

Prevalence, Seroprevalence and Risk Factors of Avian Influenza in Wild Bird Populations in Korea: A Systematic Review and Meta-Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Systematic Review Protocol
2.2. Literature Search Strategy
2.3. Eligibility and Exclusion Criteria
2.4. Data Extraction
2.5. Risk of Bias Assessment
2.6. Data Synthesis
3. Results
3.1. Search Results
3.2. Study Characteristics
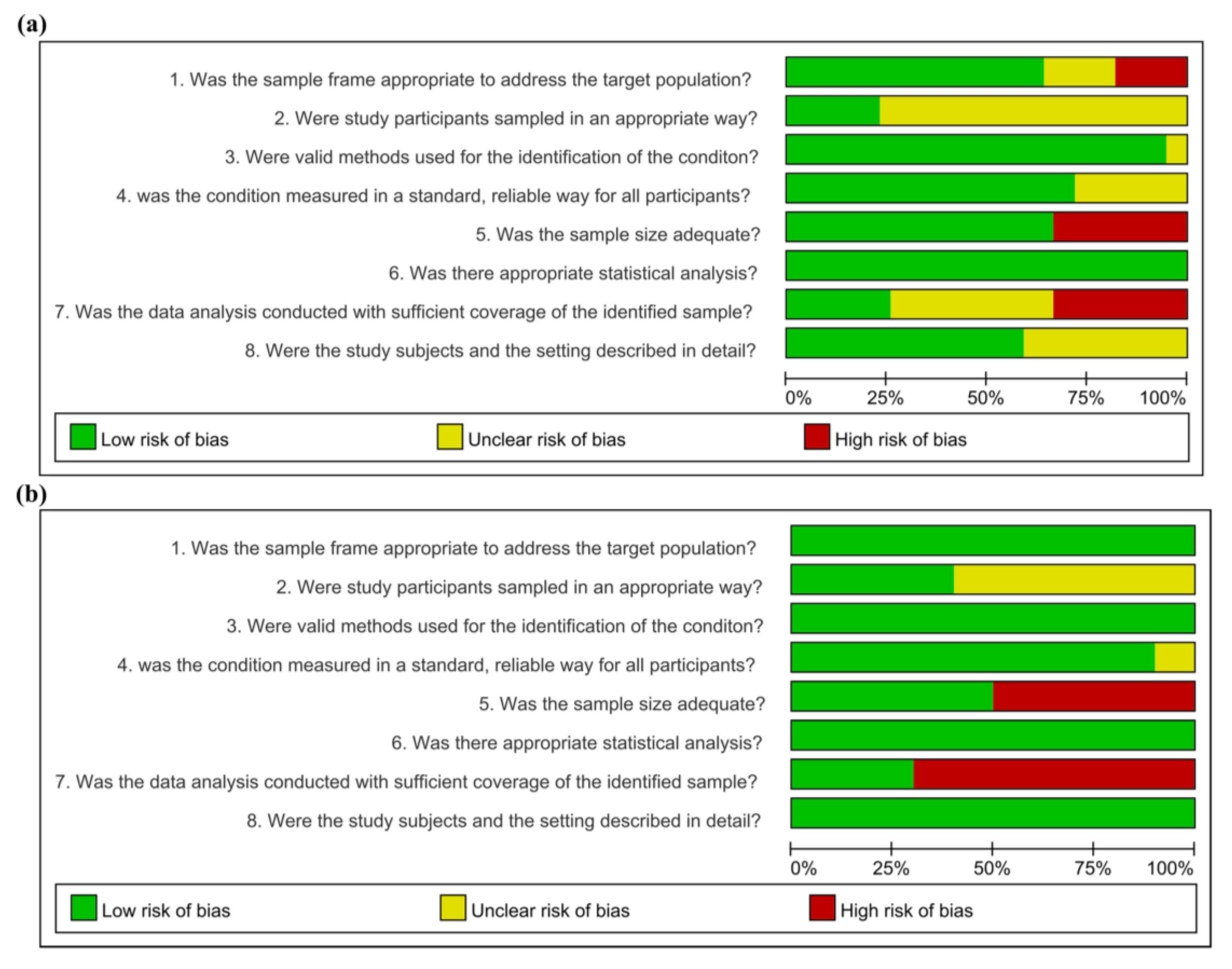
3.3. Risk of Bias Assessment
3.4. Meta-Analysis Results
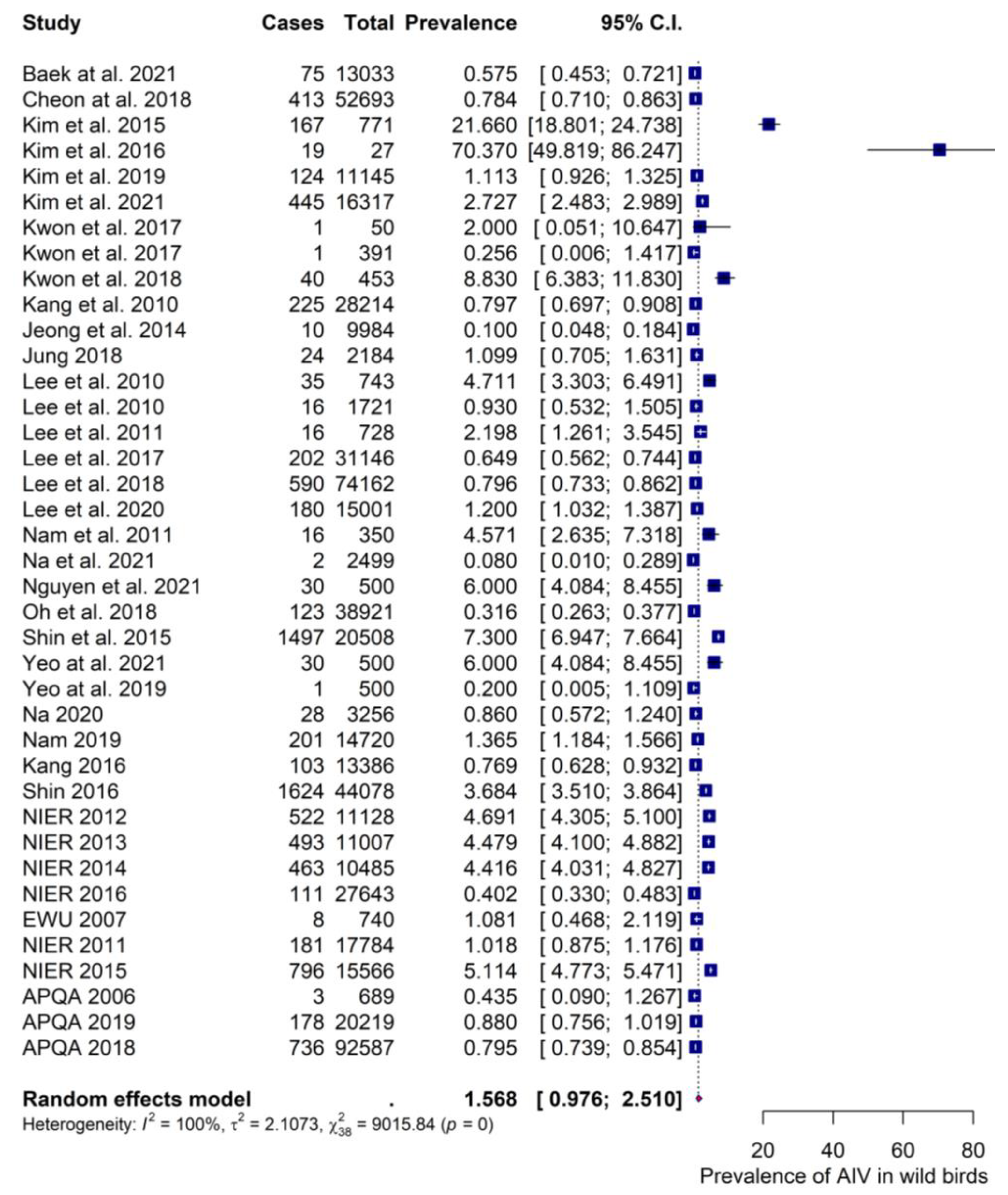
3.4.1. Prevalence Estimates
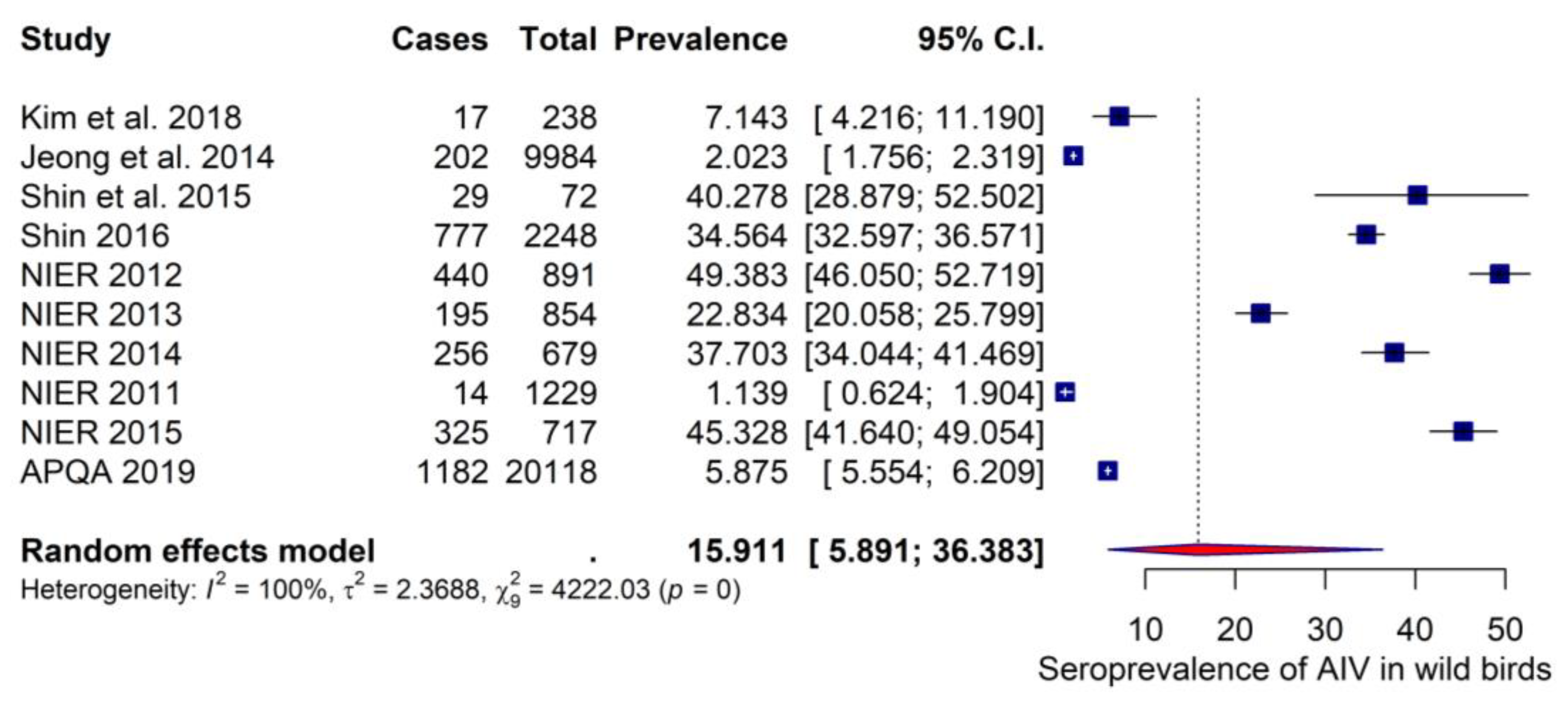
3.4.2. Seroprevalence Estimates
3.5. Publication Bias
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bodewes, R.; Kuiken, T. Changing role of wild birds in the epidemiology of avian influenza A viruses. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2018; pp. 279–307. [Google Scholar]
- Shin, J.H.; Woo, C.; Wang, S.J.; Jeong, J.; An, I.J.; Hwang, J.K.; Jo, S.D.; Yu, S.D.; Choi, K.; Chung, H.M.; et al. Prevalence of avian influenza virus in wild birds before and after the HPAI H5N8 outbreak in 2014 in South Korea. J. Microbiol. 2015, 53, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Vandegrift, K.J.; Sokolow, S.; Daszak, P.; Kilpatrick, A.M. Ecology of avian influenza viruses in a changing world. Ann. N. Y. Acad. Sci. 2010, 1195, 113–128. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Avian Influenza: Assessing the Pandemic Threat; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Stallknecht, D.E.; Brown, J.D. Wild bird infections and the ecology of avian influenza viruses. In Animal Influenza; Wiley Online Library: Hoboken, NJ, USA, 2017; pp. 153–176. [Google Scholar]
- MCEIRS. Avian Influenza: Detection in Wild Birds-Field Collection of Samples; MCEIRS: Minneapolis, MN, USA, 2017. [Google Scholar]
- Lee, Y.-N.; Cheon, S.-H.; Lee, E.-K.; Heo, G.-B.; Bae, Y.-C.; Joh, S.-J.; Lee, M.-H.; Lee, Y.-J. Pathogenesis and genetic characteristics of novel reassortant low-pathogenic avian influenza H7 viruses isolated from migratory birds in the Republic of Korea in the winter of 2016–2017. Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Jeong, D.G.; Yoon, S.W. Recent outbreaks of highly pathogenic avian influenza viruses in South Korea. Clin. Exp. Vaccine Res. 2017, 6, 95–103. [Google Scholar] [CrossRef]
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Bui, C.; Rahman, B.; Heywood, A.E.; MacIntyre, C.R. A Meta-Analysis of the Prevalence of Influenza A H5N1 and H7N9 Infection in Birds. Transbound Emerg. Dis. 2017, 64, 967–977. [Google Scholar] [CrossRef]
- Rabinowitz, P.; Perdue, M.; Mumford, E. Contact variables for exposure to avian influenza H5N1 virus at the human-animal interface. Zoonoses Public Health 2010, 57, 227–238. [Google Scholar] [CrossRef]
- Lu, L.; Lycett, S.J.; Leigh Brown, A.J. Reassortment patterns of avian influenza virus internal segments among different subtypes. BMC Evol. Biol. 2014, 14, 16. [Google Scholar] [CrossRef]
- To, K.K.; Chan, J.F.; Chen, H.; Li, L.; Yuen, K.-Y. The emergence of influenza A H7N9 in human beings 16 years after influenza A H5N1: A tale of two cities. Lancet Infect. Dis. 2013, 13, 809–821. [Google Scholar] [CrossRef]
- Mo, I.-P.; Bae, Y.-J.; Lee, S.-B.; Mo, J.-S.; Oh, K.-H.; Shin, J.-H.; Kang, H.-M.; Lee, Y.-J. Review of Avian Influenza Outbreaks in South Korea from 1996 to 2014. Avian Dis. 2016, 60 (Suppl. S1), 172–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; Choi, Y.-K.; Kim, Y.-J.; Song, M.-S.; Jeong, O.-M.; Lee, E.-K.; Jeon, W.-J.; Jeong, W.; Joh, S.-J.; Choi, K.-S.; et al. Highly pathogenic avian influenza virus (H5N1) in domestic poultry and relationship with migratory birds, South Korea. Emerg. Infect. Dis. 2008, 14, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Jeong, O.M.; Kim, M.C.; Kwon, J.S.; Paek, M.R.; Choi, J.G.; Lee, E.K.; Kim, Y.J.; Lee, Y.J. Surveillance of avian influenza virus in wild bird fecal samples from South Korea, 2003–2008. J. Wildl. Dis. 2010, 46, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.M.; Bousfield, R.B.; Bissett, L.A.; Dyrting, K.C.; Luk, G.S.M.; Tsim, S.T.; Sturm-Ramirez, K.; Webster, R.G.; Guan, Y.; Peiris, J.S.M. Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. Avian Pathol. 2004, 33, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, Y.; Li, Z.; Shi, J.; Shinya, K.; Deng, G.; Qi, Q.; Tian, G.; Fan, S.; Zhao, H.; et al. Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in western China. J. Virol. 2006, 80, 5976–5983. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Smith, G.J.; Zhang, S.Y.; Qin, K.; Wang, J.; Li, K.S.; Webster, R.G.; Peiris, J.S.; Guan, Y. Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature 2005, 436, 191–192. [Google Scholar] [CrossRef]
- Paudyal, N.; Pan, H.; Liao, X.; Zhang, X.; Li, X.; Fang, W.; Yue, M. A Meta-Analysis of Major Foodborne Pathogens in Chinese Food Commodities Between 2006 and 2016. Foodborne Pathog. Dis. 2018, 15, 187–197. [Google Scholar] [CrossRef]
- Field, A.P.; Gillett, R. How to do a meta-analysis. Br. J. Math Stat. Psychol. 2010, 63, 665–694. [Google Scholar] [CrossRef]
- Haidich, A.B. Meta-analysis in medical research. Hippokratia 2010, 14, 29–37. [Google Scholar]
- Barendregt, J.J.; Doi, S.A.; Lee, Y.Y.; Norman, R.E.; Vos, T. Meta-analysis of prevalence. J. Epidemiol. Community Health 2013, 67, 974–978. [Google Scholar] [CrossRef]
- Islam, M.Z.; Musekiwa, A.; Islam, K.; Ahmed, S.; Chowdhury, S.; Ahad, A.; Biswas, P.K. Regional variation in the prevalence of E. coli O157 in cattle: A meta-analysis and meta-regression. PLoS ONE 2014, 9, e93299. [Google Scholar] [CrossRef]
- Ferreira, G.C.M.; Canozzi, M.E.A.; Peripolli, V.; Moura, G.d.P.; Sánchez, J.; Martins, C.E.N. Prevalence of bovine Babesia spp., Anaplasma marginale, and their co-infections in Latin America: Systematic review-meta-analysis. Ticks Tick Borne Dis. 2022, 13, 101967. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Munn, Z.; Moola, S.; Lisy, K.; Riitano, D.; Tufanaru, C. Methodological guidance for systematic reviews of observational epidemiological studies reporting prevalence and cumulative incidence data. Int. J. Evid. Based Health C 2015, 13, 147–153. [Google Scholar] [CrossRef]
- Porritt, K.; Gomersall, J.; Lockwood, C. JBI’s Systematic Reviews: Study selection and critical appraisal. Am. J. Nurs. 2014, 114, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 24 December 2021).
- Balduzzi, S.; Rucker, G.; Schwarzer, G. How to perform a meta-analysis with R: A practical tutorial. Evid. Based Ment. Health 2019, 22, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Alboukadel, K. ggcorrplot: Visualization of a Correlation Matrix using‘ggplot2’. CRAN 2019. [Google Scholar]
- Harrer, M.; Cuijpers, P.; Furukawa, T.A.; Ebert, D.D. Doing Meta-Analysis with R: A Hands-On Guide; Chapman and Hall/CRC: London, UK, 2021. [Google Scholar]
- Viechtbauer, W. Conducting Meta-Analyses in R with the metafor Package. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef]
- Golden, C.E.; Mishra, A. Prevalence of Salmonella and Campylobacter spp. in Alternative and Conventionally Produced Chicken in the United States: A Systematic Review and Meta-Analysis. J. Food Prot. 2020, 83, 1181–1197. [Google Scholar] [CrossRef]
- Lin, L.F.; Chu, H.T. Meta-analysis of Proportions Using Generalized Linear Mixed Models. Epidemiology 2020, 31, 713–717. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Sabarimurugan, S.; Kumarasamy, C.; Baxi, S.; Devi, A.; Jayaraj, R. Systematic review and meta-analysis of prognostic microRNA biomarkers for survival outcome in nasopharyngeal carcinoma. PLoS ONE 2019, 14, e0209760. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Duval, S.; Tweedie, R. Trim and fill: A simple funnel-plot-based method of testing and adjusting for publication bias in meta-analysis. Biometrics 2000, 56, 455–463. [Google Scholar] [CrossRef]
- National Institute of Environmental Research. Surveillance and Mornitoring of WiIdlife Diseases in Korea—Avian Influenza Surveillance in Wild Animals; National Institute of Environmental Research: Incheon, Republic of Korea, 2012.
- National Institute of Environmental Research. Avian Influenza Surveillance in Wild Animals; National Institute of Environmental Research: Incheon, Republic of Korea, 2013.
- National Institute of Environmental Research. Avian Influenza Surveillance and Molecular-Biological Characterization of the Spatiotemporal Distribution of Avian Influenza Viruses in 2014; National Institute of Environmental Research: Incheon, Republic of Korea, 2014.
- National Institute of Environmental Research. Research on Early Detection and Characterization of Zoonotic Diseases in Wild Birds (I); National Institute of Environmental Research: Incheon, Republic of Korea, 2016.
- Animal and Plant Quarantine Agency. A Study on the Epidemiology of Highly Pathogenic Avian Influenza; Animal and Plant Quarantine Agency: Gimcheon, Republic of Korea, 2018. [Google Scholar]
- Animal and Plant Quarantine Agency. A Study on the Epidemiology of Highly Pathogenic Avian Influenza; Animal and Plant Quarantine Agency: Gimcheon, Republic of Korea, 2019. [Google Scholar]
- Cheon, S.-H.; Lee, Y.-N.; Kang, S.-I.; Kye, S.-J.; Lee, E.-K.; Heo, G.-B.; Lee, M.-H.; Kim, J.-W.; Lee, K.-N.; Son, H.-M.; et al. Genetic evidence for the intercontinental movement of avian influenza viruses possessing North American-origin nonstructural gene allele B into South Korea. Infect. Genet. Evol. 2018, 66, 18–25. [Google Scholar] [CrossRef]
- National Institute of Environmental Research. Monitoring the Infection and Transmission Status of Avian Influenza in the Migratory and Resident Wild Birds. In Monitoring the Infection and Transmission Status of Avian Influenza in the Migratory and Resident Wild Birds; National Institute of Environmental Research: Incheon, Republic of Korea, 2011. [Google Scholar]
- Jeong, J.; Kang, H.-M.; Lee, E.-K.; Song, B.-M.; Kwon, Y.-K.; Kim, H.-R.; Choi, K.-S.; Kim, J.-Y.; Lee, H.-J.; Moon, O.-K.; et al. Highly pathogenic avian influenza virus (H5N8) in domestic poultry and its relationship with migratory birds in South Korea during 2014. Vet. Microbiol. 2014, 173, 249–257. [Google Scholar] [CrossRef]
- National Institute of Environmental Research. Surveillance on Avian Influenza of Wild Birds and Molecular-Biological Characterization of the Spatiotemporal Distribution of Avian Influenza Viruses in 2015; National Institute of Environmental Research: Incheon, Republic of Korea, 2015.
- Shin, J.-H. Spatiotemporal Distribution of Avian Influenza Viruses in Wild Birds and Tracking Their Migration Route; Chungbuk National University: Chungju, Republic of Korea, 2016. [Google Scholar]
- Lee, E.-K.; Kang, H.-M.; Song, B.-M.; Lee, Y.-N.; Heo, G.-B.; Lee, H.-S.; Lee, Y.-J.; Kim, J.-H. Surveillance of avian influenza viruses in South Korea between 2012 and 2014. Virol. J. 2017, 14, 54. [Google Scholar] [CrossRef]
- Kim, G.-S.; Kim, T.-S.; Son, J.-S.; Lai, V.D.; Park, J.-E.; Wang, S.-J.; Jheong, W.-H.; Mo, I.-P. The difference of detection rate of avian influenza virus in the wild bird surveillance using various methods. J. Vet. Sci. 2019, 20, 1134050. [Google Scholar] [CrossRef]
- Nam, J.-H. Surveillance and Genetic Characterization of Avian and Canine Influenza Viruses Isolated from South Korea; Korea University: Seoul, Republic of Korea, 2019. [Google Scholar]
- Lee, Y.-N.; Lee, D.-H.; Cheon, S.-H.; Park, Y.-R.; Baek, Y.-G.; Si, Y.-J.; Kye, S.-J.; Lee, E.-K.; Heo, G.-B.; Bae, Y.-C. Genetic characteristics and pathogenesis of H5 low pathogenic avian influenza viruses from wild birds and domestic ducks in South Korea. Sci. Rep. 2020, 10, 12151. [Google Scholar] [CrossRef]
- Yeo, S.-J.; Hoang, V.T.; Duong, T.B.; Nguyen, N.M.; Tuong, H.T.; Azam, M.; Sung, H.W.; Park, H. Emergence of a Novel Reassortant H5N3 Avian Influenza Virus in Korean Mallard Ducks in 2018. Intervirology 2021, 65, 1–16. [Google Scholar] [CrossRef]
- Animal and Plant Quarantine Agency. Investigation and Epidemiological Analysis of Avian Influenza Infection in Ornamental Birds and Birds Inhabiting Dadohaehaesang National Park; Animal and Plant Quarantine Agency: Gimcheon, Republic of Korea, 2006. [Google Scholar]
- Studies on Findings of Mechanism and Separation of Avian Influenza Virus; Ministry of Agriculture and Forestry: Gwacheon, Republic of Korea, 2007.
- Lee, D.-H.; Lee, H.-J.; Kang, H.-M.; Jeong, O.-M.; Kim, M.-C.; Kwon, J.-S.; Kwon, J.-H.; Kim, C.-B.; Lee, J.-B.; Park, S.-Y.; et al. DNA Barcoding Techniques for Avian Influenza Virus Surveillance in Migratory Bird Habitats. J. Wildl. Dis. 2010, 46, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Kang, M. Characterization of Influenza A Virus Derived from Animals and Risk Assessment for Interspecies Transmissibility; Jeonbuk National Uuniversity: Iksan, Republic of Korea, 2016. [Google Scholar]
- Jung, S.-H. Surveillance and Genetic Characterization of Avian Influenza Viruses in South Korea During the Winter of 2016–2017; Korea University: Seoul, Republic of Korea, 2018. [Google Scholar]
- Kim, E.-H.; Kim, Y.-L.; Kim, S.M.; Yu, K.-M.; Casel, M.A.B.; Jang, S.-G.; Pascua, P.N.Q.; Webby, R.J.; Choi, Y.K. Pathogenic assessment of avian influenza viruses in migratory birds. Emerg. Microbes Infect. 2021, 10, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Na, E.-J.; Kim, Y.-S.; Kim, Y.-J.; Park, J.-S.; Oem, J.-K. Genetic Characterization and Pathogenicity of H7N7 and H7N9 Avian Influenza Viruses Isolated from South Korea. Viruses 2021, 13, 2057. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Park, J.-K.; Youn, H.-N.; Lee, Y.-N.; Lim, T.-H.; Kim, M.-S.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; Song, C.-S. Surveillance and Isolation of HPAI H5N1 from Wild Mandarin Ducks (Aix galericulata). J. Wildl. Dis. 2011, 47, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.-H.; Kim, E.-H.; Song, D.; Choi, Y.K.; Kim, J.-K.; Poo, H. Emergence of Mammalian Species-Infectious and -Pathogenic Avian Influenza H6N5 Virus with No Evidence of Adaptation. J. Virol. 2011, 85, 13271–13277. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Lee, D.-H.; Swayne, D.; Noh, J.-Y.; Yuk, S.-S.; Erdene-Ochir, T.-O.; Hong, W.-T.; Jeong, J.-H.; Jeong, S.; Gwon, G.-B.; et al. Reassortant Clade 2.3.4.4 Avian Influenza A(H5N6) Virus in a Wild Mandarin Duck, South Korea. Emerg. Infect. Dis. 2017, 23, 5. [Google Scholar]
- Kwon, J.-H.; Jeong, S.; Lee, D.-H.; Swayne, D.E.; Kim, Y.; Lee, S.; Noh, J.-Y.; Erdene-Ochir, T.-O.; Jeong, J.-H.; Song, C.-S. New Reassortant Clade 2.3.4.4b Avian Influenza A(H5N6) Virus in Wild Birds, South Korea, 2017–2018. Infect. Dis. J. CDC 2018, 24, 10. [Google Scholar]
- Kim, H.-R.; Kwon, Y.-K.; Jang, I.; Lee, Y.-J.; Kang, H.-M.; Lee, E.-K.; Song, B.-M.; Lee, H.-S.; Joo, Y.-S.; Lee, K.-H.; et al. Pathologic Changes in Wild Birds Infected with Highly Pathogenic Avian Influenza A(H5N8) Viruses, South Korea, 2014. Emerg. Infect. Dis. 2015, 21, 5. [Google Scholar] [CrossRef]
- Kim, S.-H.; Hur, M.; Suh, J.-H.; Woo, C.; Wang, S.-J.; Park, E.-R.; Hwang, J.; An, I.-J.; Jo, S.-D.; Shin, J.-H.; et al. Molecular characterization of highly pathogenic avian influenza H5N8 viruses isolated from Baikal teals found dead during a 2014 outbreak in Korea. J. Vet. Sci. 2016, 17, 299–306. [Google Scholar] [CrossRef]
- Baek, Y.-G.; Lee, Y.-N.; Lee, D.-H.; Shin, J.-I.; Lee, J.-H.; Chung, D.; Lee, E.-K.; Heo, G.-B.; Sagong, M.; Kye, S.-J.; et al. Multiple Reassortants of H5N8 Clade 2.3.4.4b Highly Pathogenic Avian Influenza Viruses Detected in South Korea during the Winter of 2020–2021. Viruses 2021, 13, 490. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Lee, D.-H.; Jeong, J.-H.; Yuk, S.-S.; Erdene-Ochir, T.-O.; Noh, J.-Y.; Hong, W.-T.; Jeong, S.; Gwon, G.-B.; Lee, S.-W.; et al. Isolation of an H5N8 Highly Pathogenic Avian Influenza Virus Strain from Wild Birds in Seoul, a Highly Urbanized Area in South Korea. J. Wildl. Dis. 2017, 53, 630–635. [Google Scholar] [CrossRef]
- Oh, K.-H.; Mo, J.-S.; Bae, Y.-J.; Lee, S.-B.; Lai, V.D.; Wang, S.-J.; Mo, I.-P. Amino acid substitutions in low pathogenic avian influenza virus strains isolated from wild birds in Korea. Virus Genes 2018, 54, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.-J.; Than, D.-D.; Park, H.-S.; Sung, H.W.; Park, H. Molecular Characterization of a Novel Avian Influenza A (H2N9) Strain Isolated from Wild Duck in Korea in 2018. Viruses 2019, 11, 1046. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.T.V.; Hoang, V.T.; Sung, H.W.; Yeo, S.-J.; Park, H. Genetic Characterization and Pathogenesis of Three Novel Reassortant H5N2 Viruses in South Korea, 2018. Viruses 2021, 13, 2192. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Lee, H.-J.; Lee, Y.-N.; Jeong, O.-M.; Kang, H.-M.; Kim, M.-C.; Kwon, J.-S.; Kwon, J.-H.; Lee, J.-B.; Park, S.-Y.; et al. Application of DNA Barcoding Technique in Avian Influenza Virus Surveillance of Wild Bird Habitats in Korea and Mongolia. Avian Dis. 2010, 54, 677–681. [Google Scholar] [CrossRef]
- Na, E.-J. Genotyping and Genetic Characteristics of Avian Influenza Virus isolated from Wild Birds in South Korea, 2019–2020; Jeonbuk National University: Iksan, Republic of Korea, 2020. [Google Scholar]
- Animal and Plant Quarantine Agency. Survey of HPAI Infection Status in Wild Birds; Animal and Plant Quarantine Agency: Gimcheon, Republic of Korea, 2018. [Google Scholar]
- Kim, H.K.; Kim, H.J.; Noh, J.Y.; Van Phan, L.; Kim, J.H.; Song, D.; Na, W.; Kang, A.; Nguyen, T.L.; Shin, J.H. Serological evidence of H5-subtype influenza A virus infection in indigenous avian and mammalian species in Korea. Arch. Virol. 2018, 163, 649–657. [Google Scholar] [CrossRef]
- Takekawa, J.Y.; Heath, S.R.; Douglas, D.C.; Perry, W.M.; Javed, S.; Newman, S.; Suwal, R.N.; Rahmani, A.R.; Choudhury, B.C.; Prosser, D.J. Geographic variation in Bar-headed geese Anser indicus: Connectivity of wintering and breeding grounds across a broad front. Wildfowl 2009, 59, 100–123. [Google Scholar]
- NIBR (National Institute of Biological Resources). Winter Waterbird Census of Korea; National Institute of Biological Resources: Incheon, Republic of Korea, 2020. [Google Scholar]
- Chen, X.; Li, C.; Sun, H.-T.; Ma, J.; Qi, Y.; Qin, S.-Y. Prevalence of avian influenza viruses and their associated antibodies in wild birds in China: A systematic review and meta-analysis. Microb. Pathog. 2019, 135, 103613. [Google Scholar] [CrossRef]
- Chen, X.; Qi, Y.; Wang, H.; Wang, Y.; Wang, H.; Ni, H. Prevalence of Multiple Subtypes of Avian Influenza Virus Antibodies in Egg Yolks of Mallards (Anas platyrhynchos) and White-winged Terns (Chlidonias leucopterus) in the Northeastern Republic of China. J. Wildl. Dis. 2018, 54, 834–837. [Google Scholar] [CrossRef]
- Muzaffar, S.B.; Ydenberg, R.C.; Jones, I.L. Avian influenza: An ecological and evolutionary perspective for waterbird scientists. Waterbirds 2006, 29, 243–257. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Kang, H.-M.; Lee, E.K.; Song, B.M.; Jeong, J.; Kwon, Y.K.; Kim, H.R.; Lee, K.J.; Hong, M.S.; Jang, I. Novel reassortant influenza A (H5N8) viruses, South Korea, 2014. Emerg. Infect. Dis. 2014, 20, 1087. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, A.; Rezaei, F.; Nasab, G.S.F.; Adjaminezhad-Fard, F.; Noroozbabaei, Z.; Mokhtari-Azad, T. The Comparison of Sensitivity and Specificity of ELISA-based Microneutralization Test with Hemagglutination Inhibition Test to Evaluate Neutralizing Antibody against Influenza Virus (H1N1). Iran J. Public Health 2017, 46, 1690–1696. [Google Scholar] [PubMed]
- Rowe, T.; Abernathy, R.A.; Hu-Primmer, J.; Thompson, W.W.; Lu, X.; Lim, W.; Fukuda, K.; Cox, N.J.; Katz, J.M. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. J. Clin. Microbiol. 1999, 37, 937–943. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Prevalence Estimates (95%) | I2 (%) | τ2 | Psubgroup |
|---|---|---|---|---|
| 1. Bird species | <0.01 a | |||
| Anseriformes | 4.34 [1.44; 12.30] | 98 | 3.2233 | |
| Charadriiformes | 0.19 [0.03; 1.33] | 0 | 0 | |
| Other species | 0.22 [0.04; 1.20] | 0 | 1.9568 | |
| Not reported | 1.20 [0.74; 1.94] | 100 | 2.1236 | |
| Overall prevalence | 1.14 [0.72; 1.82] | 99 | 2.8912 | |
| 2. Sample type | <0.01 a | |||
| Feces | 1.58 [1.06; 2.36] | 99 | 1.2236 | |
| Carcass | 4.59 [0.76; 23.07] | 95 | 7.1496 | |
| Cloacal swabs | 1.00 [0.36; 2.75] | 100 | 2.6837 | |
| Mixed samples | 0.63 [0.56; 0.71] | 0 | 0 | |
| Not reported | 0.97 [0.73; 1.28] | 96 | 0.039 | |
| Overall prevalence | 1.7 [1.10; 2.64] | 100 | 2.6417 | |
| 3. Detection method | =0.25 b | |||
| RT-PCR | 1.98 [1.11; 3.53] | 100 | 2.3799 | |
| HA-test | 1.06 [0.59; 1.90] | 95 | 0.5665 | |
| rRT-PCR | 0.75 [0.32;1.75] | 85 | 0.8644 | |
| Virus isolation | 1.25 [0.14; 10.25] | 88 | 2.0493 | |
| Overall prevalence | 1.54 [0.99; 2.38] | 100 | 2.0266 | |
| 4. Sample size | =0.05 b | |||
| Less than 1000 | 3.28 [1.20; 8.68] | 97 | 3.3332 | |
| More than 1000 | 1.10 [0.74; 1.65] | 100 | 1.1152 | |
| Overall prevalence | 1.57 [0.99; 2.47] | 100 | 2.1073 | |
| 5. Sampling season | =0.48 b | |||
| Fall to winter | 1.94 [0.80; 4.61] | 98 | 3.1168 | |
| Not reported | 1.35 [0.82; 2.20] | 100 | 1.4628 | |
| Overall prevalence | 1.57 [0.99; 2.47] | 100 | 2.1073 | |
| 6. Publication status | =0.82 b | |||
| Published | 1.63 [0.80; 3.27] | 100 | 3.07 | |
| Non-published | 1.47 [0.95; 2.28] | 100 | 0.7476 | |
| Overall prevalence | 1.57 [0.99; 2.47] | 100 | 2.1073 |
| Variables | Seroprevalence Estimates (95%) | I2 (%) | τ2 | Psubgroup |
|---|---|---|---|---|
| 1. Bird species | <0.01 a | |||
| Anseriformes | 30.45 [18.97; 45.03] | 100 | 0.7793 | |
| Charadriiformes | 2.95 [0.24; 27.43] | 49 | 5.2003 | |
| Other species | 2.83 [0.40; 17.26] | 94 | 7.0157 | |
| Not reported | 2.85 [1.17; 6.76] | 99 | 0.6122 | |
| Overall prevalence | 7.71 [3.33; 16.86] | 99 | 4.3612 | |
| 2. Detection method | <0.01 a | |||
| ELISA | 31.47 [20.47; 45.02] | 97 | 0.5904 | |
| HI test | 2.46 [1.12; 5.31] | 99 | 0.475 | |
| Overall prevalence | 15.90 [6.76; 33.01] | 100 | 2.3668 | |
| 3. Sample size | <0.01 a | |||
| Less than 1000 | 30.93 [18.48; 46.93] | 98 | 0.7001 | |
| More than 1000 | 5.03 [1.25; 18.20] | 100 | 2.1246 | |
| Overall prevalence | 15.90 [6.76; 33.01] | 100 | 2.3668 | |
| 4. Sampling season | =0.02 a | |||
| Fall to winter | 36.48 [24.05; 51.01] | 98 | 0.2553 | |
| Not reported | 10.48 [3.46; 27.66] | 100 | 2.53 | |
| Overall prevalence | 15.90 [6.76; 33.01] | 100 | 2.3668 | |
| 5. Publication status | =0.37 b | |||
| Published | 9.07 [1.91; 33.78] | 99 | 2.0361 | |
| Non-published | 19.85 [7.52; 43.01] | 100 | 2.2482 | |
| Overall prevalence | 15.9 [ 6.76; 33.01] | 100 | 2.2482 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntakiyisumba, E.; Lee, S.; Park, B.-Y.; Tae, H.-J.; Won, G. Prevalence, Seroprevalence and Risk Factors of Avian Influenza in Wild Bird Populations in Korea: A Systematic Review and Meta-Analysis. Viruses 2023, 15, 472. https://doi.org/10.3390/v15020472
Ntakiyisumba E, Lee S, Park B-Y, Tae H-J, Won G. Prevalence, Seroprevalence and Risk Factors of Avian Influenza in Wild Bird Populations in Korea: A Systematic Review and Meta-Analysis. Viruses. 2023; 15(2):472. https://doi.org/10.3390/v15020472
Chicago/Turabian StyleNtakiyisumba, Eurade, Simin Lee, Byung-Yong Park, Hyun-Jin Tae, and Gayeon Won. 2023. "Prevalence, Seroprevalence and Risk Factors of Avian Influenza in Wild Bird Populations in Korea: A Systematic Review and Meta-Analysis" Viruses 15, no. 2: 472. https://doi.org/10.3390/v15020472