
Human Coronavirus OC43 as a Low-Risk Model to Study COVID-19

College of Pharmacy, Dongguk University-Seoul, Goyang 10326, Republic of Korea
*
Author to whom correspondence should be addressed.
Viruses 2023, 15(2), 578; https://doi.org/10.3390/v15020578
Submission received: 18 January 2023
/
Revised: 8 February 2023
/
Accepted: 16 February 2023
/
Published: 20 February 2023
(This article belongs to the Special Issue SARS-CoV-2 and Other Coronaviruses)
Abstract
:The coronavirus disease 2019 (COVID-19) pandemic has had irreversible and devastating impacts on every aspect of human life. To better prepare for the next similar pandemic, a clear understanding of coronavirus biology is a prerequisite. Nevertheless, the high-risk nature of the causative agent of COVID-19, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), requires the use of a cumbersome biosafety level-3 (BSL-3) confinement facility. To facilitate the development of preventive and therapeutic measures against SARS-CoV-2, one of the endemic strains of low-risk coronaviruses has gained attention as a useful research alternative: human coronavirus OC43 (HCoV-OC43). In this review, its history, classification, and clinical manifestations are first summarized. The characteristics of its viral genomes, genes, and evolution process are then further explained. In addition, the host factors necessary to support the life cycle of HCoV-OC43 and the innate, as well as adaptive, immunological responses to HCoV-OC43 infection are discussed. Finally, the development of in vitro and in vivo systems to study HCoV-OC43 and its application to the discovery of potential antivirals for COVID-19 by using HCoV-OC43 models are also presented. This review should serve as a concise guide for those who wish to use HCoV-OC43 to study coronaviruses in a low-risk research setting.
1. Introduction
Despite the successful development and active administration of several commercially available vaccines and antivirals, the suffering inflicted by the coronavirus disease 2019 (COVID-19) pandemic seems likely to continue for the foreseeable future. The emergence of more contagious variants of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is further aggravating the already-devasted healthcare systems around the world. The classification of SARS-CoV-2 as a biosafety level 3 (BSL-3) pathogen due to its relatively high fatality rate has restricted its accessibility to the scientific community for research purposes. This has been one of the biggest hurdles to overcome in the prompt development of effective vaccines and therapeutics against COVID-19. In this regard, human coronavirus OC43 (HCoV-OC43) has gained new attention as an attractive and safe model to study coronaviruses in the low-risk research setting (BSL-2). To better understand its utility as an alternative to SARS-CoV-2 research, the history and classification of HCoV-OC43 together with the clinical manifestations associated with its infection are first summarized. The characteristics of its viral genomes and genes are also presented in comparison with SARS-CoV-2. Then, its evolution process in the context of interspecies and intraspecies perspectives is further explained. The host–virus interaction sections cover the host factors necessary to support the viral life cycle of HCoV-OC43, followed by a description of the innate—as well as adaptive—immunological responses to HCoV-OC43 infection. The development and application of in vitro and in vivo models to study HCoV-OC43 and the discovery of potential antivirals for COVID-19 by using HCoV-OC43 models are also discussed.
2. Classification of Human Coronaviruses
In general, all coronaviruses are members of the Coronaviridae family of the order Nidovirales. Mammals including bats, cows, mice, birds, and humans serve as their natural hosts [1]. Based on sequence homology, they are further classified into four genera including alpha, beta, gamma, and delta coronaviruses. Endemic strains of human coronavirus such as 229E and NL63 are typical examples of alpha coronaviruses. The genus beta coronavirus is further divided into 2a (OC43 and HKU1), 2b (SARS), and 2c (middle east respiratory syndrome, MERS) coronaviruses [2,3,4]. Animal viruses such as mouse hepatitis virus (MHV) and bovine coronavirus (BCoV) also belong to the genus beta coronavirus. HCoV-229E, HCoV-NL63, HCoV-HKU1, and HCoV-OC43 are four typical co-circulating low-risk human coronaviruses. They are predominantly associated with mild infection of the upper respiratory tract [5]. They have become endemic respiratory coronaviruses since their introduction into the human population [2]. Due to their zoonotic origin, all these endemic strains are thought to have broken the interspecies barrier, resulting in altered tropism for humans [6]. With regard to the zoonosis of SARS-CoV-2, the origin of SARS-CoV-2 infection in the wild was proposed to begin from bats based on sequencing results of bat coronaviruses [7,8]. In the wild, reptiles, horses, camels, sheep, llamas, marine mammals, and badgers were suspected to be infected with SARS-CoV-2 with no experimental evidence [9]. Although some cases of SARS-CoV-2 infection in pigs, birds, and cows were reported, they all seem to be inconclusive or lack experimental evidence [9]. Interestingly, the transmission of SARS-CoV-2 from human to animals including cats, dogs, tigers, lions, and mink has also been confirmed [9]. As experimental models to study SARS-CoV-2, mice, hamsters, and ferrets are generally used for this purpose [9,10].
3. Brief History of HCoV-OC43
As explained previously, HCoV-OC43 is one of the seasonally circulating community-acquired coronaviruses, together with other three endemic strains. HCoV-OC43 was first discovered by using a human embryonic tracheal organ culture (OC) technique [11,12]. An examination of nasopharyngeal washings from an acute respiratory tract disease patient led to the recovery of multiple strains of infectious agents. Their sensitivity to ether indicated the requirement of a lipid-containing protein coat for productive infection [11,12]. Among them, two of the six strains, designated OC38 and OC43, were able to cause encephalitis when inoculated into newborn mice, highlighting their distinct neurotrophic pathogenesis despite their respiratory origin [13]. Due to the morphological resemblance to avian infectious bronchitis virus (IBV), HCoV-OC43 has initially been named an “IBV-like” virus [11,14]. A hemagglutination inhibition test was frequently utilized as a preferred method for its characterization [15]. In addition, owing to its close structural similarity to MHV, an adaptation of OC43 to the suckling-mouse brain was empirically tested and successfully utilized for its efficient propagation [12,13].
4. Clinical Manifestations of HCoV-OC43 Infection
In general, infection with human coronaviruses is responsible for common colds. Some patients show symptoms of diarrhea and enterocolitis. Roughly, 15 to 35% of all common colds are estimated to be attributed to coronavirus infection [12,16,17]. On occasion, they have been associated with neurologic disorders such as multiple sclerosis [18]. According to surveillance data, HCoV-OC43 has been regarded as the most frequently found strain responsible for coronavirus-associated mild colds [19]. In particular, among all respiratory viruses identified in respiratory infection samples, 4.7% turned out to be related to HCoV-OC43 [20]. Along with HCoV-229E and HCoV-NL63, HCoV-OC43 has also been shown to be a primary etiological agent for mild upper respiratory tract infections in children [12,20,21,22]. In line with this, a higher rate of infections with HCoV-OC43 has been consistently reported [23,24,25,26,27]. Although HCoV-OC43 infections are generally mild, subclinical, and self-limited [28], more severe respiratory tract infections including bronchiolitis and pneumonia have been reported in high-risk groups. These are infants, elderly individuals, and immunocompromised patients [29,30,31]. Despite the potential involvement of HCoV-OC43 in neurologic diseases, its association with neurotrophic etiology has never been validated [32,33]. The main symptoms of HCoV-OC43 infection are similar to respiratory tract infection symptoms like fever and cough [20]. Infection with other respiratory viruses has also been suggested as one of the contributing factors to the development of lower respiratory tract infections by HCoV-OC43 [20].
Unlike common community-acquired endemic human coronaviruses, recently emerged novel coronaviruses such as SARS-CoV-1, SARS-CoV-2, and MERS-CoV can cause a highly severe clinical course of infection and fatality mainly in elderly patients, particularly those with previous chronic medical conditions [34]. However, given the gradually decreased clinical severity shown in recently reported variants, SARS-CoV-2 seems to be in the process of establishing itself as one of these endemic human respiratory coronaviruses [2]. In addition, several reports suggest a protective role of pre-existing cross-immunity against endemic human coronaviruses in SARS-CoV-2 patients [35]. Therefore, understanding the mutual relationship between high- and low-risk coronaviruses will be critical not only for the development of effective countermeasures against current pandemics but also for the correct prediction of the evolutionary direction of current high-risk coronaviruses in the future.
5. Genome Structure of HCoV-OC43
Structurally, the single-stranded RNA genome of coronavirus is encapsidated in a helical nucleocapsid [28]. In general, the initial portion of the coronavirus genome encodes two long open reading frames (ORF1a and 1b), which generate sixteen non-structural (NS) proteins necessary for viral replication. Four structural proteins and accessory proteins (NS2a and NS5a) are translated from their separate subgenomic RNAs. Viral structural proteins include spike (S), envelope (E), matrix (M), and nucleocapsid (N) proteins [36] (Figure 1). The complete genome sequence of SARS-CoV shares 53.1% identity with that of HCoV-OC43, suggesting its significant relation to HCoV-OC43 [31,37]. In particular, comparisons with the SARS-CoV nucleotide and amino acid sequences suggest that HCoV-OC43 shares highly homologous motifs necessary for viral replication and pathogenesis [31]. In this regard, HCoV-OC43 can be regarded as a good alternative model to study SARS-CoV biology without the need for level three containment facilities [31]. However, unlike SARS-CoV, HCoV-OC43 and HCoV-HKU1 contain extra NS2a and hemagglutinin-esterase (HE) genes in front of the S gene (Figure 2). The relatively high homology (92%) of the HCoV-OC43 NS2a protein with that of BCoV strongly suggests its bovine origin [38]. As shown in Figure 2, the 3′ terminal region of the HCoV-OC43 genome encodes the NS2a, HE, S, NS5a, E, M, and N genes in order [28,39,40,41]. The exact roles of the accessory proteins such as NS2a and 5a in the life cycle of HCoV-OC43 are unknown, although they are suggested to be associated with the host immune evasion process [42,43].
Thus far, only three papers have described the functions of the NS proteins of HCoV-OC43 in the context of HCoV-OC43 biology [38,44,45]. Mounir et al. characterized the nucleotide and amino acid sequences of NS2 [45]. Labonte et al. confirmed the expression of the NS2 protein in HCoV-OC43-infected HRT-18 cells. Dolliver et al. showed the inhibitory ability of the NS1 protein of HCoV-OC43 against stress granule formation [44]. Since most of early studies examined the functions of the structural proteins of HCoV-OC43, we focus on the description of characteristics of the structural proteins of HCoV-OC43 in the following section.
6. Characteristics of Structural Proteins of HCoV-OC43
6.1. Hemagglutinin-Esterase
Only group 2a coronaviruses have an envelope-associated HE gene [46]. Among the structural proteins expressed by HCoV-OC43, HE possesses several unique characteristics, which are absent in SARS-CoVs [12]. The HE gene of HCoV-OC43 is hypothesized to have borrowed from the influenza C virus by heterologous recombination through a common ancestral origin [47,48]. Morphologically, HE is observed to be interspersed as stubby homodimeric projections in the HCoV-OC43 virion [49,50,51]. In general, HE possesses dual receptor-binding and receptor-degrading functions (Table 1). A lectin domain specific for O-acetylated sialic acid and a sialate-O-acetylesterase domain are required to exert these two functions in HE [52,53,54,55,56]. In HCoV-OC43, the inhibition of the acetyl esterase activity of HE led to the blockage of the production of infectious viruses and subsequent virus spread in cell culture [57]. Whereas the carbohydrate-binding activity of HE is strictly maintained in BCoV [58], the progressive accumulation of mutations has rendered HCoV-OC43 incapable of this function. This seems to be a strategy of HCoV-OC43 for adaptation to a new host, the human [59]. Despite this loss, HCoV-OC43 was still able to execute a reversible virion–sialic acid attachment strategy similar to the influenza virus by using its S protein [60]. In this regard, the S and HE proteins of HCoV-OC43 appear to have coevolved into functionally interdependent viral structural proteins to maximize their host attachment and release abilities [60].
6.2. Spike Protein
The amino acid sequence of the S protein of HCoV-OC43 demonstrates 91% similarity with that of BCoV [63]. Only the S protein was able to agglutinate chicken erythrocytes; thus, the S protein can be regarded as the main determinant for hemagglutination by HCoV-OC43 (Table 1) [61]. In regards to carbohydrate-binding specificity, the S protein of HCoV-OC43 has shown a high binding affinity to carbohydrates with alpha 2,6 linkage [61]. For most coronaviruses, the C-terminal domain of the S protein functions as the protein receptor-binding domain (RBD), whereas the N-terminal domain of the S protein was shown to mediate sugar receptor-binding in HCoV-OC43 [64].
6.3. Envelope Protein
The homo-oligomerization of coronavirus structural E protein leads to the formation of ion channels (Table 1) [62]. Functionally, the HCoV-OC43 E protein is required not only for the production of infectious viruses but also for the development of neurovirulence in animals [62]. In addition, the HCoV-OC43 E protein and its transmembrane domain were shown to be essential for efficient replication in the murine central nervous system (CNS) [62]. Of note, the NS5a protein of HCoV-OC43 was also shown to be another virally encoded ion channel. It was demonstrated to be involved in virion morphogenesis and pathogenesis [65].
6.4. Membrane Protein
The amino acid sequence of the M protein of HCoV-OC43 shows 94% homology with that of BCoV. Its N-terminal domain possesses six potential O-glycosylation sites [45]. The nature of this O-glycosylation was further verified by the failure to inhibit the glycosylation of the M protein by treatment of tunicamycin [45]. In line with its structural functionality, the M protein has also been shown to be required for virion morphogenesis (Table 1) [45].
6.5. Nucleocapsid
The nucleotide and amino acid sequences of the N gene of HCoV-OC43 displayed high homology to those of BCoV [66]. The binding of the N protein to the viral RNA genome is necessary for the formation of a helical nucleocapsid that is coated by the viral envelope (Table 1) [1]. An internal N protein is thought to be in touch with the internal portion of the M protein [67]. The N-terminal of the HCoV-OC43 N protein has three RNA-binding regions (residues 1–173). The C-terminal region (residues 301–448) of the HCoV-OC43 N protein does not have RNA-binding activity. Most of this region is responsible for the oligomerization of the HCoV-OC43 N protein [68]. The N-terminal domain of the N protein from HCoV-OC43 is enriched with positively charged amino acids. This domain has been found to mediate RNA-binding to form the RNA genome¬–capsid complex [69].
7. Evolution of HCoV-OC43
The mutating abilities of coronaviruses and their high recombination frequencies have been the main driving forces for introducing a new coronavirus from animals to humans [70,71]. In particular, cattle and swine have served as intermediate hosts for interspecies transmission [28,72]. Therefore, its constant genetic recombination with different coronaviruses in animal populations is thought to have contributed to its evolution into the current form of HCoV-OC43 [36,73].
Understanding the adaptive process of an animal coronavirus to a human host is a prerequisite to revealing the origin of the COVID-19 pandemic [28]. Remarkable antigenic and genetic similarities have been consistently found between HCoV-OC43 and BCoV [28,74,75,76,77]. In essence, HCoV-OC43 appears to have emerged from a spillover of BCoV [60]. The sequence analysis of the S gene of BCoV and HCoV-OC43 indicates a high probability of a recent host jump from animals to humans [28]. Specifically, HCoV-OC43 was postulated to have diverged from BCoV in 1890 [28]. The absence of 290 nucleotides from the S gene of HCoV-OC43 compared with that of BCoV seems to have been necessary for HCoV-OC43 to adapt to a new host. However, despite their high levels of similarity, HCoV-OC43 has maintained several unique features distinct from BCoV. First, two coding regions located downstream of the S gene of BCoV are missing in HCoV-OC43 [78]. In addition, HCoV-OC43 has also abandoned the HE lectin function as an adaptive effort to become a human pathogen [60]. This carbohydrate-binding activity seems to be provided by the S protein, instead. In addition to interspecies evolution, the co-circulation of numerous recombinant variants seems to give rise to the intra-species variability in HCoV-OC43 [79]. In particular, the genetic instability of the S gene seems to be pivotal for genotype persistence in human populations [64]. The generation of a novel genotype by natural recombination appears likely to continue for the foreseeable future [41]. In addition, regions of the viral S protein of HCoV-OC43 are continually exposed to human humoral immunity. Since HCoV-OC43 will undergo adaptive evolution in these regions, these adaptive alterations in antigenic regions of the virus will require the continual reformulation of already-made vaccines [80].
8. Host Interactions with HCoV-OC43
8.1. Virus Entry Factors
Sialic acids are found as terminal sugars of oligosaccharides present on many glycoproteins and glycolipids on cell surfaces [81]. Like influenza viruses, HCoV-OC43 utilizes a sialic acid as an entry receptor (Table 2) [6,82,83,84]. Since this receptor binding is mediated by the S protein, it is regarded as the main determinant for coronavirus host specificity [6]. Although two studies have reported the major histocompatibility complex (MHC) class I as another potential host receptor for HCoV-OC43 (Table 2) [18,85], there were no follow-up studies to validate this discovery. As explained previously, HCoV-OC43 and BCoV exhibit remarkable antigenic and genetic similarities. In addition, they both can agglutinate erythrocytes by attachment to N-acetyl-9-O-acetylneuraminic acid [81]. The interaction between HCoV-OC43 and N-acetyl-9-O-acetylneuraminic acid is required not only for erythrocytes agglutination but also for target cell infection with a preference for the alpha 2,6 linkage [61,81]. After attachment, HCoV-OC43 has been shown to employ caveolin-1-dependent endocytosis for viral entry and dynamin-dependent budding for viral exit (Table 2) [86]. In particular, the internalization of a virus particle was shown to require actin cytoskeleton rearrangements [86].
As previously mentioned, the most striking difference between HCoV-OC43 and SARS-CoV-2 might be the usage of host receptors for their entry. HCoV-OC43 takes advantage of 9-O-acetyl sialic acid for its initial attachment to the host cell [6,82,83,84] and engages with MHC class I to reach the host cytoplasm [18,85], while SARS-CoVs mainly depends on angiotensin converting enzyme 2 (ACE2) for this same viral process [91]. Paradoxically, infection with HCoV-OC43 can even induce the expression of MHC class I to boost its entry [92]. Based on these observations, HCoV-OC43 does not seem to be the ideal alternative to study the entry process of SARS-CoVs. Therefore, extra caution needs to be taken to interpret the virus entry-related data obtained from the HCoV-OC43 system for their potential application to the development of vaccines and antivirals against SARS-CoVs.
8.2. Intracellular Host Factors
After viral entry, coronaviruses take advantage of various host factors to support their viral life cycle inside infected cells. To identify these necessary host factors, several protein¬–protein interaction studies were conducted. Davies et al. reported strong interaction of viral NS2 and NS4 from HCoV-OC43 and SARS-CoVs with proteins localized at mitochondria-associated ER membranes (Table 2) [87]. This study further confirms the importance of the ER as the major subcellular organelle to support the genome replication of coronaviruses. To validate a recently published SARS-CoV-2 protein interactome, Hoffman et al. designed a CRISPR¬–Cas9 library targeting 332 proteins [88]. According to this study, they found virus-specific differences in Rab GTPase requirements and glycosylphosphatidylinositol anchor biosynthesis. They also identified multiple pan-coronavirus factors involved in cholesterol homeostasis. They seem to be necessary for virus assembly and trafficking (Table 2) [88]. Another group of researchers also conducted genome-wide CRISPR screening by using HCoV-OC43, HCoV-229, and SARS-CoV-2 [89]. In this study, they confirmed the already-known viral entry factors ACE2 (SARS-CoV-2), aminopeptidase N (HCoV-229E), and glycosaminoglycans (HCoV-OC43). In addition, this study also identified endosome maturation, phosphatidylinositol phosphate biosynthesis, and cholesterol homeostasis as essential host pathways for all three coronaviruses. These host processes seem to be linked to virus assembly and trafficking processes [89]. In line with this, pharmacological inhibition of phosphatidylinositol kinases and cholesterol homeostasis led to the attenuated propagation of these coronaviruses (Table 2) [89]. Schneider et al. also conducted similar genome-scale CRISPR knockout screening by using HCoV-OC43, HCoV-NL63, HCoV-229, and SARS-CoV-2 [90]. According to this study, they identified the vacuole membrane protein 1 (VMP1), transmembrane protein 41B (TMEM41B), and TMEM64 as host factors required for infection by all these coronaviruses. These host factors were proposed to be involved with ER membrane remodeling (Table 2) [90]. All of these screening results further validate the utility of HCoV-OC43 as an alternative research tool to study high-risk SARS-CoVs.
8.3. Host Immunological Responses
Innate and adaptive host immune responses to virus infection are critical components of the host immune system to maintain a virus-free host environment. Therefore, as a countermeasure to the host defense, a viral immune evasion strategy is also essential for virus survival inside the host cell. The secretion of interferon (IFN) by infected host cells after virus recognition plays a central role in establishing innate immune defense in neighboring cells against virus infection. In general, interferons are detrimental to a virus due to the activation of MHC antigen processing and the antiviral action of various interferon-stimulated genes (ISGs). Paradoxically, IFN-γ was shown to enhance HCoV-OC43 infection of neuronal cells by increasing the expression of MHC class I, which was proposed as another viral entry receptor [92]. This increased host cell susceptibility to HCoV-OC43 infection was further validated by a receptor blockade with a monoclonal antibody specific for MHC class I [92]. In this regard, HCoV-OC43 seems to be able to take advantage of the virus-induced antiviral actions of IFNs. On the other hand, the expression of the HCoV-OC43 N protein led to the potentiation of NF-kB activation [93]. Mechanistically, the binding of the N protein to microRNA 9 (miR-9) was shown to relieve the negative regulation of NF-kB by miR-9 [93]. In support of the positive role of IFNs in the coronavirus life cycle, all three types of IFNs including IFN-α, IFN-γ, and IFN-λ were shown to efficiently promote HCoV-OC43 infection [94]. This enhancement of HCoV-OC43 infection by IFNs seems to be related to the increased availability of human IFN-induced transmembrane protein 2 (IFITM2) and IFITM3, which were also shown to serve as additional entry factors for HCoV-OC43 [94]. In regards to the blockage of induction of innate immunity by HCoV-OC43 infection, the transcriptional activities of interferon-sensitive response elements (ISREs), as well as IFN-β and NF-κB promoters, were significantly inhibited by HCoV-OC43 accessory proteins such as NS2a and NS5a and structural proteins such as M and N [42,43]. In line with this negative modulation of innate immunity by HCoV-OC43, Loo et al. also reported negligible innate immune activation by HCoV-OC43 infection in differentiated primary human bronchial epithelial cells [95].
With regard to the adaptive immunity induced by HCoV-OC43, SARS-CoV-2 antibodies cross-reacted with the S proteins of other beta coronaviruses [96]. From the clinical perspective, COVID-19 patients showed more severe disease progression when they had significantly lower levels of antibodies against the HCoV-OC43 N protein [97]. These results indicate the protective effects of previous infections with seasonal low-risk coronaviruses against COVID-19 [97]. Low pathogenic HCoVs strains such as OC43 and NL63 were also shown to induce T cells cross-reactive to SARS-CoV-2 [98]. In addition, memory CD8+ T cells specific for endemic coronavirus were also suggested to affect immune responses against SARS-CoV-2 [99]. This finding was remarkable considering a lower percentage (5.4%) of the epitope sharing of four endemic coronaviruses including 229E, HKU1, NL63, and OC43 with SARS-CoV-2. When the serum of SARS-CoV patients was examined, N peptide sequences were shown to play a critical role in cross-reactivity between HCoV-OC43 and SARS-CoV [100]. In general, a more severe clinical manifestation was observed following SARS-CoV-2 infection in older COVID-19 patients. This observation could be ascribed to the absence of pre-experienced T-cell immunity induced by endemic coronaviruses in elderly subjects [98]. Specifically, the C-terminal epitopes in the S protein of SARS-CoV-2 display a higher homology to the S proteins of low-risk coronaviruses such as 229E and OC43 [101]. Previous encounters with endemic coronaviruses might give rise to cross-reactive T cells specific for S proteins. Consequently, they were thought to play a protective role against COVID-19 pathogenesis [101]. In support of this finding, Mateus et al. also confirmed the presence of preexisting memory CD4+ T cells that are cross-reactive to SARS-CoV-2 and HCoV-OC43 [102]. Grifoni et al. also detected SARS-CoV-2-specific CD4+ T cells in unexposed cohorts. These data indicate the overlapping nature of T-cell recognition between low-risk coronaviruses and SARS-CoV-2 [103].
9. Neuropathology by HCoV-OC43 Infection
9.1. In Vitro Model
Human coronaviruses can infect some CNS-derived cells including astrocytoma, neuroblastoma, and the immortalized fetal microglial cell lines [104,105,106]. However, the infection of these neuronal cells failed to produce infectious HCoV-OC43 virions [107,108]. In general, the infection of glial cells by coronaviruses can lead to inflammation and induction of apoptosis, which are two relevant events for CNS pathologies [109]. Nevertheless, a causative connection between HCoV-OC43 and neurologic diseases remains unproven [110,111]. Continuous propagation of HCoV-OC43 in human neural cell cultures resulted in mutations in the viral S glycoprotein [112]. In particular, highly enhanced cell death in murine and human neuronal cells was observed by infection of HCoV-OC43 with two-point mutations in the S glycoprotein [113]. These data suggest the importance of S protein as a key determinant for HCoV-OC43-induced neuropathology.
9.2. In Vivo Model
Intracerebral or extraneural infection of suckling CD1 mice with HCoV-OC43 resulted in a neurotropic lethality in infected mice. High titers of the HCoV-OC43 virus were recovered from their brains. However, when mice grew older than 20 days, they became completely insusceptible to HCoV-OC43 inoculation. Mechanistically, the immune response seems to be ascribable to the development of resistance to HCoV-OC43 infection, in part [114]. BALB/c mice were also shown to be susceptible to an acute and persistent HCoV-OC43 infection [115]. Acute encephalitis was observed by intracerebral injection of HCoV-OC43 into BALB/c mice, followed by neuronal cell death by necrosis and apoptosis [110]. Axonal transport appears to facilitate the neuron-to-neuron propagation of HCoV-OC43 in mice brains [116]. There seems to be a significant difference in the degree of neurovirulence between mouse-CNS-adapted versus tissue-culture-adapted HCoV-OC43, since the intranasal inoculation of 8-week mice with a mouse-CNS-adapted strain was uniformly fatal while the tissue-culture-adapted strain was not [117]. The surface glycoprotein is thought to play a major role in the development of virulence in coronavirus infections. As mentioned previously, the acquirement of mutations in the S glycoprotein during viral persistence was able to produce more severe neurovirulence phenotypes when inoculated into susceptible mice. For example, the mutant HCoV-OC43 with S glycoprotein point mutations (H183R and Y241H) was able to give rise to a heightened unfolded protein response and attenuated protein translation. Therefore, this mutant virus turned out to be more neurovirulent in mice [118]. In addition, this mutant HCoV-OC43 was able to induce a highly deteriorated neuropathology in BALB/c mice. This mutant HCoV-OC43 was also involved with increased viral spread and enhanced T-cell infiltration, as well as the enhanced production of proinflammatory cytokines [112]. Mechanistically, receptor-interacting protein kinase 1 (RIP1) and mixed lineage kinase domain-like (MLKL) genes seem to play a pivotal role in the neuronal cell death induced by HCoV-OC43 [113]. A reporter HCoV-OC43 strain expressing renilla luciferase induced fatality in suckling mice after intranasal inoculation [119]. Le Coupanec et al. suggested the strong association of efficient cleavage of the S protein with HCoV-OC43-induced neurovirulence [120].
9.3. Clinical Model
HCoV-OC43 has been detected in the brain of pediatric patients with acute disseminated encephalomyelitis or fatal encephalitis [121,122]. This HCoV-OC43-induced fatal encephalitis was also diagnosed in an infant with aplastic thymus and chronic T-cell lymphopenia [123]. However, an etiologic connection between HCoVs and neurotropic diseases is not yet clear [111]. Although a higher prevalence of HCoV-OC43 in multiple sclerosis patients than in controls was observed [124], no significant differences were found between the multiple sclerosis patients and the normal subjects in their antibody titer to HCoV-229E and HCoV-OC43 [125].
10. Discovery of Antiviral Candidates by Using HCoV-OC43
Several antiviral candidates for coronavirus infection were identified by using HCo-OC43 to treat endemic coronavirus infections. Since the COVID-19 pandemic, the utility of HCoV-OC43 has been further applied to the discovery of anti-SARS-CoV-2 compounds due to its low-risk potential. Degiaggi et al. found that phosphatidyl-serine was able to inhibit HCoV-OC43 (Table 3) [126]. Cystatin C and D, potent inhibitors of cysteine proteases such as papain and cathepsin B, were also shown to inhibit HCoV-OC43 at its physiologic concentration (Table 3) [127,128]. Chloroquine, which once showed promise as a new class of SARS-CoV-2 drugs, potently suppressed HCoV-OC43 replication in vitro, with an IC50 of 0.33 μM by using the HCoV-OC43 reporter virus with a renilla luciferase gene (Table 3) [129]. Chloroquine was also able to reduce the mortality of HCoV-OC43-infected newborn C57BL/6 mice after transplacental or maternal milk delivery [130]. Emodin, an active component of several plants used in traditional Chinese medicine was able to inhibit the 3a ion channel of coronavirus SARS-CoV and HCoV-OC43 (Table 3) [131]. Meanwhile, memantine, an NMDA receptor antagonist was demonstrated to attenuate mortality rates and body weight loss in the HCoV-OC43-infected mice (Table 3) [132]. HTCC, a cationically modified chitosan, inhibited the replication of several endemic strains of human coronaviruses, including HCoV-NL63, HCoV-OC43, and HCoV-HKU1 replication in human airway epithelial cells, by disrupting the virus–receptor interactions (Table 3) [133]. The mechanism of the antiviral actions of this polymer seems to involve the blockage of virus entry into the host cell by interaction with the S protein [134]. Lycorine, a toxic crystalline alkaloid found in various Amaryllidaceae, was able to reduce the fatality of BALB/c mice by HCoV-OC43 by attenuating the viral spread in the CNS of infected mice (Table 3) [135]. Kim et al. reported that bis-benzylisoquinoline alkaloids were able to significantly inhibit HCoV-OC43-induced cell death at the early step of the virus life cycle (Table 3) [136]. Amiloride, which is a diuretic to treat hypertension, was shown to inhibit HCoV-OC43 replication via specific interactions with stem-loop structures of viral RNAs (Table 3) [137]. Tylophorine-based compounds and natural cardiotonic steroids such as cardenolides and bufadienolides were also able to inhibit HCoV-OC43 with nanomolar EC50 values (Table 3) [138]. Kurarinone, a flavanone from Sophora flavescens roots, inhibited HCoV-OC43 infection in human lung fibroblasts with an IC50 of 3.458 μM by impairing the virus-induced autophagy (Table 3) [139]. An anti-protozoal drug, emetine, has also been reported to have anti-coronavirus activity (Table 3) [140]. This compound was again found as an HCoV-OC43 inhibitor in HCT-8 cells (Table 3) [141]. The in vitro inhibitory capacity of the redox-active oxysterol 27-hydroxycholesterol against SARS-CoV-2 was further verified by using HCoV-OC43 (Table 3) [142]. Valinomycin, a naturally occurring dodecadepsipeptide, has been used as an antibiotic; its broad-spectrum antiviral activity was further confirmed by using HCoV-OC43 (Table 3) [40]. AT-527, a prodrug of a guanosine nucleotide analog was identified as a potent inhibitor of both SARS-CoV-2 and HCoV-OC43 in vitro (Table 3) [143]. Epigallocatechin gallate (EGCG), which is a green tea polyphenol, was able to decrease the 3CL-protease activity of HCoV-OC43 and HCoV-229E. In addition, EGCG treatment attenuated HCoV-OC43-associated cytotoxicity (Table 3) [144]. Lactoferrin is a multifunctional protein of the transferrin family. The broad-spectrum antiviral activity of lactoferrin was also confirmed by using HCoV-OC43. The binding of lactoferrin to heparan sulfate proteoglycans seems to be responsible for blocking viral attachment to the host cell (Table 3) [145].
11. Conclusions
In this review paper, we overviewed the history, classification, and clinical manifestations of HCoV-OC43. The characteristics of its genomes and genes and its evolution process were further explored. The essential host factors for the viral life cycle and the innate, as well as adaptive, immunological responses were also summarized. Finally, the available in vitro and in vivo systems to study HCoV-OC43 and its application to the discovery of potential antivirals for COVID-19 were also presented. HCoV-OC43 should provide a valuable tool, not only for the general study of coronavirus biology but also for the rapid identification of promising drugs against coronaviruses without the drawbacks of biosafety level 3 confinement.
Author Contributions
Writing—original draft preparation, M.I.K.; writing—review and editing, C.L.; visualization, C.L.; supervision, C.L.; funding acquisition, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MEST) (NRF-2022R1A2C1011629). This work was also supported by the Dongguk University Research Fund of 2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Masters, P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar] [CrossRef]
- Cimolai, N. Complicating Infections Associated with Common Endemic Human Respiratory Coronaviruses. Health Secur. 2020, 19, 195–208. [Google Scholar] [CrossRef]
- Gaunt, E.R.; Hardie, A.; Claas, E.C.; Simmonds, P.; Templeton, K.E. Epidemiology and clinical presentations of the four human coronaviruses 229E, HKU1, NL63, and OC43 detected over 3 years using a novel multiplex real-time PCR method. J. Clin. Microbiol. 2010, 48, 2940–2947. [Google Scholar] [CrossRef] [Green Version]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, N.M. The epidemiology of acute respiratory infections in children and adults: A global perspective. Epidemiol. Rev. 1990, 12, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; van Kuppeveld, F.J.M.; Boons, G.J.; Bosch, B.J.; et al. Human coronaviruses OC43 and HKU1 bind to 9-O-acetylated sialic acids via a conserved receptor-binding site in spike protein domain A. Proc. Natl. Acad. Sci. USA 2019, 116, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Chen, X.; Hu, T.; Li, J.; Song, H.; Liu, Y.; Wang, P.; Liu, D.; Yang, J.; Holmes, E.C.; et al. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr. Biol. 2020, 30, 2196–2203.e2193. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, T.; Smith, S.L.; Radford, A.D.; Solomon, T.; Hughes, G.L.; Patterson, E.I. SARS-CoV-2 Infections in Animals: Reservoirs for Reverse Zoonosis and Models for Study. Viruses 2021, 13, 494. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Yeh, I.J.; Phan, N.N.; Wu, Y.H.; Yen, M.C.; Hung, J.H.; Chiao, C.C.; Chen, C.F.; Sun, Z.; Jiang, J.Z.; et al. Gene signatuRes. of SARS-CoV/SARS-CoV-2-infected ferret lungs in short- and long-term models. Infect. Genet. Evol. 2020, 85, 104438. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Dees, J.H.; Becker, W.B.; Kapikian, A.Z.; Chanock, R.M. Recovery in tracheal organ cultuRes. of novel viruses from patients with respiratory disease. Proc. Natl. Acad. Sci. USA 1967, 57, 933–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, J.S.; McIntosh, K. History and recent advances in coronavirus discovery. Pediatr. Infect. Dis. J. 2005, 24, S223–S227, discussion S226. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Becker, W.B.; Chanock, R.M. Growth in suckling-mouse brain of “IBV-like” viruses from patients with upper respiratory tract disease. Proc. Natl. Acad. Sci. USA 1967, 58, 2268–2273. [Google Scholar] [CrossRef] [Green Version]
- Bracci, N.; Pan, H.C.; Lehman, C.; Kehn-Hall, K.; Lin, S.C. Improved plaque assay for human coronaviruses 229E and OC43. PeerJ. 2020, 8, e10639. [Google Scholar] [CrossRef]
- Kaye, H.S.; Dowdle, W.R. Some characteristics of hemagglutination of certain strains of “IBV-like” virus. J. Infect. Dis. 1969, 120, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Myint, S.; Johnston, S.; Sanderson, G.; Simpson, H. Evaluation of nested polymerase chain methods for the detection of human coronaviruses 229E and OC43. Mol. Cell. Probes 1994, 8, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Wege, H.; Siddell, S.; ter Meulen, V. The biology and pathogenesis of coronaviruses. Curr. Top. Microbiol. Immunol. 1982, 99, 165–200. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R. Human coronavirus OC43 interacts with major histocompatibility complex class I molecules at the cell surface to establish infection. Immunol. Investig. 1994, 23, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Ogimi, C.; Kim, Y.J.; Martin, E.T.; Huh, H.J.; Chiu, C.H.; Englund, J.A. What’s New With the Old Coronaviruses? J. Pediatr. Infect. Dis. Soc. 2020, 9, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Jean, A.; Quach, C.; Yung, A.; Semret, M. Severity and outcome associated with human coronavirus OC43 infections among children. Pediatr. Infect. Dis. J. 2013, 32, 325–329. [Google Scholar] [CrossRef]
- Kim, J.M.; Jeon, J.S.; Kim, J.K. Climate and Human coronaviruses 229E and Human coronaviruses OC43 Infections: Respiratory Viral Infections Prevalence in Hospitalized Children in Cheonan, Korea. J. Microbiol. Biotechnol. 2020, 30, 1495–1499. [Google Scholar] [CrossRef]
- Talbot, H.K.; Shepherd, B.E.; Crowe, J.E., Jr.; Griffin, M.R.; Edwards, K.M.; Podsiad, A.B.; Tollefson, S.J.; Wright, P.F.; Williams, J.V. The pediatric burden of human coronaviruses evaluated for twenty years. Pediatr. Infect. Dis. J. 2009, 28, 682–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, S.K.; Lee, P.; Tsang, A.K.; Yip, C.C.; Tse, H.; Lee, R.A.; So, L.Y.; Lau, Y.L.; Chan, K.H.; Woo, P.C.; et al. Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombination. J. Virol. 2011, 85, 11325–11337. [Google Scholar] [CrossRef] [Green Version]
- Monto, A.S.; Lim, S.K. The Tecumseh study of respiratory illness. VI. Frequency of and relationship between outbreaks of coronavirus infection. J. Infect. Dis. 1974, 129, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Morgello, S. Coronaviruses and the central nervous system. J. Neurovirol. 2020, 26, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Nickbakhsh, S.; Ho, A.; Marques, D.F.P.; McMenamin, J.; Gunson, R.N.; Murcia, P.R. Epidemiology of Seasonal Coronaviruses: Establishing the Context for the Emergence of Coronavirus Disease 2019. J. Infect. Dis. 2020, 222, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saib, I.; Aleisa, S.; Ardah, H.; Mahmoud, E.; Alharbi, A.O.; Alsaedy, A.; Aljohani, S.; Alshehri, A.; Alharbi, N.K.; Bosaeed, M. Non-SARS Non-MERS Human Coronaviruses: Clinical Characteristics and Outcome. Pathogens 2021, 10, 1549. [Google Scholar] [CrossRef]
- Vijgen, L.; Keyaerts, E.; Moes, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.M.; Van Ranst, M. Complete genomic sequence of human coronavirus OC43: Molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- El-Sahly, H.M.; Atmar, R.L.; Glezen, W.P.; Greenberg, S.B. Spectrum of clinical illness in hospitalized patients with “common cold” virus infections. Clin. Infect. Dis. 2000, 31, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Gagneur, A.; Sizun, J.; Vallet, S.; Legr, M.C.; Picard, B.; Talbot, P.J. Coronavirus-related nosocomial viral respiratory infections in a neonatal and paediatric intensive care unit: A prospective study. J. Hosp. Infect. 2002, 51, 59–64. [Google Scholar] [CrossRef]
- St-Jean, J.R.; Jacomy, H.; Desforges, M.; Vabret, A.; Freymuth, F.; Talbot, P.J. Human respiratory coronavirus OC43: Genetic stability and neuroinvasion. J. Virol. 2004, 78, 8824–8834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.N.; Mounir, S.; Talbot, P.J. Human coronavirus gene expression in the brains of multiple sclerosis patients. Virology 1992, 191, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.S.; Brown, B.; Brian, D.; Cabirac, G.F. Detection of coronavirus RNA and antigen in multiple sclerosis brain. Ann. Neurol. 1992, 31, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.; Dixit, D. Human and novel coronavirus infections in children: A review. Paediatr. Int. Child. Health 2020, 41, 36–55. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Quan, Y.; Xin, Z.T.; Wrammert, J.; Ma, M.J.; Lv, H.; Wang, T.B.; Yang, H.; Richardus, J.H.; Liu, W.; et al. Lack of peripheral memory B cell responses in recovered patients with severe acute respiratory syndrome: A six-year follow-up study. J. Immunol. 2011, 186, 7264–7268. [Google Scholar] [CrossRef] [Green Version]
- Leao, J.C.; Gusmao, T.P.L.; Zarzar, A.M.; Leao Filho, J.C.; Barkokebas Santos de Faria, A.; Morais Silva, I.H.; Gueiros, L.A.M.; Robinson, N.A.; Porter, S.; Carvalho, A.A.T. Coronaviridae-Old friends, new enemy! Oral Dis. 2020, 28, 858–866. [Google Scholar] [CrossRef]
- Marra, M.A.; Jones, S.J.; Astell, C.R.; Holt, R.A.; Brooks-Wilson, A.; Butterfield, Y.S.; Khattra, J.; Asano, J.K.; Barber, S.A.; Chan, S.Y.; et al. The Genome sequence of the SARS-associated coronavirus. Science 2003, 300, 1399–1404. [Google Scholar] [CrossRef] [Green Version]
- Labonte, P.; Mounir, S.; Talbot, P.J. Sequence and expression of the ns2 protein gene of human coronavirus OC43. J. Gen. Virol. 1995, 76 Pt 2, 431–435. [Google Scholar] [CrossRef]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Ma, Z.; Chen, H.; Lu, Y.; Chen, X. Valinomycin as a potential antiviral agent against coronaviruses: A review. Biomed. J. 2020, 43, 414–423. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Xiao, Y.; Zhang, J.; Wang, Y.; Chen, L.; Paranhos-Baccala, G.; Ren, L.; Wang, J. Genotype shift in human coronavirus OC43 and emergence of a novel genotype by natural recombination. J. Infect. 2015, 70, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beidas, M.; Chehadeh, W. PCR array profiling of antiviral genes in human embryonic kidney cells expressing human coronavirus OC43 structural and accessory proteins. Arch. Virol. 2018, 163, 2065–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beidas, M.; Chehadeh, W. Effect of Human Coronavirus OC43 Structural and Accessory Proteins on the Transcriptional Activation of Antiviral Response Elements. Intervirology 2018, 61, 30–35. [Google Scholar] [CrossRef]
- Dolliver, S.M.; Kleer, M.; Bui-Marinos, M.P.; Ying, S.; Corcoran, J.A.; Khaperskyy, D.A. Nsp1 proteins of human coronaviruses HCoV-OC43 and SARS-CoV2 inhibit stress granule formation. PLoS Pathog. 2022, 18, e1011041. [Google Scholar] [CrossRef] [PubMed]
- Mounir, S.; Labonte, P.; Talbot, P.J. Characterization of the nonstructural and spike proteins of the human respiratory coronavirus OC43: Comparison with bovine enteric coronavirus. Adv. Exp. Med. Biol. 1993, 342, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, M.M.; Cavanagh, D. The molecular biology of coronaviruses. Adv. Virus Res. 1997, 48, 1–100. [Google Scholar]
- Zhang, X.M.; Kousoulas, K.G.; Storz, J. The hemagglutinin/esterase gene of human coronavirus strain OC43: Phylogenetic relationships to bovine and murine coronaviruses and influenza C virus. Virology 1992, 186, 318–323. [Google Scholar] [CrossRef]
- Luytjes, W.; Bredenbeek, P.J.; Noten, A.F.; Horzinek, M.C.; Spaan, W.J. Sequence of mouse hepatitis virus A59 mRNA 2: Indications for RNA recombination between coronaviruses and influenza C virus. Virology 1988, 166, 415–422. [Google Scholar] [CrossRef]
- Lissenberg, A.; Vrolijk, M.M.; van Vliet, A.L.; Langereis, M.A.; de Groot-Mijnes, J.D.; Rottier, P.J.; de Groot, R.J. Luxury at a cost? Recombinant mouse hepatitis viruses expressing the accessory hemagglutinin esterase protein display reduced fitness in vitro. J. Virol. 2005, 79, 15054–15063. [Google Scholar] [CrossRef] [Green Version]
- Kienzle, T.E.; Abraham, S.; Hogue, B.G.; Brian, D.A. Structure and orientation of expressed bovine coronavirus hemagglutinin-esterase protein. J. Virol. 1990, 64, 1834–1838. [Google Scholar] [CrossRef] [Green Version]
- King, B.; Brian, D.A. Bovine coronavirus structural proteins. J. Virol. 1982, 42, 700–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, B.; Potts, B.J.; Brian, D.A. Bovine coronavirus hemagglutinin protein. Virus Res. 1985, 2, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Vlasak, R.; Luytjes, W.; Leider, J.; Spaan, W.; Palese, P. The E3 protein of bovine coronavirus is a receptor-destroying enzyme with acetylesterase activity. J. Virol. 1988, 62, 4686–4690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultze, B.; Wahn, K.; Klenk, H.D.; Herrler, G. Isolated HE-protein from hemagglutinating encephalomyelitis virus and bovine coronavirus has receptor-destroying and receptor-binding activity. Virology 1991, 180, 221–228. [Google Scholar] [CrossRef]
- Zeng, Q.; Langereis, M.A.; van Vliet, A.L.; Huizinga, E.G.; de Groot, R.J. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 9065–9069. [Google Scholar] [CrossRef] [Green Version]
- de Groot, R.J. Structure, function and evolution of the hemagglutinin-esterase proteins of corona- and toroviruses. Glycoconj. J. 2006, 23, 59–72. [Google Scholar] [CrossRef]
- Desforges, M.; Desjardins, J.; Zhang, C.; Talbot, P.J. The acetyl-esterase activity of the hemagglutinin-esterase protein of human coronavirus OC43 strongly enhances the production of infectious virus. J. Virol. 2013, 87, 3097–3107. [Google Scholar] [CrossRef] [Green Version]
- Langereis, M.A.; Bakkers, M.J.; Deng, L.; Padler-Karavani, V.; Vervoort, S.J.; Hulswit, R.J.; van Vliet, A.L.; Gerwig, G.J.; de Poot, S.A.; Boot, W.; et al. Complexity and Diversity of the Mammalian Sialome Revealed by Nidovirus Virolectins. Cell Rep. 2015, 11, 1966–1978. [Google Scholar] [CrossRef] [Green Version]
- Bakkers, M.J.; Lang, Y.; Feitsma, L.J.; Hulswit, R.J.; de Poot, S.A.; van Vliet, A.L.; Margine, I.; de Groot-Mijnes, J.D.; van Kuppeveld, F.J.; Langereis, M.A.; et al. Betacoronavirus Adaptation to Humans Involved Progressive Loss of Hemagglutinin-Esterase Lectin Activity. Cell Host Microbe 2017, 21, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Lang, Y.; Li, W.; Li, Z.; Koerhuis, D.; van den Burg, A.C.S.; Rozemuller, E.; Bosch, B.J.; van Kuppeveld, F.J.M.; Boons, G.J.; Huizinga, E.G.; et al. Coronavirus hemagglutinin-esterase and spike proteins coevolve for functional balance and optimal virion avidity. Proc. Natl. Acad. Sci. USA 2020, 117, 25759–25770. [Google Scholar] [CrossRef]
- Kunkel, F.; Herrler, G. Structural and functional analysis of the surface protein of human coronavirus OC43. Virology 1993, 195, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Stodola, J.K.; Dubois, G.; Le Coupanec, A.; Desforges, M.; Talbot, P.J. The OC43 human coronavirus envelope protein is critical for infectious virus production and propagation in neuronal cells and is a determinant of neurovirulence and CNS pathology. Virology 2018, 515, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Mounir, S.; Talbot, P.J. Molecular characterization of the S protein gene of human coronavirus OC43. J. Gen. Virol. 1993, 74 Pt 9, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhang, Y.; Li, J.; Xiao, Y.; Zhang, J.; Wang, Y.; Chen, L.; Paranhos-Baccala, G.; Wang, J. Genetic drift of human coronavirus OC43 spike gene during adaptive evolution. Sci. Rep. 2015, 5, 11451. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, K.; Ping, X.; Yu, W.; Qian, Z.; Xiong, S.; Sun, B. The ns12.9 Accessory Protein of Human Coronavirus OC43 Is a Viroporin Involved in Virion Morphogenesis and Pathogenesis. J. Virol. 2015, 89, 11383–11395. [Google Scholar] [CrossRef] [Green Version]
- Kamahora, T.; Soe, L.H.; Lai, M.M. Sequence analysis of nucleocapsid gene and leader RNA of human coronavirus OC43. Virus Res. 1989, 12, 1–9. [Google Scholar] [CrossRef]
- Sturman, L.S.; Holmes, K.V.; Behnke, J. Isolation of coronavirus envelope glycoproteins and interaction with the viral nucleocapsid. J. Virol. 1980, 33, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Hsu, Y.L.; Chiang, W.L.; Hou, M.H. Elucidation of the stability and functional regions of the human coronavirus OC43 nucleocapsid protein. Protein Sci. 2009, 18, 2209–2218. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.J.; Chou, C.C.; Liu, C.L.; Lee, C.C.; Kan, L.S.; Hou, M.H. Crystallization and preliminary X-ray diffraction analysis of the N-terminal domain of human coronavirus OC43 nucleocapsid protein. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 815–818. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, C.M.; Gebauer, F.; Sune, C.; Mendez, A.; Dopazo, J.; Enjuanes, L. Genetic evolution and tropism of transmissible gastroenteritis coronaviruses. Virology 1992, 190, 92–105. [Google Scholar] [CrossRef]
- Lam, T.T.; Jia, N.; Zhang, Y.W.; Shum, M.H.; Jiang, J.F.; Zhu, H.C.; Tong, Y.G.; Shi, Y.X.; Ni, X.B.; Liao, Y.S.; et al. Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 2020, 583, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijgen, L.; Keyaerts, E.; Lemey, P.; Maes, P.; Van Reeth, K.; Nauwynck, H.; Pensaert, M.; Van Ranst, M. Evolutionary history of the closely related group 2 coronaviruses: Porcine hemagglutinating encephalomyelitis virus, bovine coronavirus, and human coronavirus OC43. J. Virol. 2006, 80, 7270–7274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, S.K.; Li, K.S.; Huang, Y.; Shek, C.T.; Tse, H.; Wang, M.; Choi, G.K.; Xu, H.; Lam, C.S.; Guo, R.; et al. Ecoepidemiology and complete genome comparison of different strains of severe acute respiratory syndrome-related Rhinolophus bat coronavirus in China reveal bats as a reservoir for acute, self-limiting infection that allows recombination events. J. Virol. 2010, 84, 2808–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogue, B.G.; King, B.; Brian, D.A. Antigenic relationships among proteins of bovine coronavirus, human respiratory coronavirus OC43, and mouse hepatitis coronavirus A59. J. Virol. 1984, 51, 384–388. [Google Scholar] [CrossRef] [Green Version]
- Lapps, W.; Brian, D.A. Oligonucleotide fingerprints of antigenically related bovine coronavirus and human coronavirus OC43. Arch. Virol. 1985, 86, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurier, F.; Beury, D.; Flechon, L.; Varre, J.S.; Touzet, H.; Goffard, A.; Hot, D.; Caboche, S. A complete protocol for whole-genome sequencing of virus from clinical samples: Application to coronavirus OC43. Virology 2019, 531, 141–148. [Google Scholar] [CrossRef]
- Vieler, E.; Schlapp, T.; Anders, C.; Herbst, W. Genomic relationship of porcine hemagglutinating encephalomyelitis virus to bovine coronavirus and human coronavirus OC43 as studied by the use of bovine coronavirus S gene-specific probes. Arch. Virol. 1995, 140, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Mounir, S.; Talbot, P.J. Human coronavirus OC43 RNA 4 lacks two open reading frames located downstream of the S gene of bovine coronavirus. Virology 1993, 192, 355–360. [Google Scholar] [CrossRef]
- Kin, N.; Miszczak, F.; Lin, W.; Gouilh, M.A.; Vabret, A.; Consortium, E. Genomic Analysis of 15 Human Coronaviruses OC43 (HCoV-OC43s) Circulating in France from 2001 to 2013 Reveals a High Intra-Specific Diversity with New Recombinant Genotypes. Viruses 2015, 7, 2358–2377. [Google Scholar] [CrossRef] [Green Version]
- Kistler, K.E.; Bedford, T. Evidence for adaptive evolution in the receptor-binding domain of seasonal coronaviruses OC43 and 229E. Elife 2021, 10, e64509. [Google Scholar] [CrossRef]
- Krempl, C.; Schultze, B.; Herrler, G. Analysis of cellular receptors for human coronavirus OC43. Adv. Exp. Med. Biol. 1995, 380, 371–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlasak, R.; Luytjes, W.; Spaan, W.; Palese, P. Human and bovine coronaviruses recognize sialic acid-containing receptors similar to those of influenza C viruses. Proc. Natl. Acad. Sci. USA 1988, 85, 4526–4529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Dong, W.; Milewska, A.; Golda, A.; Qi, Y.; Zhu, Q.K.; Marasco, W.A.; Baric, R.S.; Sims, A.C.; Pyrc, K.; et al. Human Coronavirus HKU1 Spike Protein Uses O-Acetylated Sialic Acid as an Attachment Receptor Determinant and Employs Hemagglutinin-Esterase Protein as a Receptor-Destroying Enzyme. J. Virol. 2015, 89, 7202–7213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortorici, M.A.; Walls, A.C.; Lang, Y.; Wang, C.; Li, Z.; Koerhuis, D.; Boons, G.J.; Bosch, B.J.; Rey, F.A.; de Groot, R.J.; et al. Structural basis for human coronavirus attachment to sialic acid receptors. Nat. Struct. Mol. Biol. 2019, 26, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.R. HLA class I antigen serves as a receptor for human coronavirus OC43. Immunol. Investig. 1993, 22, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Owczarek, K.; Szczepanski, A.; Milewska, A.; Baster, Z.; Rajfur, Z.; Sarna, M.; Pyrc, K. Early events during human coronavirus OC43 entry to the cell. Sci. Rep. 2018, 8, 7124. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.P.; Almasy, K.M.; McDonald, E.F.; Plate, L. Comparative Multiplexed Interactomics of SARS-CoV-2 and Homologous Coronavirus Nonstructural Proteins Identifies Unique and Shared Host-Cell Dependencies. ACS Infect. Dis. 2020, 6, 3174–3189. [Google Scholar] [CrossRef]
- Hoffmann, H.H.; Sanchez-Rivera, F.J.; Schneider, W.M.; Luna, J.M.; Soto-Feliciano, Y.M.; Ashbrook, A.W.; Le Pen, J.; Leal, A.A.; Ricardo-Lax, I.; Michailidis, E.; et al. Functional interrogation of a SARS-CoV-2 host protein interactome identifies unique and shared coronavirus host factors. Cell Host Microbe 2020, 29, 267–280.e5. [Google Scholar] [CrossRef]
- Wang, R.; Simoneau, C.R.; Kulsuptrakul, J.; Bouhaddou, M.; Travisano, K.A.; Hayashi, J.M.; Carlson-Stevermer, J.; Zengel, J.R.; Richards, C.M.; Fozouni, P.; et al. Genetic Screens Identify Host Factors for SARS-CoV-2 and Common Cold Coronaviruses. Cell 2021, 184, 106–119.e114. [Google Scholar] [CrossRef]
- Schneider, W.M.; Luna, J.M.; Hoffmann, H.H.; Sanchez-Rivera, F.J.; Leal, A.A.; Ashbrook, A.W.; Le Pen, J.; Ricardo-Lax, I.; Michailidis, E.; Peace, A.; et al. Genome-Scale Identification of SARS-CoV-2 and Pan-coronavirus Host Factor Networks. Cell 2021, 184, 120–132.e114. [Google Scholar] [CrossRef]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R. Interferon gamma potentiates human coronavirus OC43 infection of neuronal cells by modulation of HLA class I expression. Immunol. Investig. 1995, 24, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.W.; Stephenson, K.B.; Mahony, J.; Lichty, B.D. Human coronavirus OC43 nucleocapsid protein binds microRNA 9 and potentiates NF-kappaB activation. J. Virol. 2014, 88, 54–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Guo, F.; Liu, F.; Cuconati, A.; Chang, J.; Block, T.M.; Guo, J.T. Interferon induction of IFITM proteins promotes infection by human coronavirus OC43. Proc. Natl. Acad. Sci. USA 2014, 111, 6756–6761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loo, S.L.; Wark, P.A.B.; Esneau, C.; Nichol, K.S.; Hsu, A.C.; Bartlett, N.W. Human coronaviruses 229E and OC43 replicate and induce distinct antiviral responses in differentiated primary human bronchial epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L926–L931. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.; Klumpp-Thomas, C.; Kalish, H.; Shunmugavel, A.; Mehalko, J.; Denson, J.P.; Snead, K.; Drew, M.; Corbett, K.; Graham, B.; et al. Serologic cross-reactivity of SARS-CoV-2 with endemic and seasonal Betacoronaviruses. medRxiv 2020. [Google Scholar] [CrossRef]
- Dugas, M.; Grote-Westrick, T.; Vollenberg, R.; Lorentzen, E.; Brix, T.; Schmidt, H.; Tepasse, P.R.; Kuhn, J. Less severe course of COVID-19 is associated with elevated levels of antibodies against seasonal human coronaviruses OC43 and HKU1 (HCoV OC43, HCoV HKU1). Int. J. Infect. Dis. 2021, 105, 304–306. [Google Scholar] [CrossRef]
- Saletti, G.; Gerlach, T.; Jansen, J.M.; Molle, A.; Elbahesh, H.; Ludlow, M.; Li, W.; Bosch, B.J.; Osterhaus, A.; Rimmelzwaan, G.F. Older adults lack SARS CoV-2 cross-reactive T lymphocytes directed to human coronaviruses OC43 and NL63. Sci. Rep. 2020, 10, 21447. [Google Scholar] [CrossRef]
- Lee, C.H.; Pinho, M.P.; Buckley, P.R.; Woodhouse, I.B.; Ogg, G.; Simmons, A.; Napolitani, G.; Koohy, H. Potential CD8+ T Cell Cross-Reactivity Against SARS-CoV-2 Conferred by Other Coronavirus Strains. Front. Immunol. 2020, 11, 579480. [Google Scholar] [CrossRef]
- Patrick, D.M.; Petric, M.; Skowronski, D.M.; Guasparini, R.; Booth, T.F.; Krajden, M.; McGeer, P.; Bastien, N.; Gustafson, L.; Dubord, J.; et al. An Outbreak of Human Coronavirus OC43 Infection and Serological Cross-reactivity with SARS Coronavirus. Can. J. Infect. Dis. Med. Microbiol. 2006, 17, 330–336. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e1415. [Google Scholar] [CrossRef] [PubMed]
- Arbour, N.; Ekande, S.; Cote, G.; Lachance, C.; Chagnon, F.; Tardieu, M.; Cashman, N.R.; Talbot, P.J. Persistent infection of human oligodendrocytic and neuroglial cell lines by human coronavirus 229E. J. Virol. 1999, 73, 3326–3337.e1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbour, N.; Cote, G.; Lachance, C.; Tardieu, M.; Cashman, N.R.; Talbot, P.J. Acute and persistent infection of human neural cell lines by human coronavirus OC43. J. Virol. 1999, 73, 3338–3350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.R.; Sorensen, O. Regulation of viral persistence in human glioblastoma and rhabdomyosarcoma cells infected with coronavirus OC43. Microb. Pathog. 1986, 1, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Bonavia, A.; Arbour, N.; Yong, V.W.; Talbot, P.J. Infection of primary cultuRes. of human neural cells by human coronaviruses 229E and OC43. J. Virol. 1997, 71, 800–806. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.; Mims, C.A. Differential susceptibility of cultured neural cells to the human coronavirus OC43. J. Virol. 1985, 53, 1016–1019. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.A.; Denis, F.; Talbot, P.J. Activation of glial cells by human coronavirus OC43 infection. J. Neuroimmunol. 2000, 108, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Jacomy, H.; Talbot, P.J. HCoV-OC43-induced apoptosis of murine neuronal cells. Adv. Exp. Med. Biol. 2006, 581, 473–478. [Google Scholar] [CrossRef]
- Desforges, M.; Le Coupanec, A.; Dubeau, P.; Bourgouin, A.; Lajoie, L.; Dube, M.; Talbot, P.J. Human Coronaviruses and Other Respiratory Viruses: Underestimated Opportunistic Pathogens of the Central Nervous System? Viruses 2019, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacomy, H.; St-Jean, J.R.; Brison, E.; Marceau, G.; Desforges, M.; Talbot, P.J. Mutations in the spike glycoprotein of human coronavirus OC43 modulate disease in BALB/c mice from encephalitis to flaccid paralysis and demyelination. J. Neurovirol. 2010, 16, 279–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meessen-Pinard, M.; Le Coupanec, A.; Desforges, M.; Talbot, P.J. Pivotal Role of Receptor-Interacting Protein Kinase 1 and Mixed Lineage Kinase Domain-Like in Neuronal Cell Death Induced by the Human Neuroinvasive Coronavirus OC43. J. Virol. 2017, 91, e01513-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, J.; Mims, C.A. Selective vulnerability of neural cells and age-related susceptibility to OC43 virus in mice. Arch. Virol. 1983, 77, 109–118. [Google Scholar] [CrossRef]
- Jacomy, H.; Talbot, P.J. Susceptibility of murine CNS to OC43 infection. Adv. Exp. Med. Biol. 2001, 494, 101–107. [Google Scholar] [CrossRef]
- Dube, M.; Le Coupanec, A.; Wong, A.H.M.; Rini, J.M.; Desforges, M.; Talbot, P.J. Axonal Transport Enables Neuron-to-Neuron Propagation of Human Coronavirus OC43. J. Virol. 2018, 92, e00404-18. [Google Scholar] [CrossRef] [Green Version]
- Butler, N.; Pewe, L.; Trandem, K.; Perlman, S. HCoV-OC43-induced encephalitis is in part immune-mediated. Adv. Exp. Med. Biol. 2006, 581, 531–534. [Google Scholar]
- Favreau, D.J.; Desforges, M.; St-Jean, J.R.; Talbot, P.J. A human coronavirus OC43 variant harboring persistence-associated mutations in the S glycoprotein differentially induces the unfolded protein response in human neurons as compared to wild-type virus. Virology 2009, 395, 255–267. [Google Scholar] [CrossRef]
- Niu, J.; Shen, L.; Huang, B.; Ye, F.; Zhao, L.; Wang, H.; Deng, Y.; Tan, W. Non-invasive bioluminescence imaging of HCoV-OC43 infection and therapy in the central nervous system of live mice. Antivir. Res. 2020, 173, 104646. [Google Scholar] [CrossRef]
- Le Coupanec, A.; Desforges, M.; Kaufer, B.; Dubeau, P.; Cote, M.; Talbot, P.J. Potential differences in cleavage of the S protein and type-1 interferon together control human coronavirus infection, propagation, and neuropathology within the central nervous system. J. Virol. 2021, 95, e00140-21. [Google Scholar] [CrossRef]
- Yeh, E.A.; Collins, A.; Cohen, M.E.; Duffner, P.K.; Faden, H. Detection of coronavirus in the central nervous system of a child with acute disseminated encephalomyelitis. Pediatrics 2004, 113, e73–e76. [Google Scholar] [CrossRef] [Green Version]
- Morfopoulou, S.; Brown, J.R.; Davies, E.G.; Anderson, G.; Virasami, A.; Qasim, W.; Chong, W.K.; Hubank, M.; Plagnol, V.; Desforges, M.; et al. Human Coronavirus OC43 Associated with Fatal Encephalitis. N. Engl. J. Med. 2016, 375, 497–498. [Google Scholar] [CrossRef]
- Kasereka, M.C.; Hawkes, M.T. Neuroinvasive potential of human coronavirus OC43: Case report of fatal encephalitis in an immunocompromised host. J. Neurovirol. 2021, 27, 340–344. [Google Scholar] [CrossRef]
- Arbour, N.; Day, R.; Newcombe, J.; Talbot, P.J. Neuroinvasion by human respiratory coronaviruses. J. Virol. 2000, 74, 8913–8921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovanec, D.L.; Flanagan, T.D. Detection of antibodies to human coronaviruses 229E and OC43 in the sera of multiple sclerosis patients and normal subjects. Infect. Immun. 1983, 41, 426–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debiaggi, M.; Perduca, M.; Romero, E.; Cereda, P.M. Phosphatidyl-serine inhibition of OC43 and NCDCV coronavirus infectivity. Microbiologica 1985, 8, 313–317. [Google Scholar] [PubMed]
- Collins, A.R.; Grubb, A. Inhibitory effects of recombinant human cystatin C on human coronaviruses. Antimicrob. Agents Chemother. 1991, 35, 2444–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.R.; Grubb, A. Cystatin D, a natural salivary cysteine protease inhibitor, inhibits coronavirus replication at its physiologic concentration. Oral Microbiol. Immunol. 1998, 13, 59–61. [Google Scholar] [CrossRef]
- Shen, L.; Yang, Y.; Ye, F.; Liu, G.; Desforges, M.; Talbot, P.J.; Tan, W. Safe and Sensitive Antiviral Screening Platform Based on Recombinant Human Coronavirus OC43 Expressing the Luciferase Reporter Gene. Antimicrob. Agents Chemother. 2016, 60, 5492–5503. [Google Scholar] [CrossRef] [Green Version]
- Keyaerts, E.; Li, S.; Vijgen, L.; Rysman, E.; Verbeeck, J.; Van Ranst, M.; Maes, P. Antiviral activity of chloroquine against human coronavirus OC43 infection in newborn mice. Antimicrob. Agents Chemother. 2009, 53, 3416–3421. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Wang, K.; Yu, W.; Sun, B.; Schwarz, W. Emodin inhibits current through SARS-associated coronavirus 3a protein. Antivir. Res. 2011, 90, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Brison, E.; Jacomy, H.; Desforges, M.; Talbot, P.J. Novel Treatment with Neuroprotective and Antiviral Properties against a Neuroinvasive Human Respiratory Virus. J. Virol. 2014, 88, 1548–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milewska, A.; Kaminski, K.; Ciejka, J.; Kosowicz, K.; Zeglen, S.; Wojarski, J.; Nowakowska, M.; Szczubialka, K.; Pyrc, K. HTCC: Broad Range Inhibitor of Coronavirus Entry. PLoS ONE 2016, 11, e0156552. [Google Scholar] [CrossRef] [Green Version]
- Milewska, A.; Chi, Y.; Szczepanski, A.; Barreto-Duran, E.; Dabrowska, A.; Botwina, P.; Obloza, M.; Liu, K.; Liu, D.; Guo, X.; et al. HTCC as a Polymeric Inhibitor of SARS-CoV-2 and MERS-CoV. J. Virol. 2021, 95, e01622-20. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Niu, J.; Wang, C.; Huang, B.; Wang, W.; Zhu, N.; Deng, Y.; Wang, H.; Ye, F.; Cen, S.; et al. High-Throughput Screening and Identification of Potent Broad-Spectrum Inhibitors of Coronaviruses. J. Virol. 2019, 93, e00023-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.H.; Kwon, S. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules 2019, 9, 696. [Google Scholar] [CrossRef] [Green Version]
- Zafferani, M.; Haddad, C.; Luo, L.; Davila-Calderon, J.; Yuan-Chiu, L.; Shema Mugisha, C.; Monaghan, A.G.; Kennedy, A.A.; Yesselman, J.D.; Gifford, R.R.; et al. Amilorides inhibit SARS-CoV-2 replication in vitro by targeting RNA structures. bioRxiv 2020. [Google Scholar] [CrossRef]
- Yang, C.W.; Lee, Y.Z.; Hsu, H.Y.; Jan, J.T.; Lin, Y.L.; Chang, S.Y.; Peng, T.T.; Yang, R.B.; Liang, J.J.; Liao, C.C.; et al. Inhibition of SARS-CoV-2 by Highly Potent Broad-Spectrum Anti-Coronaviral Tylophorine-Based Derivatives. Front. Pharm. 2020, 11, 606097. [Google Scholar] [CrossRef]
- Min, J.S.; Kim, D.E.; Jin, Y.H.; Kwon, S. Kurarinone Inhibits HCoV-OC43 Infection by Impairing the Virus-Induced Autophagic Flux in MRC-5 Human Lung Cells. J. Clin. Med. 2020, 9, 2230. [Google Scholar] [CrossRef]
- Bleasel, M.D.; Peterson, G.M. Emetine, Ipecac, Ipecac Alkaloids and Analogues as Potential Antiviral Agents for Coronaviruses. Pharmaceuticals 2020, 13, 51. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Peng, T.T.; Hsu, H.Y.; Lee, Y.Z.; Wu, S.H.; Lin, W.H.; Ke, Y.Y.; Hsu, T.A.; Yeh, T.K.; Huang, W.Z.; et al. Repurposing old drugs as antiviral agents for coronaviruses. Biomed. J. 2020, 43, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Marcello, A.; Civra, A.; Milan Bonotto, R.; Nascimento Alves, L.; Rajasekharan, S.; Giacobone, C.; Caccia, C.; Cavalli, R.; Adami, M.; Brambilla, P.; et al. The cholesterol metabolite 27-hydroxycholesterol inhibits SARS-CoV-2 and is markedly decreased in COVID-19 patients. Redox Biol. 2020, 36, 101682. [Google Scholar] [CrossRef] [PubMed]
- Good, S.S.; Westover, J.; Jung, K.H.; Zhou, X.J.; Moussa, A.; La Colla, P.; Collu, G.; Canard, B.; Sommadossi, J.P. AT-527, a double prodrug of a guanosine nucleotide analog, is a potent inhibitor of SARS-CoV-2 in vitro and a promising oral antiviral for treatment of COVID-19. Antimicrob. Agents Chemother. 2021, 65, e02479-20. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Park, R.; Park, Y.I.; Cha, Y.E.; Yamamoto, A.; Lee, J.I.; Park, J. EGCG, a green tea polyphenol, inhibits human coronavirus replication in vitro. Biochem. Biophys. Res. Commun. 2021, 547, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Meng, X.; Zhang, F.; Xiang, Y.; Wang, J. The in vitro antiviral activity of lactoferrin against common human coronaviruses and SARS-CoV-2 is mediated by targeting the heparan sulfate co-receptor. Emerg. Microbes Infect. 2021, 10, 317–330. [Google Scholar] [CrossRef] [PubMed]
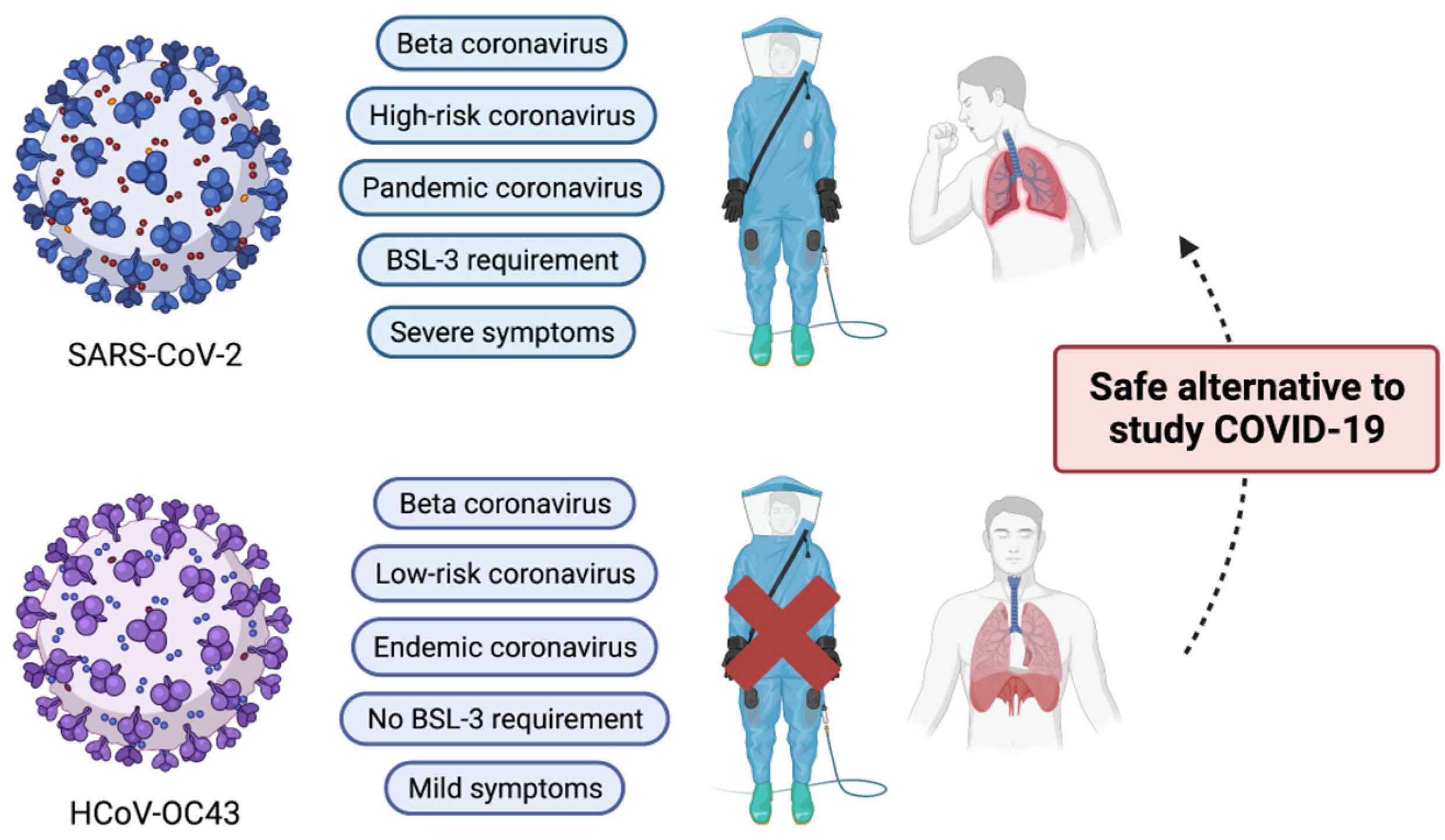
Figure 1.
Comparison of major characteristics of SARS-CoV-2 and HCoV-OC43. Despite belonging to the same beta coronavirus, HCoV-OC43 is classified as an endemic coronavirus. In addition, due to its mild clinical symptoms, its biology can be studied under the BSL-2 research setting. Therefore, HCoV-OC43 can serve as a good research alternative to SARS-CoV-2. This illustration was created with BioRender.com.
Figure 1.
Comparison of major characteristics of SARS-CoV-2 and HCoV-OC43. Despite belonging to the same beta coronavirus, HCoV-OC43 is classified as an endemic coronavirus. In addition, due to its mild clinical symptoms, its biology can be studied under the BSL-2 research setting. Therefore, HCoV-OC43 can serve as a good research alternative to SARS-CoV-2. This illustration was created with BioRender.com.

Figure 2.
Comparison of abbreviated genomic maps of HCoV-OC43 with other coronaviruses including HCoV-229E, HCoV-NL63, HCoV-HKU, MERS-CoV, and SARS-CoV-2 (not to the scale). Used acronyms are as follows. ORF; open reading frame, HE; hemagglutinin-esterase, S; spike, E; envelope, M; membrane, N; nucleocapsid. Number-only genes (2a, 5a, 3a, 3b, 3, 4, 5, 6, 7, 8, 10) indicate nonstructural genes. Some in-between nonstructural genes from SARS-CoV-2 were not shown for purposes of better comparison.
Figure 2.
Comparison of abbreviated genomic maps of HCoV-OC43 with other coronaviruses including HCoV-229E, HCoV-NL63, HCoV-HKU, MERS-CoV, and SARS-CoV-2 (not to the scale). Used acronyms are as follows. ORF; open reading frame, HE; hemagglutinin-esterase, S; spike, E; envelope, M; membrane, N; nucleocapsid. Number-only genes (2a, 5a, 3a, 3b, 3, 4, 5, 6, 7, 8, 10) indicate nonstructural genes. Some in-between nonstructural genes from SARS-CoV-2 were not shown for purposes of better comparison.


{kind=link}
{kind=link}
Table 1.
Major characteristics of structural proteins of HCoV-OC43.
| Structural Protein | Key Function | References |
|---|---|---|
| Hemagglutinin-esterase | Receptor-binding and degradation | [52,53,54,55,56] |
| Spike protein | Receptor-binding and hemagglutination | [61] |
| Envelope protein | Ion channel formation | [62] |
| Membrane protein | Virion morphogenesis | [45] |
| Nucleocapsid | Helical nucleocapsid formation | [1] |
Table 2.
Host factors or processes identified necessary for the HCoV-OC43 life cycle.
| Host Factor or Process | Steps of Virus Life Cycle | References |
|---|---|---|
| 9-O-acetyl sialic acid | Host receptor binding | [6,82,83,84] |
| MHC class I | Host receptor binding | [18,85] |
| Caveolin-1-dependent endocytosis | Virus entry | [86] |
| ER structure | Virus RNA replication | [87] |
| Rab GTPase and glycosylphosphatidylinositol | Virus assembly and trafficking | [88] |
| Endosome maturation, phosphatidylinositol phosphate, and cholesterol homeostasis | Virus assembly and trafficking | [89] |
| VMP1, TMEM41B, and TMEM64 | ER membrane remodeling | [90] |
| Dynamin-dependent budding | Virus exit | [86] |
Table 3.
Anti-coronavirus candidate molecules identified by using the HCoV-OC43 infection model.
| Name | Experiment Type | EC50 (mM) * | References |
|---|---|---|---|
| Phosphatidyl-serine | in vitro | na | [126] |
| Cystatin C and D | in vitro | 0.8 | [127,128] |
| Chloroquine | in vitro and in vivo | 0.33 | [129] |
| Emodin | in vitro (inhibition of 3a ion channel) | <10 | [131] |
| Memantine | in vivo | NA ** | [132] |
| HTCC | in vitro | NA | [133] |
| Lycorine | in vivo | <5 | [135] |
| Bis-benzylisoquinoline alkaloids | in vitro | <0.1 | [136] |
| Amiloride | in vitro | >10 | [137] |
| Tylophorine-based compounds | in vitro | 0.1–1 | [138] |
| Cardenolides and bufadienolides | in vitro | 0.1–1 | [138] |
| Kurarinone | in vitro | 3.458 | [139] |
| Emetine | in vitro | 0.21 | [140,141] |
| Oxysterol 27-hydroxycholesterol | in vitro | <10 | [142] |
| Valinomycin | in vitro | 6.15 | [40] |
| AT-527 | in vitro | 2.2 | [143] |
| EGCG | in vitro | 14.6 | [144] |
| Lactoferrin | in vitro | <50 mg/mL | [145] |
* Effective concentration to reduce the amount of virus proliferation in half. ** Not available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, M.I.; Lee, C. Human Coronavirus OC43 as a Low-Risk Model to Study COVID-19. Viruses 2023, 15, 578. https://doi.org/10.3390/v15020578
AMA Style
Kim MI, Lee C. Human Coronavirus OC43 as a Low-Risk Model to Study COVID-19. Viruses. 2023; 15(2):578. https://doi.org/10.3390/v15020578
Chicago/Turabian StyleKim, Mi Il, and Choongho Lee. 2023. "Human Coronavirus OC43 as a Low-Risk Model to Study COVID-19" Viruses 15, no. 2: 578. https://doi.org/10.3390/v15020578
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.