
Transcriptional Reprogramming of Autographa Californica Multiple Nucleopolyhedrovirus Chitinase and Cathepsin Genes Enhances Virulence


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cells, Viruses, and Insects
2.2. RNA Isolation, Electrophoresis, and Blotting
2.3. Northern Blot Hybridization and Analysis
2.4. Temporal CHIA and V-CATH Protein Analysis
2.5. Chitinase and Cathepsin Activity Assays
3. Results
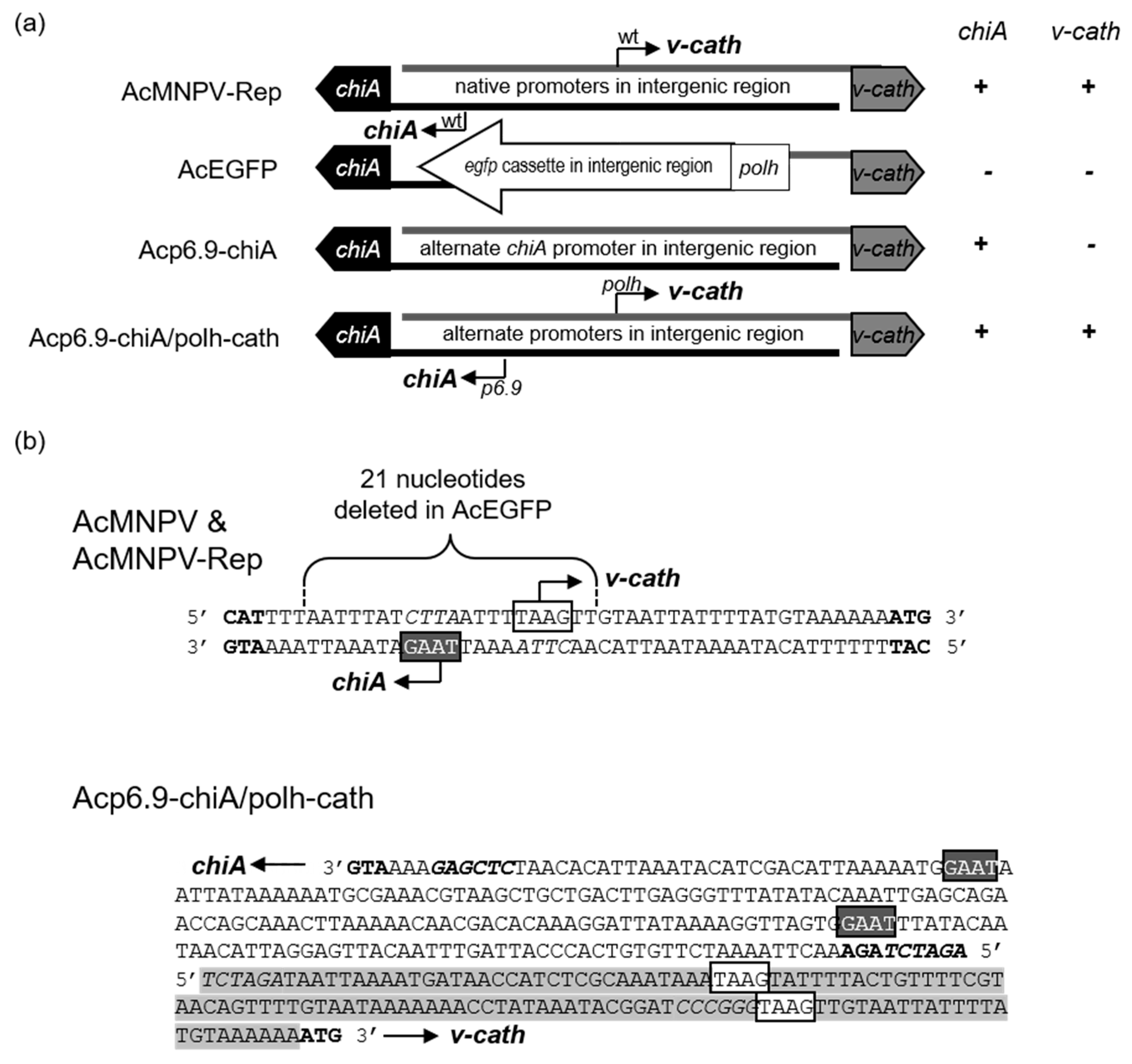
3.1. Dual Reprogramming of chiA and v-cath Expression
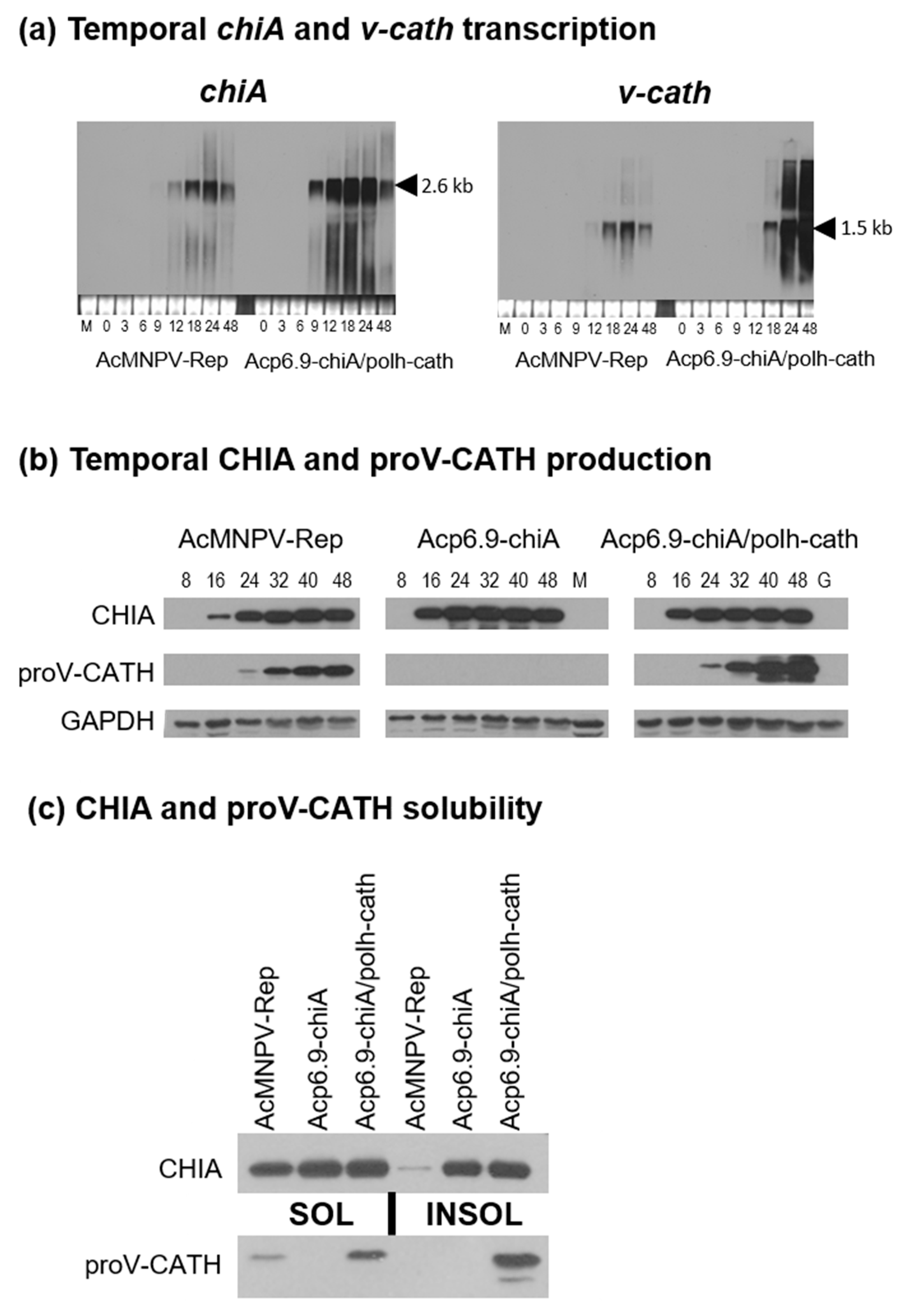
3.2. CHIA and proV-CATH Protein Expression Profiles from Acp6.9-chiA/polh-cath
3.3. Dual Transcriptional Reprogramming Increases CHIA and V-CATH Enzymatic Activity
3.4. Dual chiA/v-cath Reprogramming Does Not Affect Virus Replication
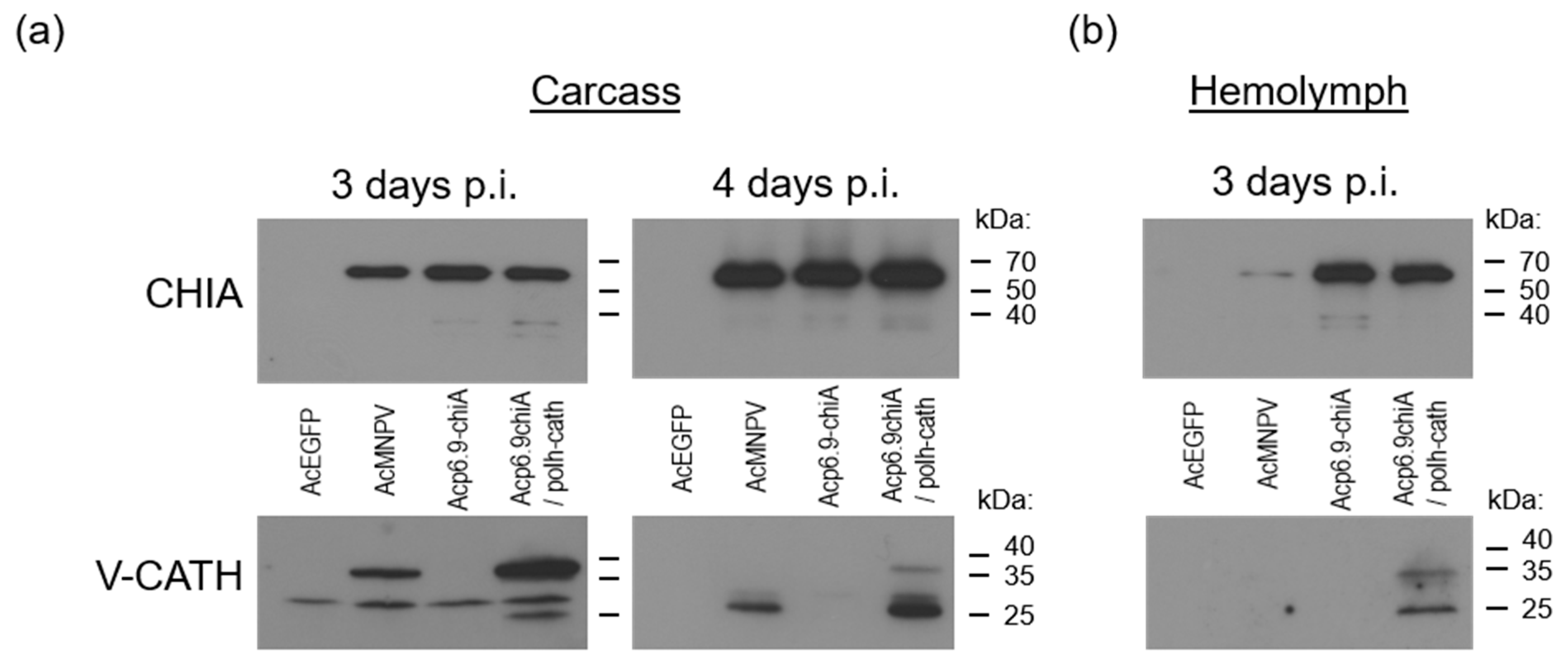
3.5. CHIA and V-CATH Expression in Infected Larvae
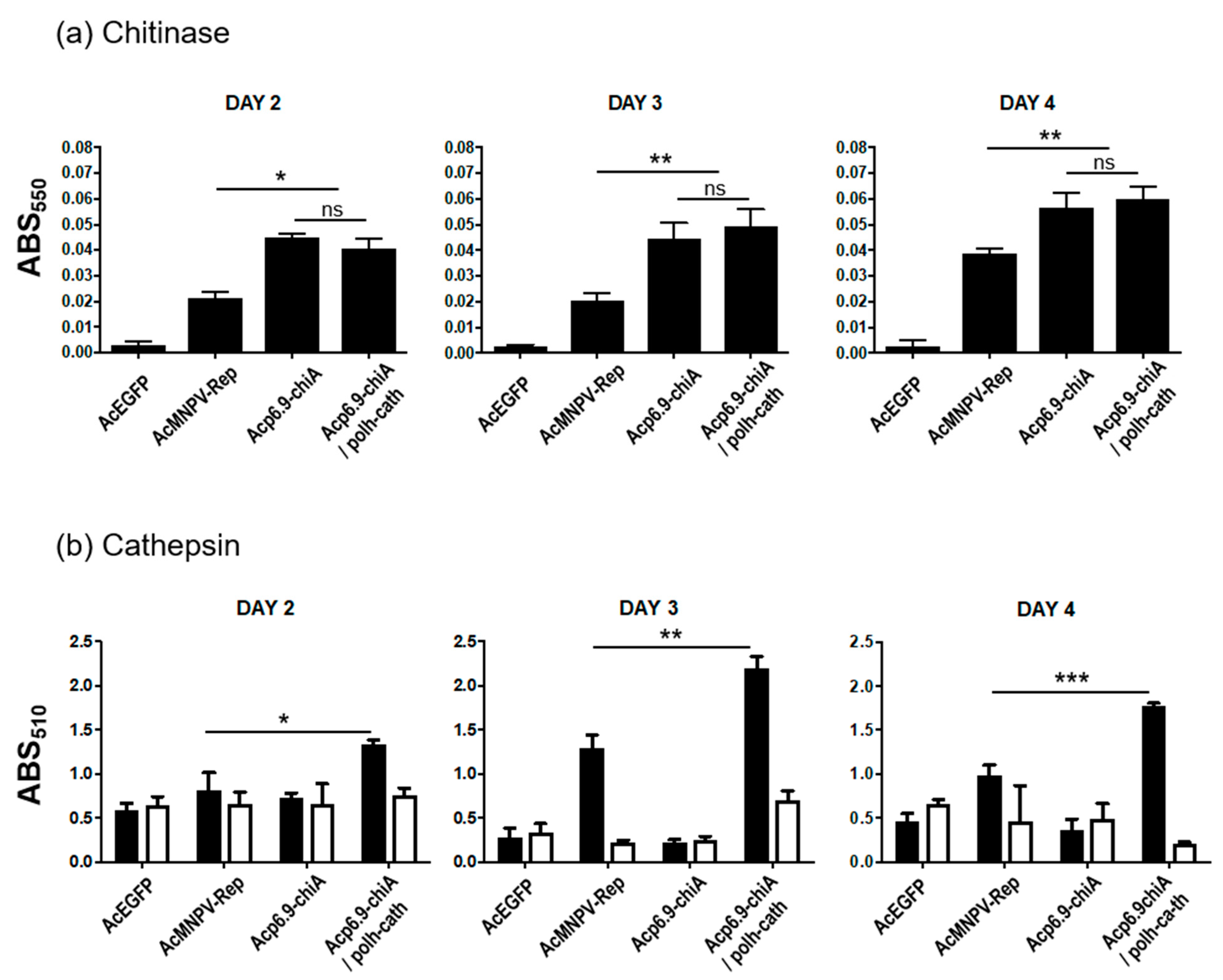
3.6. CHIA and V-CATH Enzyme Activity in Infected Larvae
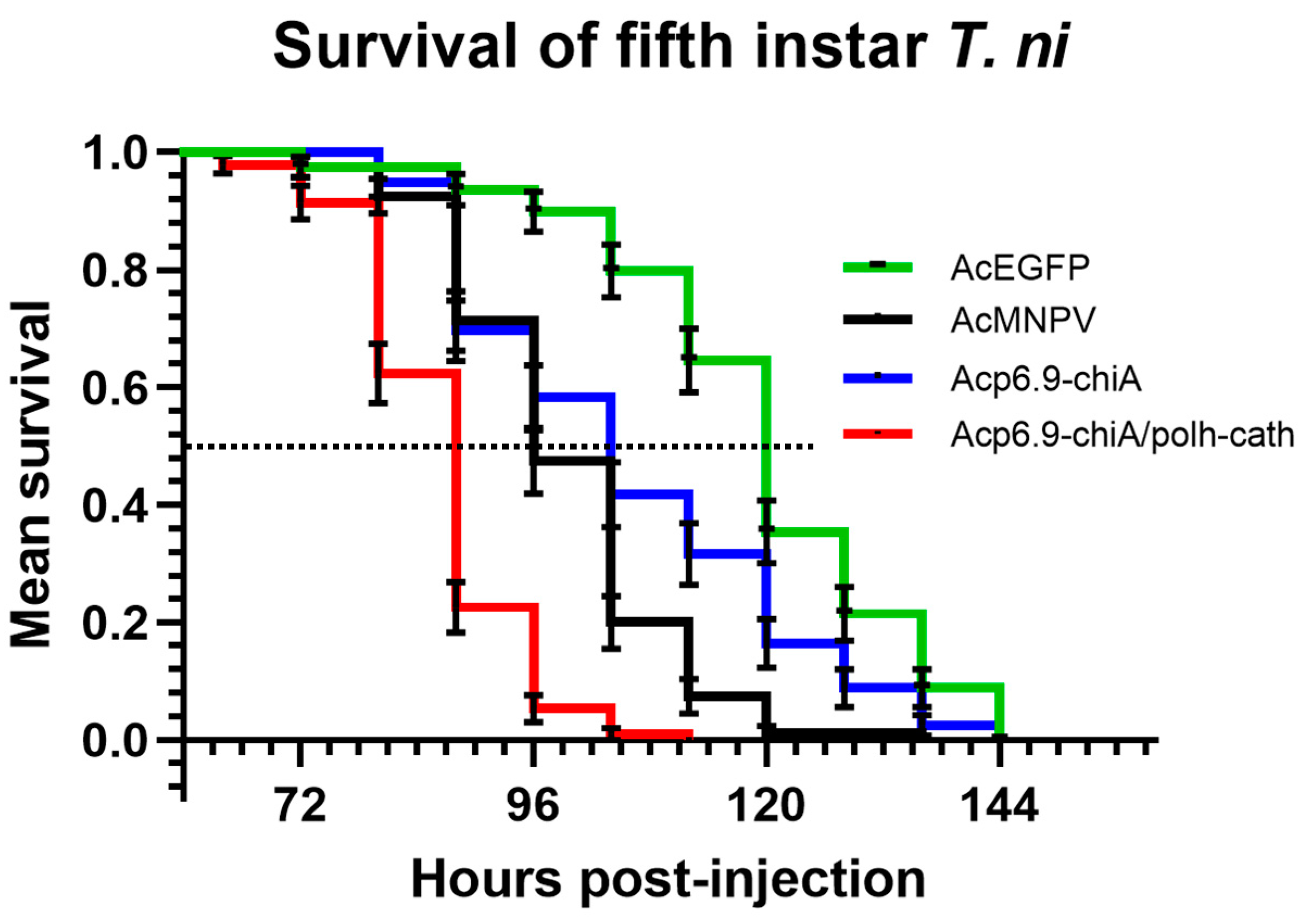
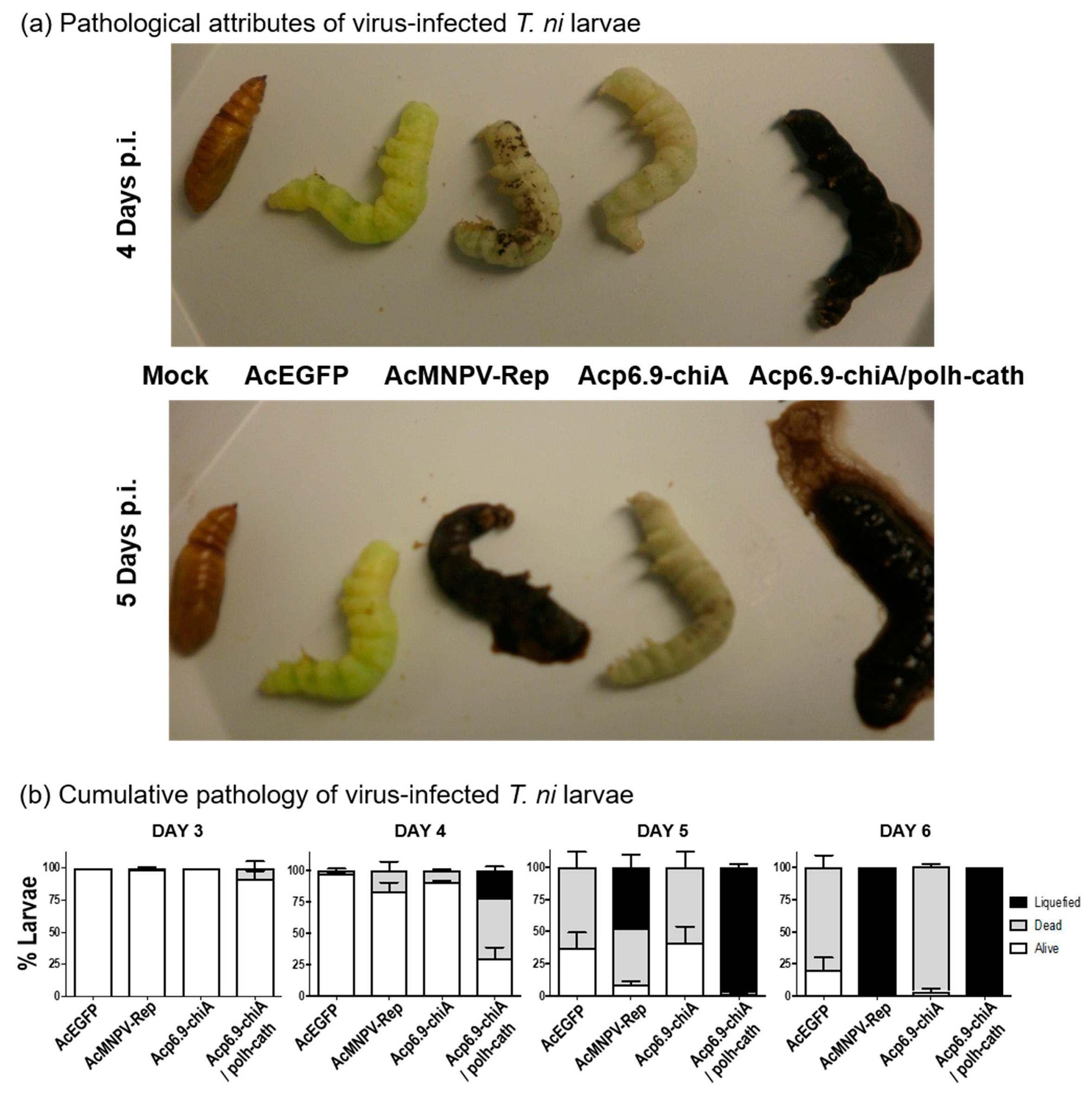
3.7. Dual chiA/v-cath Reprogramming Increases Host Mortality
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawtin, R.E.; Zarkowska, T.; Arnold, K.; Thomas, C.J.; Gooday, G.W.; King, L.A.; Kuzio, J.A.; Possee, R.D. Liquefaction of Autographa californica nucleopolyhedrovirus-infected insects is dependent on the integrity of virus-encoded chitinase and cathepsin genes. Virology 1997, 238, 243–253. [Google Scholar] [CrossRef]
- O’Reilly, D.R. Auxiliary Genes of Baculoviruses. In The Baculoviruses; Miller, L.K., Ed.; Springer: Boston, MA, USA, 1997. [Google Scholar] [CrossRef]
- Corrado, G.; Arciello, S.; Fanti, P.; Fiandra, L.; Garonna, A.; Digilio, M.C.; Lorito, M.; Giordana, B.; Pennacchio, F.; Rao, R. The Chitinase A from the baculovirus AcMNPV enhances resistance to both fungi and herbivorous pests in tobacco. Transgenic Res. 2008, 17, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.A.; Aragao, C.W.; de Castro, M.E.; Oliveira, J.V.; Sosa Gomez, D.R.; Ribeiro, B.M. A recombinant Anticarsia gemmatalis MNPV harboring chiA and v-cath genes from Choristoneura fumiferana defective NPV induce host liquefaction and increased insecticidal activity. PLoS ONE 2013, 8, e74592. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Choi, J.Y.; Roh, J.Y.; Tao, X.Y.; Liu, Q.; Lee, J.H.; Kim, J.S.; Kim, W.J.; Je, Y.H. Insecticidal activity of the chitinase from the Spodoptera litura nucleopolyhedrovirus. Entomol. Res. 2013, 43, 63–69. [Google Scholar] [CrossRef]
- Saville, G.P.; Patmanidi, A.L.; Possee, R.D.; King, L.A. Deletion of the Autographa californica nucleopolyhedrovirus chitinase KDEL motif and in vitro and in vivo analysis of the modified virus. J. Gen. Virol. 2004, 85, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Saville, G.P.; Thomas, C.J.; Possee, R.D.; King, L.A. Partial redistribution of the Autographa californica nucleopolyhedrovirus chitinase in virus-infected cells accompanies mutation of the carboxy-terminal KDEL ER-retention motif. J. Gen. Virol. 2002, 83, 685–694. [Google Scholar] [CrossRef]
- Gopalakrishnan, B.; Muthukrishnan, S.; Kramer, K.J. Baculovirus-Mediated Expression of a Manduca-Sexta Chitinase Gene-Properties of the Recombinant Protein. Insect Biochem. Molec. 1995, 25, 255–265. [Google Scholar] [CrossRef]
- Li, H.R.; Tang, H.L.; Sivakumar, S.; Philip, J.; Harrison, R.L.; Gatehouse, J.A.; Bonning, B.C. Insecticidal activity of a basement membrane-degrading protease against Heliothis virescens (Fabricius) and Acyrthosiphon pisum (Harris). J. Insect Physiol. 2008, 54, 777–789. [Google Scholar] [CrossRef]
- Slack, J.M.; Ribeiro, B.M.; de Souza, M.L. The gp64 locus of Anticarsia gemmatalis multicapsid nucleopolyhedrovirus contains a 3′ repair exonuclease homologue and lacks v-cath and ChiA genes. J. Gen. Virol. 2004, 85, 211–219. [Google Scholar] [CrossRef]
- Hodgson, J.J.; Arif, B.M.; Krell, P.J. Interaction of Autographa californica multiple nucleopolyhedrovirus cathepsin protease progenitor (proV-CATH) with insect baculovirus chitinase as a mechanism for proV-CATH cellular retention. J. Virol. 2011, 85, 3918–3929. [Google Scholar] [CrossRef]
- Hom, L.G.; Ohkawa, T.; Trudeau, D.; Volkman, L.E. Autographa californica M nucleopolyhedrovirus ProV-CATH is activated during infected cell death. Virology 2002, 296, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J.J.; Krell, P.J.; Passarelli, A.L. Mature viral cathepsin is required for release of viral occlusion bodies from Autographa californica multiple nucleopolyhedrovirus-infected cells. Virology 2021, 556, 23–32. [Google Scholar] [CrossRef]
- Daimon, T.; Katsuma, S.; Shimada, T. Mutational analysis of active site residues of chitinase from Bombyx mori nucleopolyhedrovirus. Virus Res. 2007, 124, 168–175. [Google Scholar] [CrossRef]
- Hom, L.G.; Volkman, L.E. Autographa californica M nucleopolyhedrovirus chiA is required for processing of V-CATH. Virology 2000, 277, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Katsuma, S.; Nakanishi, T.; Daimon, T.; Shimada, T. N-linked glycans located in the pro-region of Bombyx mori nucleopolyhedrovirus V-CATH are essential for the proper folding of V-CATH and V-CHIA. J. Gen. Virol. 2009, 90, 170–176. [Google Scholar] [CrossRef]
- Hodgson, J.J.; Arif, B.M.; Krell, P.J. Role of interactions between Autographa californica multiple nucleopolyhedrovirus procathepsin and chitinase chitin-binding or active-site domains in viral cathepsin processing. J. Virol. 2013, 87, 3471–3483. [Google Scholar] [CrossRef] [PubMed]
- Ishimwe, E.; Hodgson, J.J.; Clem, R.J.; Passarelli, A.L. Reaching the melting point: Degradative enzymes and protease inhibitors involved in baculovirus infection and dissemination. Virology 2015, 479, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J.J.; Arif, B.M.; Krell, P.J. Reprogramming the chiA expression profile of Autographa californica multiple nucleopolyhedrovirus. J. Gen. Virol. 2007, 88, 2479–2487. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J.J.; Arif, B.M.; Krell, P.J. Autographa californica multiple nucleopolyhedrovirus and Choristoneura fumiferana multiple nucleopolyhedrovirus v-cath genes are expressed as pre-proenzymes. J. Gen. Virol. 2009, 90, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, D.R. Use of baculovirus expression vectors. Methods Mol. Biol. 1997, 62, 235–246. [Google Scholar] [CrossRef]
- Hill, J.E.; Kuzio, J.; Faulkner, P. Identification and characterization of the v-cath gene of the baculovirus, CfMNPV. Gene Struct. Expr. 1995, 1264, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Daimon, T.; Katsuma, S.; Kang, W.K.; Shimada, T. Functional characterization of chitinase from Cydia pomonella granulovirus. Arch. Virol. 2007, 152, 1655–1664. [Google Scholar] [CrossRef]
- Katsuma, S.; Nakanishi, T.; Shimada, T. Bombyx mori nucleopolyhedrovirus FP25K is essential for maintaining a steady-state level of v-cath expression throughout the infection. Virus Res. 2009, 140, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.M.; Kuzio, J.; Faulkner, P. Characterization of v-cath, a cathepsin L-like proteinase expressed by the baculovirus Autographa californica multiple nuclear polyhedrosis virus. J. Gen. Virol. 1995, 76 Pt 5, 1091–1098. [Google Scholar] [CrossRef]
- Hom, L.G.; Volkman, L.E. Preventing proteolytic artifacts in the baculovirus expression system. Biotechniques 1998, 25, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Ishimwe, E.; Hodgson, J.J.; Passarelli, A.L. Expression of the Cydia pomonella granulovirus matrix metalloprotease enhances Autographa californica multiple nucleopolyhedrovirus virulence and can partially substitute for viral cathepsin. Virology 2015, 481, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Friesen, P.D.; Miller, L.K. Temporal regulation of baculovirus RNA: Overlapping early and late transcripts. J. Virol. 1985, 54, 392–400. [Google Scholar] [CrossRef]
- Ooi, B.G.; Rankin, C.; Miller, L.K. Downstream sequences augment transcription from the essential initiation site of a baculovirus polyhedrin gene. J. Mol. Biol. 1989, 210, 721–736. [Google Scholar] [CrossRef]
- Possee, R.D.; Howard, S.C. Analysis of the polyhedrin gene promoter of the Autographa californica nuclear polyhedrosis virus. Nucleic Acids Res. 1987, 15, 10233–10248. [Google Scholar] [CrossRef] [Green Version]
- Hawtin, R.E.; Arnold, K.; Ayres, M.D.; Zanotto, P.M.; Howard, S.C.; Gooday, G.W.; Chappell, L.H.; Kitts, P.A.; King, L.A.; Possee, R.D. Identification and preliminary characterization of a chitinase gene in the Autographa californica nuclear polyhedrosis virus genome. Virology 1995, 212, 673–685. [Google Scholar] [CrossRef]
- Ohkawa, T.; Majima, K.; Maeda, S. A cysteine protease encoded by the baculovirus Bombyx mori nuclear polyhedrosis virus. J. Virol. 1994, 68, 6619–6625. [Google Scholar] [CrossRef] [PubMed]
- Moscardi, F.; de Souza, M.L.; de Castro, M.E.B.; Moscardi, M.L.; Szewczyk, B. Baculovirus Pesticides: Present State and Future Perspectives. In Microbes and Microbial Technology: Agricultural and Environmental Applications; Springer: New York, NY, USA, 2011; pp. 415–445. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodgson, J.J.; Passarelli, A.L.; Krell, P.J. Transcriptional Reprogramming of Autographa Californica Multiple Nucleopolyhedrovirus Chitinase and Cathepsin Genes Enhances Virulence. Viruses 2023, 15, 503. https://doi.org/10.3390/v15020503
Hodgson JJ, Passarelli AL, Krell PJ. Transcriptional Reprogramming of Autographa Californica Multiple Nucleopolyhedrovirus Chitinase and Cathepsin Genes Enhances Virulence. Viruses. 2023; 15(2):503. https://doi.org/10.3390/v15020503
Chicago/Turabian StyleHodgson, Jeffrey J., A. Lorena Passarelli, and Peter J. Krell. 2023. "Transcriptional Reprogramming of Autographa Californica Multiple Nucleopolyhedrovirus Chitinase and Cathepsin Genes Enhances Virulence" Viruses 15, no. 2: 503. https://doi.org/10.3390/v15020503