
Monitoring of Insecticide Resistance Mutations and Pathogen Circulation in Sand Flies from Emilia-Romagna, a Leishmaniasis Endemic Region of Northern Italy

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
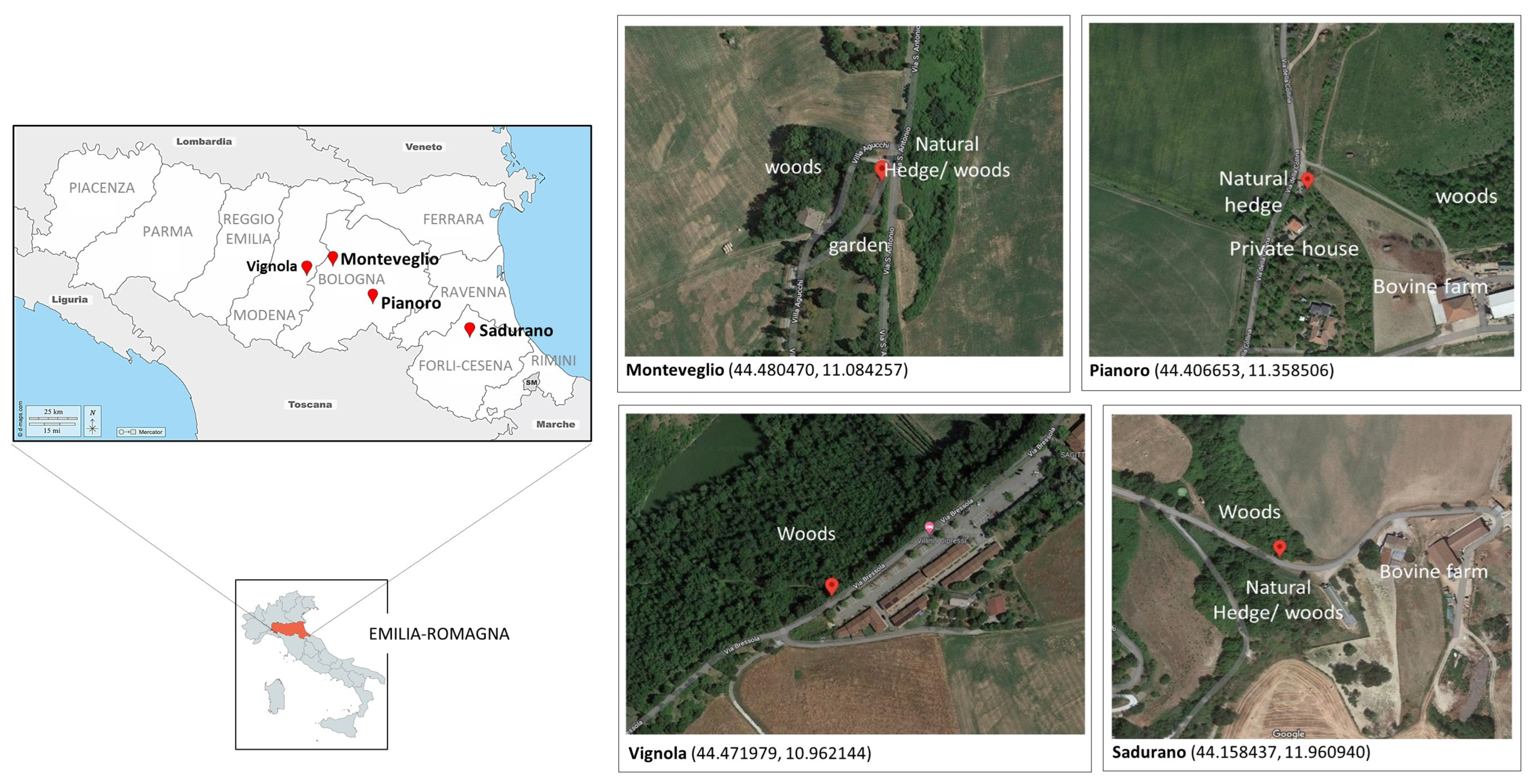
2.1. Sampling Areas, Sand Fly Collection, and Sample Handling

2.2. Genetic Material Extraction from Sand Flies
2.3. Molecular Identification of Species and Genotyping of Mutations in the Voltage-Gated Sodium Channel (VGSC)
2.4. Pathogen Detection
3. Results
3.1. Sand Fly Collections and Species Identification
3.2. Monitoring of Knock-Down Resistance (kdr) Mutations
3.3. Detection of Leishmania and Phleboviruses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Available online: https://www.euro.who.int/__data/assets/pdf_file/0007/246166/Fact-sheet-Leishmaniasis-Eng.pdf (accessed on 2 November 2022).
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M. Who Leishmaniasis Control the WHO Leishmaniasis Control Team Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Semenza, J.C. Vector-borne disease emergence and spread in the European Union. In Global Health Impacts of Vector-Borne Diseases: Workshop Summary. Forum on Microbial Threats; Board on Global Health; Health and Medicine Division; National Academies of Sciences, Engineering, and Medicine; National Academies Press: Washington, DC, USA, 2016; p. A11. Available online: https://www.ncbi.nlm.nih.gov/books/NBK390443/ (accessed on 28 October 2022).
- Maroli, M.; Rossi, L.; Baldelli, R.; Capelli, G.; Ferroglio, E.; Genchi, C.; Gramiccia, M.; Mortarino, M.; Pietrobelli, M.; Gradoni, L. The northward spread of leishmaniasis in Italy: Evidence from retrospective and ongoing studies on the canine reservoir and phlebotomine vectors. Trop. Med. Int. Health 2008, 13, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Roldan, J.; Benelli, G.; Panarese, R.; Iatta, R.; Furlanello, T.; Beugnet, F.; Zatelli, A.; Otranto, D. Leishmania infantum and Dirofilaria immitis infections in Italy, 2009–2019: Changing distribution patterns. Parasites Vectors 2020, 13, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Kasap, O.E.; Alten, B.; De Lamballerie, X.; Charrel, R.N. Sandfly-Borne Phlebovirus Isolations from Turkey: New Insight into the Sandfly fever Sicilian and Sandfly fever Naples Species. PLoS Negl. Trop. Dis. 2016, 10, e0004519. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Chiapponi, C.; Bellini, R.; Bonilauri, P.; Lelli, D.; Moreno, A.; Barbieri, I.; Pongolini, S.; Lavazza, A.; Dottori, M. Isolation of three novel reassortant phleboviruses, Ponticelli I, II, III, and of Toscana virus from field-collected sand flies in Italy. Parasites Vectors 2018, 11, 84. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Ferrarini, G.; Bonilauri, P.; Lelli, D.; Chiapponi, C.; Bellini, R.; Dottori, M. Co-circulation of eight different phleboviruses in sand flies collected in the Northern Apennine Mountains (Italy). Infect. Genet. Evol. 2018, 64, 131–134. [Google Scholar] [CrossRef]
- Calzolari, M.; Romeo, G.; Callegari, E.; Bonilauri, P.; Chiapponi, C.; Carra, E.; Rugna, G.; Taddei, R.; Lelli, D.; Dottori, M. Co-Circulation of Phleboviruses and Leishmania Parasites in Sand Flies from a Single Site in Italy Monitored between 2017 and 2020. Viruses 2021, 13, 1660. [Google Scholar] [CrossRef]
- Moriconi, M.; Rugna, G.; Calzolari, M.; Bellini, R.; Albieri, A.; Angelini, P.; Cagarelli, R.; Landini, M.P.; Charrel, R.N.; Varani, S. Phlebotomine sand fly–borne pathogens in the Mediterranean Basin: Human leishmaniasis and phlebovirus infections. PLoS Negl. Trop. Dis. 2017, 11, e0005660. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Leishmaniasis. 2020. Available online: https://www.who.int/health-topics/leishmaniasis#tab=tab_1 (accessed on 28 September 2022).
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef]
- Malagoli, C.; Costanzini, S.; Heck, J.E.; Malavolti, M.; De Girolamo, G.; Oleari, P.; Palazzi, G.; Teggi, S.; Vinceti, M. Passive exposure to agricultural pesticides and risk of childhood leukemia in an Italian community. Int. J. Hyg. Environ. Health 2016, 219, 742–748. [Google Scholar] [CrossRef] [Green Version]
- EU. Regulation (EU) No 528/2012 of the European Parliament and of the Council of 22 May 2012 concerning the making availa-ble on the market and use of biocidal products. Off. J. Eur. Union 2012, L167, 1–123. Available online: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2012:167:0001:0123:EN:PDF (accessed on 28 October 2022).
- Pichler, V.; Giammarioli, C.; Bellini, R.; Veronesi, R.; Arnoldi, D.; Rizzoli, A.; Lia, R.P.; Otranto, D.; Ballardini, M.; Cobre, P.; et al. First evidence of pyrethroid resistance in Italian populations of West Nile virus vector Culex pipiens. Med. Veter. Ѐntomol. 2022, 36, 390–395. [Google Scholar] [CrossRef]
- Canali, M.; Rivas-Morales, S.; Beutels, P.; Venturelli, C. The Cost of Arbovirus Disease Prevention in Europe: Area-Wide Integrated Control of Tiger Mosquito, Aedes albopictus, in Emilia-Romagna, Northern Italy. Int. J. Environ. Res. Public Health 2017, 14, 444. [Google Scholar] [CrossRef] [Green Version]
- Porretta, D.; Mastrantonio, V.; Lucchesi, V.; Bellini, R.; Vontas, J.; Urbanelli, S. Historical samples reveal a combined role of agriculture and public-health applications in vector resistance to insecticides. Pest Manag. Sci. 2022, 78, 1567–1572. [Google Scholar] [CrossRef]
- Balaska, S.; Fotakis, E.A.; Chaskopoulou, A.; Vontas, J. Chemical control and insecticide resistance status of sand fly vectors worldwide. PLoS Negl. Trop. Dis. 2021, 15, e0009586. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Tarallo, V.D.; Otranto, D. Morphological keys for the identification of Italian phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae). Parasites Vectors 2014, 7, 479. [Google Scholar] [CrossRef]
- d-maps.com. Free Maps. Available online: https://d-maps.com/carte.php?num_car=8072&lang=en (accessed on 22 December 2022).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c ox-idase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Gutiérrez, M.A.C.; Vivero, R.J.; Vélez, I.D.; Porter, C.H.; Uribe, S. DNA Barcoding for the Identification of Sand Fly Species (Diptera, Psychodidae, Phlebotominae) in Colombia. PLoS ONE 2014, 9, e85496. [Google Scholar] [CrossRef]
- Gomes, B.; Purkait, B.; Deb, R.M.; Rama, A.; Singh, R.P.; Foster, G.M.; Coleman, M.; Kumar, V.; Paine, M.; Das, P.; et al. Knockdown resistance mutations predict DDT resistance and pyrethroid tolerance in the visceral leishmaniasis vector Phlebotomus argentipes. PLoS Negl. Trop. Dis. 2017, 11, e0005504. [Google Scholar] [CrossRef] [Green Version]
- Galletti, E.; Bonilauri, P.; Bardasi, L.; Fontana, M.C.; Ramini, M.; Renzi, M.; Dosa, G.; Merialdi, G. Development of a minor groove binding probe based real-time PCR for the diagnosis and quantification of Leishmania infantum in dog specimens. Res. Veter Sci. 2011, 91, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.J.; Lanciotti, R.S. Consensus Amplification and Novel Multiplex Sequencing Method for S Segment Species Identification of 47 Viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus Genera of the Family Bunyaviridae. J. Clin. Microbiol. 2009, 47, 2398–2404. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ruiz, M.; Collao, X.; Navarro-Marí, J.-M.; Tenorio, A. Reversetranscription, real-time PCR assay for detection of Toscana virus. J. Clin. Virol. 2007, 39, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Corradetti, A. Phlebotomus and leishmaniasis in North-Central Italy (Apennine Region). Sci. Rep. Ist. Super. Sanità 1962, 2, 103–109. [Google Scholar]
- Defilippo, F.; Carrera, M.; Lelli, D.; Canziani, S.; Moreno, A.; Sozzi, E.; Manarolla, G.; Chiari, M.; Marco, F.; Cerioli, M.P.; et al. Distribution of Phlebotomine Sand Flies (Diptera: Psychodidae) in the Lombardy Region, Northern Italy. Insects 2022, 13, 463. [Google Scholar] [CrossRef]
- Michelutti, A.; Toniolo, F.; Bertola, M.; Grillini, M.; Simonato, G.; Ravagnan, S.; Montarsi, F. Occurrence of Phlebotomine sand flies (Diptera: Psychodidae) in the northeastern plain of Italy. Parasites Vectors 2021, 14, 164. [Google Scholar] [CrossRef]
- Calzolari, M.; Carra, E.; Rugna, G.; Bonilauri, P.; Bergamini, F.; Bellini, R.; Varani, S.; Dottori, M. Isolation and Molecular Typing of Leishmania infantum from Phlebotomus perfiliewi in a Re-Emerging Focus of Leishmaniasis, Northeastern Italy. Microorganisms 2019, 7, 644. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Romeo, G.; Bergamini, F.; Dottori, M.; Rugna, G.; Carra, E. Host preference and Leishmania infantum natural infection of the sand fly Phlebotomus perfiliewi in northern Italy. Acta Trop. 2022, 226, 106246. [Google Scholar] [CrossRef]
- Rath, C.T.; Schnellrath, L.C.; Damaso, C.R.; De Arruda, L.B.; Vasconcelos, P.F.D.C.; Gomes, C.; Laurenti, M.D.; Silva, T.C.C.; Vivarini, D.C.; Fasel, N.; et al. Amazonian Phlebovirus (Bunyaviridae) potentiates the infection of Leishmania (Leishmania) amazonensis: Role of the PKR/IFN1/IL-10 axis. PLoS Negl. Trop. Dis. 2019, 13, e0007500. [Google Scholar] [CrossRef] [Green Version]
- Lavagino, A.; Ansaldi, G. Susceptibility tests on Phlebotomus perniciosus and Phlebotomus perfiliewi wild populations in Sicily. Parassitologia 1991, 33, 349–351. [Google Scholar]
- Maroli, M.; Cianchi, T.; Bianchi, R.; Khoury, C. Testing insecticide susceptibility of Phlebotomus perniciosus and P. papatasi (Diptera: Psychodidae) in Italy. Ann. Ist. Super. Sanita 2002, 38, 419–423. [Google Scholar]
- Casanova, C.; Costa, A.I.P.; Natal, D. Dispersal pattern of the sand fly Lutzomyia neivai (Diptera: Psychodidae) in a cutaneous leishmaniasis endemic rural area in Southeastern Brazil. Mem. Inst. Oswaldo Cruz. 2005, 100, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Galvis-Ovallos, F.; Casanova, C.; Bergamaschi, D.P.; Galati, E.A.B. A field study of the survival and dispersal pattern of Lutzomyia longipalpis in an endemic area of visceral leishmaniasis in Brazil. PLoS Negl. Trop. Dis. 2018, 12, e0006333. [Google Scholar] [CrossRef] [Green Version]
- Pichler, V.; Bellini, R.; Veronesi, R.; Arnoldi, D.; Rizzoli, A.; Lia, R.P.; Otranto, D.; Montarsi, F.; Carlin, S.; Ballardini, M.; et al. First evidence of resistance to pyrethroid insecticides in Italian Aedes albopictus populations 26 years after invasion. Pest Manag. Sci. 2018, 74, 1319–1327. [Google Scholar] [CrossRef] [Green Version]
- Haas, J.; Hayward, A.; Buer, B.; Maiwald, F.; Nebelsiek, B.; Glaubitz, J.; Bass, C.; Nauen, R. Phylogenomic and functional characterization of an evolutionary conserved cytochrome P450-based insecticide detoxification mechanism in bees. Proc. Natl. Acad. Sci. USA 2022, 119, e2205850119. [Google Scholar] [CrossRef]

{kind=link}
| Location— Province (X, Y) | Date | Environmental Settings | N | Species Composition (%) | kdr L1014F/S | Pathogen Detection | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n1 | perfi | perni | n2 (p) | L (%) | P (n3) | Leish | TOSV | FERMV | ||||||
| Monteveglio—BO (44.480470, 11.084257) | July 2021 | Abandoned villa in farmland, with sparse trees/shrubs, vineyards | 18,579 | 512 | 99.2 | 0.8 | 60 (12) | 100 | 59 (5462) | 24 | 0 | 1 | ||
| Pianoro—BO (44.406653, 11.358506) | July 2021 | Inhabited villa in cattle pasture, sparse trees, grassland | 12,349 | 852 | 100 | 0 | 60 (12) | 100 | 49 (4562) | 21 | 1 | 2 | ||
| Sadurano—FC (44.158437, 11.960940) | August 2021 | Hilly area proximal to farm | 1323 | 122 | 100 | 0 | 60 (12) | 100 | 9 (441) | 2 | 2 | 1 | ||
| Vignola—MO (44.471979, 10.962144) | September 2021 | Area between villas, vineyards | 3455 | 158 | 100 | 0 | 60 (12) | 100 | 15 (1500) | 4 | 0 | 0 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balaska, S.; Calzolari, M.; Grisendi, A.; Scremin, M.; Dottori, M.; Mavridis, K.; Bellini, R.; Vontas, J. Monitoring of Insecticide Resistance Mutations and Pathogen Circulation in Sand Flies from Emilia-Romagna, a Leishmaniasis Endemic Region of Northern Italy. Viruses 2023, 15, 148. https://doi.org/10.3390/v15010148
Balaska S, Calzolari M, Grisendi A, Scremin M, Dottori M, Mavridis K, Bellini R, Vontas J. Monitoring of Insecticide Resistance Mutations and Pathogen Circulation in Sand Flies from Emilia-Romagna, a Leishmaniasis Endemic Region of Northern Italy. Viruses. 2023; 15(1):148. https://doi.org/10.3390/v15010148
Chicago/Turabian StyleBalaska, Sofia, Mattia Calzolari, Annalisa Grisendi, Mara Scremin, Michele Dottori, Konstantinos Mavridis, Romeo Bellini, and John Vontas. 2023. "Monitoring of Insecticide Resistance Mutations and Pathogen Circulation in Sand Flies from Emilia-Romagna, a Leishmaniasis Endemic Region of Northern Italy" Viruses 15, no. 1: 148. https://doi.org/10.3390/v15010148