
Human Metapneumovirus (hMPV) Infection and MPV467 Treatment in Immunocompromised Cotton Rats Sigmodon hispidus
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Viruses and Viral Assays
2.3. Animals and Animal Studies
2.4. Viral Titration
2.5. Lung Cytokine/Chemokine mRNA Analysis
2.6. Histolopathology Analysis
3. Results
3.1. Immunosuppression Results in Increased hMPV Replication and Delayed Viral Clearance in Cotton Rats
3.2. Prophylactic or Therapeutic Treatment with Anti-hMPV Antibody Reduces hMPV Load in Immunosuppressed Cotton Rats in a Dose-Dependent Manner
3.3. MPV467 Therapy Ameliorates Delayed hMPV Clearance in Immunosuppressed Cotton Rats
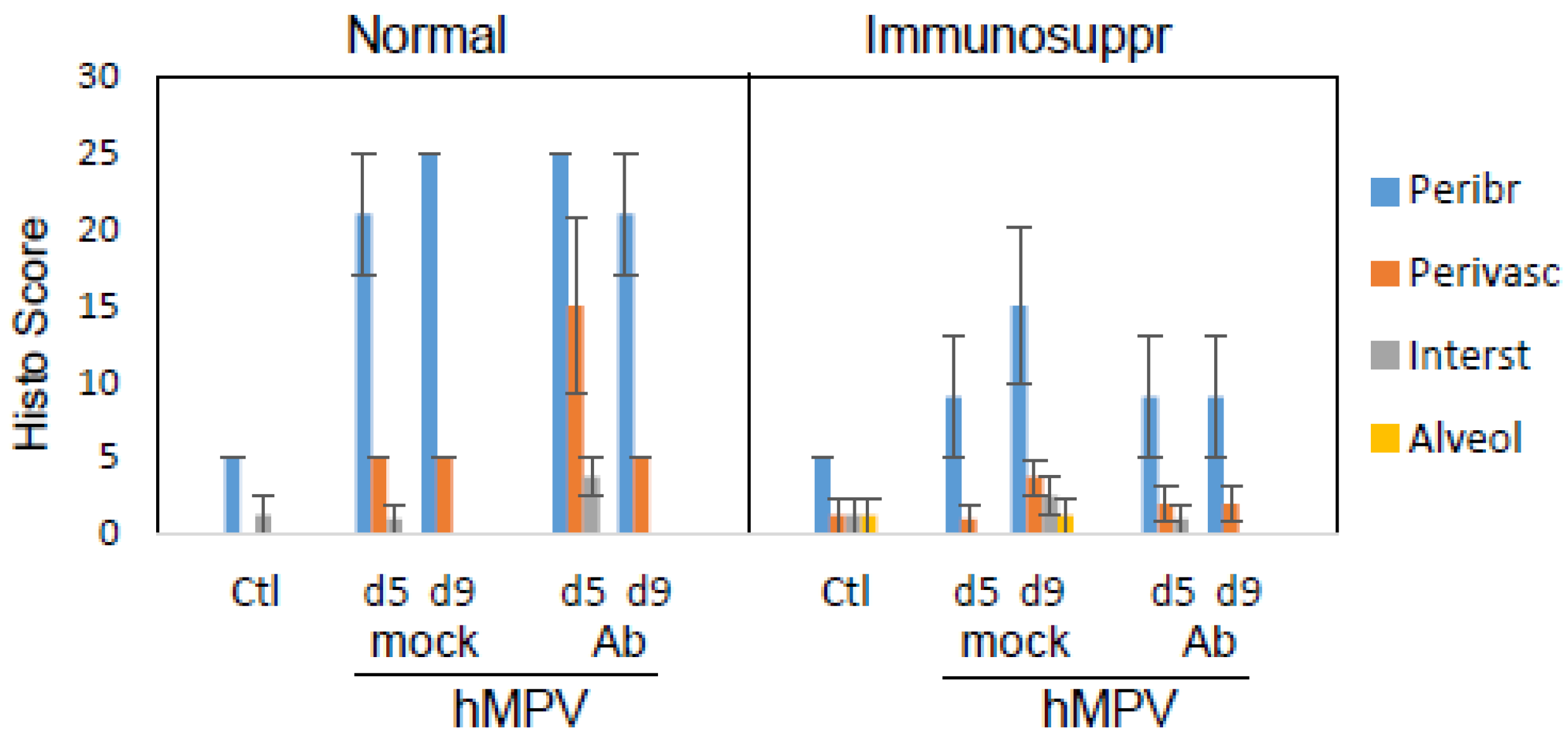
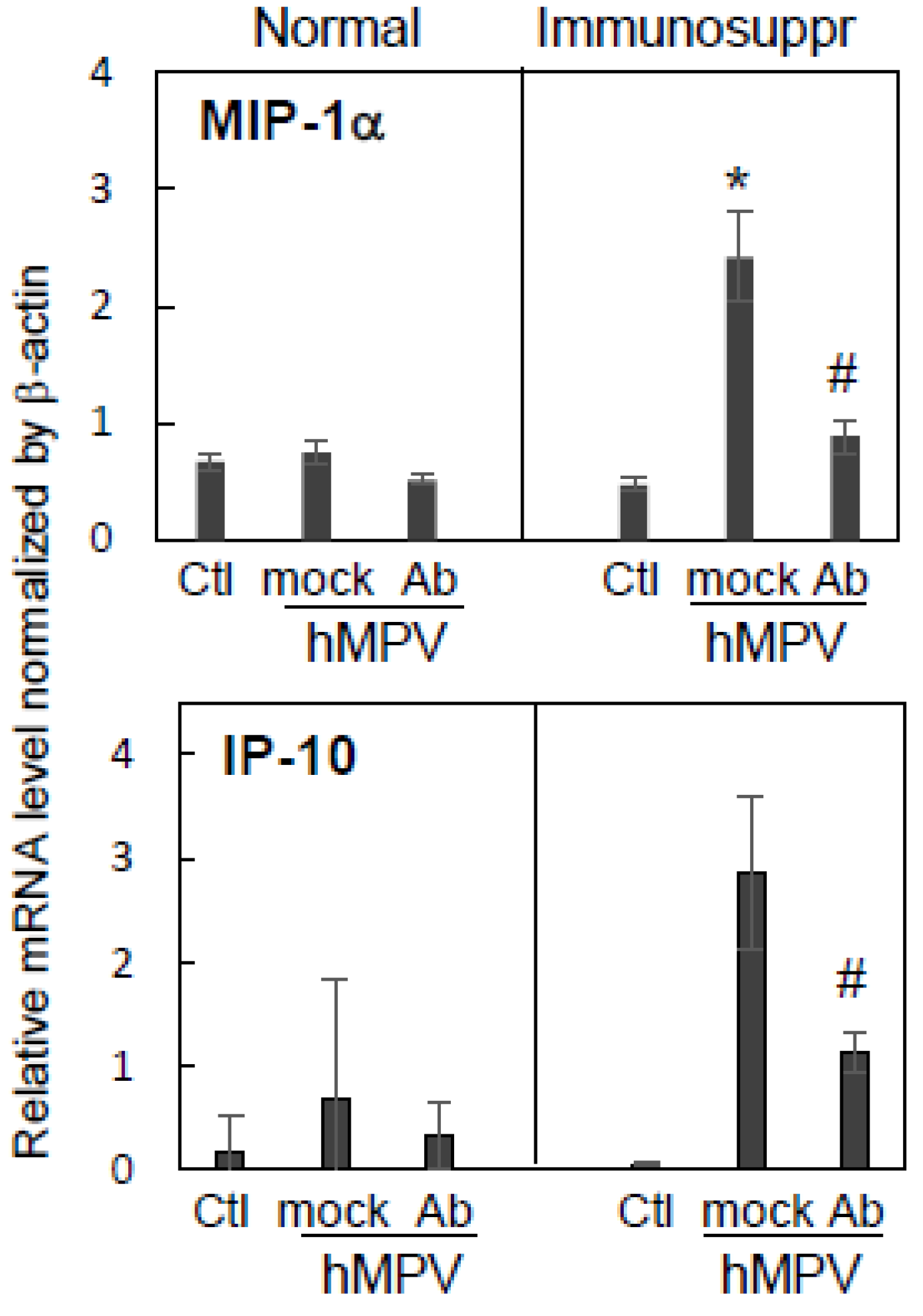
3.4. Effect of Antibody Therapy on Lung Histopathology and Chemokine Expression in Immunosuppressed Cotton Rats
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van den Hoogen, B.G.; de Jong, J.C.; Groen, J.; Kuiken, T.; de Groot, R.; Fouchier, R.A.; Osterhaus, A.D. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.; Turner, C.; Watthanaworawit, W.; Carrara, V.; Cicelia, N.; Deglise, C. Respiratory virus surveillance in hospitalised pneumonia patients on the Thailand–Myanmar border. BMC Infect. Dis. 2013, 13, 434. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Li, J.; Xie, Z.; Liu, C.; Guo, L.; Vernet, G. Human metapneumovirus associated with community-acquired pneumonia in children in Beijing, China. J. Med. Virol. 2013, 85, 138–143. [Google Scholar] [CrossRef]
- López-Huertas, M.R.; Casas, I.; Acosta-Herrera, B.; García, M.L.; Coiras, M.T.; Pérez-Breña, P. Two RT-PCR based assays to detect human metapneumovirus in nasopharyngeal aspirates. J. Virol. Methods 2005, 129, 1–7. [Google Scholar] [CrossRef] [PubMed]
- van den Hoogen, B.G.; Osterhaus, D.M.; Fouchier, R.A. Clinical impact and diagnosis of human metapneumovirus infection. Pediatr. Infect. Dis. J. 2004, 23 (Suppl. S1), S25–S32. [Google Scholar] [CrossRef]
- Edwards, K.M.; Zhu YGriffin, M.R.; Weinberg, G.A.; Hall, C.B.; Szilagyi, P.G. New vaccine surveillance network burden of human metapneumovirus infection in young children. N. Engl. J. Med. 2013, 368, 633–643. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreaks of human metapneumovirus in two skilled nursing facilities—West Virginia and Idaho, 2011–2012. Morb. Mortal. Wkly. Rep. (MMWR) 2013, 62, 909–913. [Google Scholar]
- Fontana, L.; Strasfeld, L. Respiratory Virus Infections of the Stem Cell Transplant Recipient and the Hematologic Malignancy Patient. Infect. Dis. Clin. North Am. 2019, 33, 523–544. [Google Scholar] [CrossRef]
- Martinez-Rodriguez, C.; Banos-Lara, M.D.R. HMPV in Immunocompromised Patients: Frequency and Severity in Pediatric Oncology Patients. Pathogens 2020, 9, 51. [Google Scholar] [CrossRef]
- Williams, J.V.; Harris, P.A.; Tollefson, S.J.; Halburnt-Rush, L.L.; Pingsterhaus, J.M.; Edwards, K.M.; Wright, P.F.; Crowe, J.E., Jr. Human Metapneumovirus and Lower Respiratory Tract Disease in Otherwise Healthy Infants and Children. N. Engl. J. Med. 2004, 350, 443–450. [Google Scholar] [CrossRef]
- Englund, J.A.; Boeckh, M.; Kuypers, J.; Nichols, W.G.; Hackman, R.C.; Morrow, R.A.; Fredricks, D.N.; Corey, L. Brief Communication: Fatal Human Metapneumovirus Infection in Stem-Cell Transplant Recipients. Ann. Intern. Med. 2006, 144, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Scheuerman, O.; Barkai, G.; Mandelboim, M.; Mishali, H.; Chodick, G.; Levy, I. Human metapneumovirus (hMPV) infection in immunocompromised children. J. Clin. Virol. 2016, 83, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Renaud, C.; Campbell, A. Changing epidemiology of respiratory viral infections in hematopoietic cell transplant recipients and solid organ transplant recipients. Curr. Opin. Infect. Dis. 2011, 24, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Renaud, C.; Xie, H.; Seo, S.; Kuypers, J.; Cent, A.; Corey, L.; Leisenring, W.; Boeckh, M.; Englund, J.A. Mortality Rates of Human Metapneumovirus and Respiratory Syncytial Virus Lower Respiratory Tract Infections in Hematopoietic Cell Transplantation Recipients. Biol. Blood Marrow Transplant. 2013, 19, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.Y.; Renaud, C.; Ficken, E.; Thomson, B.; Kuypers, J.; Englund, J.A. Respiratory Tract Infections Due to Human Metapneumovirus in Immunocompromised Children. J. Pediatric Infect. Dis. Soc. 2014, 3, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Debiaggi, M.; Canducci, F.; Sampaolo, M.; Marinozzi, M.C.; Parea, M.; Terulla, C.; Colombo, A.A.; Alessandrino, E.P.; Bragotti, L.Z.; Arghittu, M.; et al. Persistent Symptomless Human Metapneumovirus Infection in Hematopoietic Stem Cell Transplant Recipients. J. Infect. Dis. 2006, 194, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Debiaggi, M.; Canducci, F.; Terulla, C.; Sampaolo, M.; Marinozzi, M.C.; Alessandrino, P.E.; Colombo, A.A.; Caldera, D.; Bragotti, L.Z.; Migliavacca, R.; et al. Long-term study on symptomless human metapneumovirus infection in hematopoietic stem cell transplant recipients. New Microbiol. 2007, 30, 255–258. [Google Scholar]
- Blanco, J.C.G.; Boukhvalova, M.S.; Perez, D.R.; Vogel, S.N.; Kajon, A. Modeling Human Respiratory Viral Infections in the Cotton Rat (Sigmodon hispidus). J. Antivir. Antiretrovir. 2014, 6, 40–42. [Google Scholar] [CrossRef]
- Patel, M.C.; Wang, W.; Pletneva, L.M.; Rajagopala, S.V.; Tan, Y.; Hartert, T.V.; Boukhvalova, M.S.; Vogel, S.N.; Das, S.R.; Blanco, J.C.G. Enterovirus D-68 Infection, Prophylaxis, and Vaccination in a Novel Permissive Animal Model, the Cotton Rat (Sigmodon hispidus). PLoS ONE 2016, 11, e0166336. [Google Scholar] [CrossRef]
- Hamelin, M.-E.; Yim, K.; Kuhn, K.H.; Cragin, R.P.; Boukhvalova, M.; Blanco, J.C.G.; Prince, G.A.; Boivin, G. Pathogenesis of Human Metapneumovirus Lung Infection in BALB/c Mice and Cotton Rats. J. Virol. 2005, 79, 8894–8903. [Google Scholar] [CrossRef]
- Williams, J.V.; Tollefson, S.J.; Johnson, J.E.; Crowe, J.E., Jr. The cotton rat (Sigmodon hispidus) is a permissive small animal model of human metapneumovirus infection, pathogenesis, and protective immunity. J. Virol. 2005, 79, 10944–10951. [Google Scholar] [CrossRef] [PubMed]
- Wyde, P.R.; Chetty, S.N.; Jewell, A.M.; Schoonover, S.L.; Piedra, P.A. Development of a cotton rat-human metapneumovirus (hMPV) model for identifying and evaluating potential hMPV antivirals and vaccines. Antiviral. Res. 2005, 66, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Yim, K.C.; Cragin, R.P.; Boukhvalova, M.S.; Blanco, J.C.; Hamlin, M.È.; Boivin, G.; Porter, D.D.; Prince, G.A. Human metapneumovirus: Enhanced pulmonary disease in cotton rats immunized with formalin-inactivated virus vaccine and challenged. Vaccine 2007, 25, 5034–5040. [Google Scholar] [CrossRef]
- Zhang, Y.; Niewiesk, S.; Li, J. Small Animal Models for Human Metapneumovirus: Cotton Rat is More Permissive than Hamster and Mouse. Pathogens 2014, 3, 633–655. [Google Scholar] [CrossRef]
- MacPhail, M.; Schickli, J.H.; Tang, R.S.; Kaur, J.; Robinson, C.; Fouchier, R.A.M.; Osterhaus, A.; Spaete, R.R.; Haller, A.A. Identification of small-animal and primate models for evaluation of vaccine candidates for human metapneumovirus (hMPV) and implications for hMPV vaccine design. J. Gen. Virol. 2004, 85 Pt 6, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Huang, J.; Rush, S.A.; Murray, J.; Gingerich, A.D.; Royer, F.; Hsieh, C.-L.; Tripp, R.A.; McLellan, J.S.; Mousa, J.J. Structural basis for ultrapotent antibody-mediated neutralization of human metapneumovirus. Proc. Natl. Acad. Sci. USA 2022, 119, e2203326119. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.V.; Wang, C.K.; Yang, C.-F.; Tollefson, S.J.; House, F.S.; Heck, J.M.; Chu, M.; Brown, J.B.; Lintao, L.D.; Quinto, J.D.; et al. The role of human metapneumovirus in upper respiratory tract infections in children: A 20-year experience. J. Infect. Dis. 2006, 193, 387–395. [Google Scholar] [CrossRef]
- Boukhvalova, M.; Blanco, J.C.; Falsey, A.R.; Mond, J. Treatment with novel RSV Ig RI-002 controls viral replication and reduces pulmonary damage in immunocompromised Sigmodon hispidus. Bone Marrow Transpl. 2016, 51, 119–126. [Google Scholar] [CrossRef]
- Blanco, J.C.G.; Richardson, J.Y.; Darnell, M.E.R.; Rowzee, A.; Pletneva, L.; Porter, D.D.; Prince, G.A. Cytokine and Chemokine Gene Expression after Primary and Secondary Respiratory Syncytial Virus Infection in Cotton Rats. J. Infect. Dis. 2002, 185, 1780–1785. [Google Scholar] [CrossRef]
- Blanco, J.C.G.; Pletneva, L.; Boukhvalova, M.; Richardson, J.Y.; Harris, K.A.; Prince, G.A. The cotton rat: An underutilized animal model for human infectious diseases can now be exploited using specific reagents to cytokines, chemokines, and interferons. J. Interferon. Cytokine Res. 2004, 24, 21–28. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Yim, K.C.; Prince, G.A.; Blanco, J.C. Methods for monitoring dynamics of pulmonary RSV replication by viral culture and by real-time reverse transcription-PCR in vivo: Detection of abortive viral replication. Curr. Protoc. Cell Biol. 2010, 46, 26.6.1–26.6.19. [Google Scholar] [CrossRef] [PubMed]
- Prince, G.A.; Curtis, S.J.; Yim, K.C.; Porter, D.D. Vaccine-enhanced respiratory syncytial virus disease in cotton rats following immunization with Lot 100 or a newly prepared reference vaccine. J. Gen. Virol. 2001, 82, 2881–2888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welliver, R.C. Review of epidemiology and clinical risk factors for severe respiratory syncytial virus (RSV) infection. J. Pediatr. 2003, 143 (Suppl. S5), S112–S117. [Google Scholar] [CrossRef] [PubMed]
- Lion, T. Adenovirus infections in immunocompetent and immunocompromised patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef]
- Hijano, D.R.; Maron, G.; Hayden, R.T. Respiratory Viral Infections in Patients With Cancer or Undergoing Hematopoietic Cell Transplant. Front. Microbiol. 2018, 9, 3097. [Google Scholar] [CrossRef]
- Pantaleo, G.; Correia, B.; Fenwick, C.; Joo, V.S.; Laurent Perez, L. Antibodies to combat viral infections: Development strategies and progress. Nat. Rev. Drug Discov. 2022, 21, 676–696. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, A.; Kuba, K.; Morita, M.; Chida, S.; Tezuka, H.; Hara, H.; Sasaki, T.; Ohteki, T.; Ranieri, V.M.; dos Santos, C.C.; et al. CXCL10-CXCR3 Enhances the Development of Neutrophil-mediated Fulminant Lung Injury of Viral and Nonviral Origin. Am. J. Respir. Crit. Care Med. 2013, 187, 65–77. [Google Scholar] [CrossRef]
- Shanley, T.P.; Schmal, H.; Friedl, H.P.; Jones, M.L.; Ward, P.A. Role of macrophage inflammatory protein-1 alpha (MIP-1 alpha) in acute lung injury in rats. J. Immunol. 1995, 154, 4793–4802. [Google Scholar] [CrossRef]
- Kameda, M.; Otsuka, M.; Chiba, H.; Kuronuma, K.; Hasegawa, T.; Takahashi, H. CXCL9, CXCL10, and CXCL11; biomarkers of pulmonary inflammation associated with autoimmunity in patients with collagen vascular diseases–associated interstitial lung disease and interstitial pneumonia with autoimmune features. PLoS ONE 2020, 15, e0241719. [Google Scholar] [CrossRef]
- Smith, R.E.; Strieter, R.M.; Phan, S.H.; Lukacs, N.W.; Huffnagle, G.B.; Wilke, C.A.; Burdick, M.D.; Lincoln, P.; Evanoff, H.; Kunkel, S.L. Production and function of murine macrophage inflammatory protein-1 alpha in bleomycin-induced lung injury. J. Immunol. 1994, 153, 4704–4712. [Google Scholar] [CrossRef]
- Johnson, R.A.; Prince, G.A.; Suffin, S.C.; Horswood, R.L.; Chanock, R.M. Respiratory syncytial virus infection in cyclophosphamide-treated cotton rats. Infect. Immun. 1982, 37, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Ottolini, M.G.; Porter, D.D.; Hemming, V.G.; Zimmerman, M.N.; Schwab, N.M.; Prince, G.A. Effectiveness of RSVIG prophylaxis and therapy of respiratory syncytial virus in an immunosuppressed animal model. Bone Marrow Transpl. 1999, 24, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, R.; Harrod, K.S.; Shieh, W.J.; Zaki, S.; Tripp, R.A. Human metapneumovirus persists in BALB/c mice despite the presence of neutralizing antibodies. J. Virol. 2004, 78, 14003–14011. [Google Scholar] [CrossRef]
- Alvarez, R.; Tripp, R.A. The immune response to human metapneumovirus is associated with aberrant immunity and impaired virus clearance in BALB/c mice. J. Virol. 2005, 79, 5971–5978. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, M.E.; Prince, G.A.; Gomez, A.M.; Kinkead, R.; Boivin, G. Human metapneumovirus infection induces long-term pulmonary inflammation associated with airway obstruction and hyperresponsiveness in mice. J. Infect. Dis. 2006, 193, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Moe, N.; Stenseng, I.H.; Krokstad, S.; Christensen, A.; Skanke, L.H.; Risnes, K.R.; Nordbø, S.A.; Døllner, H. The Burden of Human Metapneumovirus and Respiratory Syncytial Virus Infections in Hospitalized Norwegian Children. J. Infect. Dis. 2017, 216, 110–116. [Google Scholar] [CrossRef]
- Ebihara, T.; Endo, R.; Kikuta, H.; Ishiguro, N.; Ishiko, H.; Hara, M.; Takahashi, Y.; Kobayashi, K. Human Metapneumovirus Infection in Japanese Children. J. Clin. Microbiol. 2004, 42, 126–132. [Google Scholar] [CrossRef]
- Spahr, Y.; Tschudin-Sutter, S.; Baettig, V.; Compagno, F.; Tamm, M.; Halter, J.; Gerull, S.; Passweg, J.R.; Hirsch, H.H.; Khanna, N. Community-Acquired Respiratory Paramyxovirus Infection After Allogeneic Hematopoietic Cell Transplantation: A Single-Center Experience. Open Forum Infect. Dis. 2018, 5, ofy077. [Google Scholar] [CrossRef]
- Prince, G.A.; Mathews, A.; Curtis, S.J.; Porter, D.D. Treatment of respiratory syncytial virus bronchiolitis and pneumonia in a cotton rat model with systemically administered monoclonal antibody (palivizumab) and glucocorticosteroid. J. Infect. Dis. 2000, 182, 1326–1330. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Yim, K.C.; Kuhn, K.H.; Hemming, J.P.; Prince, G.A.; Porter, D.D.; Blanco, J.C. Age-related differences in pulmonary cytokine response to respiratory syncytial virus infection: Modulation by anti-inflammatory and antiviral treatment. J. Infect. Dis. 2007, 195, 511–518. [Google Scholar] [CrossRef]
- Rodriguez, W.J.; Gruber, W.C.; Groothuis, J.R.; Simoes, E.A.; Rosas, A.J.; Lepow, M.; Kramer, V.A. Hemming Respiratory syncytial virus immune globulin treatment of RSV lower respiratory tract infection in previously healthy children. Pediatrics 1997, 100, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, W.J.; Gruber, W.C.; Welliver, R.C.; Groothuis, J.R.; Simoes, E.A.F.; Meissner, H.C.; Hemming, V.G.; Hall, C.B.; Lepow, M.L.; Rosas, A.J. Respiratory syncytial virus (RSV) immune globulin intravenous therapy for RSV lower respiratory tract infection in infants and young children at high risk for severe RSV infections: Respiratory Syncytial Virus Immune Globulin Study Group. Pediatrics 1997, 99, 454–461. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yim, K.C.; Mousa, J.J.; Blanco, J.C.G.; Kim, S.; Boukhvalova, M.S. Human Metapneumovirus (hMPV) Infection and MPV467 Treatment in Immunocompromised Cotton Rats Sigmodon hispidus. Viruses 2023, 15, 476. https://doi.org/10.3390/v15020476
Yim KC, Mousa JJ, Blanco JCG, Kim S, Boukhvalova MS. Human Metapneumovirus (hMPV) Infection and MPV467 Treatment in Immunocompromised Cotton Rats Sigmodon hispidus. Viruses. 2023; 15(2):476. https://doi.org/10.3390/v15020476
Chicago/Turabian StyleYim, Kevin C., Jarrod J. Mousa, Jorge C. G. Blanco, Sonnie Kim, and Marina S. Boukhvalova. 2023. "Human Metapneumovirus (hMPV) Infection and MPV467 Treatment in Immunocompromised Cotton Rats Sigmodon hispidus" Viruses 15, no. 2: 476. https://doi.org/10.3390/v15020476