viroid and Citrus Bent Leaf Viroid Variants from Blighted Citrus Plants in Kohgiluyeh–Boyerahmad Province of Iran. J. Agr. Sci. Tech. 2014, 16, 657–665. [Google Scholar]
- Cao, M.J.; Atta, S.; Liu, Y.Q.; Wang, X.F.; Zhou, C.Y.; Mustafa, A.; Iftikhar, Y. First report of Citrus bent leaf viroid and Citrus dwarfing viroid from citrus in Punjab, Pakistan. Plant Dis. 2009, 93, 840. [Google Scholar] [CrossRef]
- Iftikhar, Y.; Khoo, Y.W.; Murugan, T.; Roslin, N.A.; Adawiyah, R.; Kong, L.L.; Vadamalai, G. Molecular and biological characterization of citrus bent leaf viroid from Malaysia. Mol. Biol. Rep. 2021, 49, 1581–1586. [Google Scholar] [CrossRef]
- Wu, Q.; Cao, M.; Su, H.; Atta, S.; Yang, F.; Wang, X.; Zhou, C. Molecular characterization and phylogenetic analysis of Citrus viroid I-LSS variants from citrus in Pakistan and China reveals their possible geographic origin. Eur. J. Plant Pathol. 2014, 139, 13–17. [Google Scholar] [CrossRef]
- Najar, A.; Hamdi, I.; Ben Mahmoud, K. Citrus viroids: Characterization, prevalence, distribution and struggle methods. J. New Sci. 2018, 50, 3129–3137. [Google Scholar]
- Bernad, L.; Gandía, M.; Duran-Vila, N. Host effect on the genetic variability of Citrus exocortis viroid (CEVd). In Proceedings of the Sixteenth IOCV Conference, Monterrey, NL, Mexico, 7–12 November 2004; UC Riverside: Riverside, CA, USA, 2005; pp. 291–300. [Google Scholar]
- Gandía, M.; Bernad, L.; Rubio, L.; Duran-Vila, N. Host Effect on the Molecular and Biological Properties of a Citrus exocortis viroid Isolate from Vicia faba. Phytopathology 2007, 97, 1004–1010. [Google Scholar] [CrossRef] [Green Version]
- Szychowski, J.A.; Vidalakis, G.; Semancik, J.S. Host-directed processing of Citrus exocortis viroid. J. Gen. Virol. 2005, 86, 473–477. [Google Scholar] [CrossRef]
- Bernard, L.; Duran-Vila, N.; Elena, S. Effect of citrus hosts on the generation, maintenance and evolutionary fate of genetic variability of citrus exocortis viroid. J. Gen. Virol. 2009, 90, 2040–2049. [Google Scholar] [CrossRef] [Green Version]
- Tessitori, M.; Rizza, S.; Reina, A.; Causarano, G.; Di Serio, F. The genetic diversity of Citrus dwarfing viroid populations is mainly dependent on the infected host species. J. Gen. Virol. 2013, 94, 687–693. [Google Scholar] [CrossRef]
- Cheng, X.-F.; Wu, X.-Y.; Wang, H.-Z.; Sun, Y.-Q.; Qian, Y.-S.; Luo, L. High codon adaptation in citrus tristeza virus to its citrus host. Virol. J. 2012, 9, 113. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Leal, R.; Bryan, B.K.; Rush, C.M. Host Effect on the Genetic Diversification of Beet necrotic yellow vein virus Single-Plant Populations. Phytopathology 2010, 100, 1204–1212. [Google Scholar] [CrossRef] [Green Version]
- Mathioudakis, M.; Maliogka, V.; Candresse, T.; Nickel, O.; Fajardo, T.; Budzyńska, D.; Hasiów-Jaroszewska, B.; Katis, N. Molecular Characterization of the Coat Protein Gene of Greek Apple Stem Pitting Virus Isolates: Evolution through Deletions, Insertions, and Recombination Events. Plants 2021, 10, 917. [Google Scholar] [CrossRef]
- De Moya-Ruiz, C.; Rabadán, P.; Juárez, M.; Gómez, P. Assessment of the Current Status of Potyviruses in Watermelon and Pumpkin Crops in Spain: Epidemiological Impact of Cultivated Plants and Mixed Infections. Plants 2021, 10, 138. [Google Scholar] [CrossRef]
- Malpica, J.M.; Sacristán, S.; Fraile, A.; García-Arenal, F. Association and Host Selectivity in Multi-Host Pathogens. PLoS ONE 2006, 1, e41. [Google Scholar] [CrossRef]
- Belabess, Z.; Radouane, N.; Sagouti, T.; Tahiri, A.; Lahlali, R. A Current Overview of Two Viroids Prevailing in Citrus Orchards: Citrus Exocortis Viroid and Hop Stunt Viroid. In Citrus; Khan, M.S., Ahmad, I., Eds.; Taschen: Cologne, Germany, 2021; p. 27. [Google Scholar]
- Nagy, P.D. Plant Virus Evolution; Roossinck, M.J., Ed.; Recombination in Plant RNA Viruses; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Jakse, J.; Radisek, S.; Pokorn, T.; Matousek, J.; Javornik, B. Deep-sequencing revealed Citrus bark cracking viroid (CBCVd) as a highly aggressive pathogen on hop. Plant Pathol. 2015, 64, 831–842. [Google Scholar] [CrossRef]
- Hataya, T.; Nakahara, K.; Ohara, T.; Ieki, H.; Kano, T. Citrus viroid Ia is a derivative of citrus bent leaf viroid (CVd-Ib) by partial sequence duplications in the right terminal region. Arch. Virol. 1998, 143, 971–980. [Google Scholar] [CrossRef]
- Wang, Y.; Atta, S.; Wang, X.; Yang, F.; Zhou, C.; Cao, M. Transcriptome sequencing reveals novel Citrus bark cracking viroid (CBCVd) variants from citrus and their molecular characterization. PLoS ONE 2018, 13, e0198022. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHANIA | CEVd | HSVd | CBCVd | CDVd | CBLVd | ARTA | CEVd | HSVd | CBCVd | CDVd | CBLVd | HERAKLIO | CEVd | HSVd | CBCVd | CDVd | CBLVd |
| Valencia | 18.18 | 66.06 | 38.18 | 60.61 | 3.03 | Lane Late | 10 | 70 | 20 | 40 | 0 | Valencia | 0 | 55 | 35 | 30 | 0 |
| Chalder | 22.5 | 67.5 | 25 | 72.5 | 0 | W. Navel | 28.57 | 42.86 | 28.57 | 42.86 | 0 | New Hall | 18 | 76 | 24 | 44 | 0 |
| Lane Late | 30.16 | 65.87 | 36.51 | 61.9 | 0 | Mpotsato | 12.5 | 67.5 | 10 | 75 | 0 | W. Navel | 20 | 53.33 | 43.33 | 43.33 | 6.67 |
| New Hall | 33 | 72 | 45 | 72 | 0 | Saloustiana | 52.63 | 63.16 | 52.63 | 68.42 | 0 | Mirodato | 36.96 | 71.74 | 52.17 | 61.96 | 0 |
| W. Navel | 29.81 | 78.26 | 49.69 | 74.53 | 0 | Adamopoulou | 60 | 80 | 40 | 80 | 0 | Soultani | 55 | 75 | 50 | 60 | 0 |
| N. Late | 40 | 67.5 | 42.5 | 60 | 10 | Nuvel Athous | 51.72 | 68.97 | 48.28 | 72.41 | 0 | Zampetaki | 46.67 | 86.67 | 33.33 | 53.33 | 0 |
| Xino Chanion | 34.55 | 67.27 | 47.27 | 63.64 | 0 | Red | 60 | 60 | 40 | 60 | 0 | Adamopoulou | 42.86 | 71.43 | 0 | 42.86 | 0 |
| Zampetaki | 42.86 | 69.29 | 35.71 | 67.14 | 7.14 | Nova | 0 | 0 | 0 | 33.33 | 0 | Eureka | 75 | 50 | 50 | 50 | 0 |
| Adamopoulou | 61.07 | 74.81 | 55.73 | 74.81 | 9.16 | Clementine | 35 | 65 | 15 | 50 | 0 | White | 0 | 80 | 60 | 40 | 0 |
| Vakalou | 65.69 | 81.75 | 55.47 | 78.1 | 20.44 | Moro | 0 | 28.57 | 0 | 0 | 0 | Ancor | 0 | 70.97 | 45.16 | 71.94 | 6.45 |
| Interdonato | 32.5 | 71.25 | 23.75 | 68.75 | 10 | Persian | 0 | 0 | 0 | 0 | 0 | Clementine | 16 | 52 | 28 | 32 | 0 |
| Eureka | 51.95 | 72.73 | 44.16 | 67.53 | 0 | Kino | 3.3 | 53.85 | 23.08 | 31.87 | 0 | ||||||
| Nova | 0 | 31 | 6 | 42 | 0 | ARGOLIDA | CEVd | HSVd | CBCVd | CDVd | CBLVd | Persian | 50 | 75 | 50 | 75 | 0 |
| Ancor | 7.95 | 50.99 | 18.54 | 40.4 | 4.64 | Valencia | 20 | 60 | 60 | 40 | 0 | ||||||
| Clementine | 23.88 | 56.72 | 43.28 | 49.25 | 0 | Lane Late | 20 | 20 | 0 | 60 | 0 | RETHIMNO | CEVd | HSVd | CBCVd | CDVd | CBLVd |
| Kino | 0 | 55.74 | 52.46 | 54.1 | 0 | New Hall | 0 | 0 | 0 | 0 | 0 | New Hall | 53.57 | 85.71 | 57.14 | 67.86 | 7.14 |
| Page | 12.5 | 67.5 | 21.25 | 43.75 | 1.53 | W. Navel | 30 | 60 | 50 | 70 | 0 | W. Navel | 50 | 83.33 | 41.67 | 75 | 0 |
| White | 8.33 | 45.83 | 13.33 | 36.67 | 0 | Interdonato | 42.5 | 52.5 | 32.5 | 50 | 0 | Nova | 40 | 80 | 40 | 40 | 0 |
| Red | 11.76 | 77.21 | 51.47 | 63.24 | 2.21 | Tarocco | 30 | 60 | 50 | 70 | 0 | ||||||
| Pink | 0 | 36 | 2 | 16 | 0 | Moro | 25 | 60 | 60 | 60 | 0 | ||||||
| Tarocco | 25.81 | 51.61 | 25.81 | 43.89 | 0 | Nova | 50 | 52.5 | 35 | 50 | 0 | ||||||
| Moro | 37.04 | 48.15 | 51.85 | 44.44 | 0 | Clementine | 50 | 55 | 40 | 60 | 0 | ||||||
| Kara Kara | 43.75 | 75 | 56.25 | 75 | 0 | Page | 20 | 40 | 35 | 40 | 0 | ||||||
| Persian | 23.97 | 56.85 | 39.73 | 54.79 | 0 | Persian | 40 | 80 | 60 | 60 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathioudakis, M.M.; Tektonidis, N.; Karagianni, A.; Mikalef, L.; Gómez, P.; Hasiów-Jaroszewska, B. Incidence and Epidemiology of Citrus Viroids in Greece: Role of Host and Cultivar in Epidemiological Characteristics. Viruses 2023, 15, 605. https://doi.org/10.3390/v15030605
Mathioudakis MM, Tektonidis N, Karagianni A, Mikalef L, Gómez P, Hasiów-Jaroszewska B. Incidence and Epidemiology of Citrus Viroids in Greece: Role of Host and Cultivar in Epidemiological Characteristics. Viruses. 2023; 15(3):605. https://doi.org/10.3390/v15030605
Chicago/Turabian StyleMathioudakis, Matthaios M., Nikolaos Tektonidis, Antonia Karagianni, Louiza Mikalef, Pedro Gómez, and Beata Hasiów-Jaroszewska. 2023. "Incidence and Epidemiology of Citrus Viroids in Greece: Role of Host and Cultivar in Epidemiological Characteristics" Viruses 15, no. 3: 605. https://doi.org/10.3390/v15030605