
The First Human Vulvar Intraepithelial Neoplasia Cell Line with Naturally Infected Episomal HPV18 Genome

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation and Cell Culture
2.2. Short Tandem Repeat (STR) Analysis
2.3. Soft Agar Assay
2.4. Matrigel Three-Dimensional (3D) Culture
2.5. Karyotype Analysis
2.6. Rolling Circle Amplification (RCA) and Enzyme Digestion Reaction
2.7. Construction of Plasmid
2.8. PCR and Quantitative RT-PCR
2.9. Calculation of HPV DNA Copy
2.10. DNA Damage Response and Western Blotting Analysis
2.11. Hematoxylin-Eosin (H&E) Staining
2.12. DAB Staining
2.13. Immunofluorescence Assay
2.14. Air-Liquid Interface (ALI) 3D Culture
2.15. Viral Particle Visualization by Transmission Electron Microscopy
2.16. Statistical Analysis
3. Results
3.1. Establishment of Human Vulvar Intraepithelial Neoplasia (VIN) Cell Line
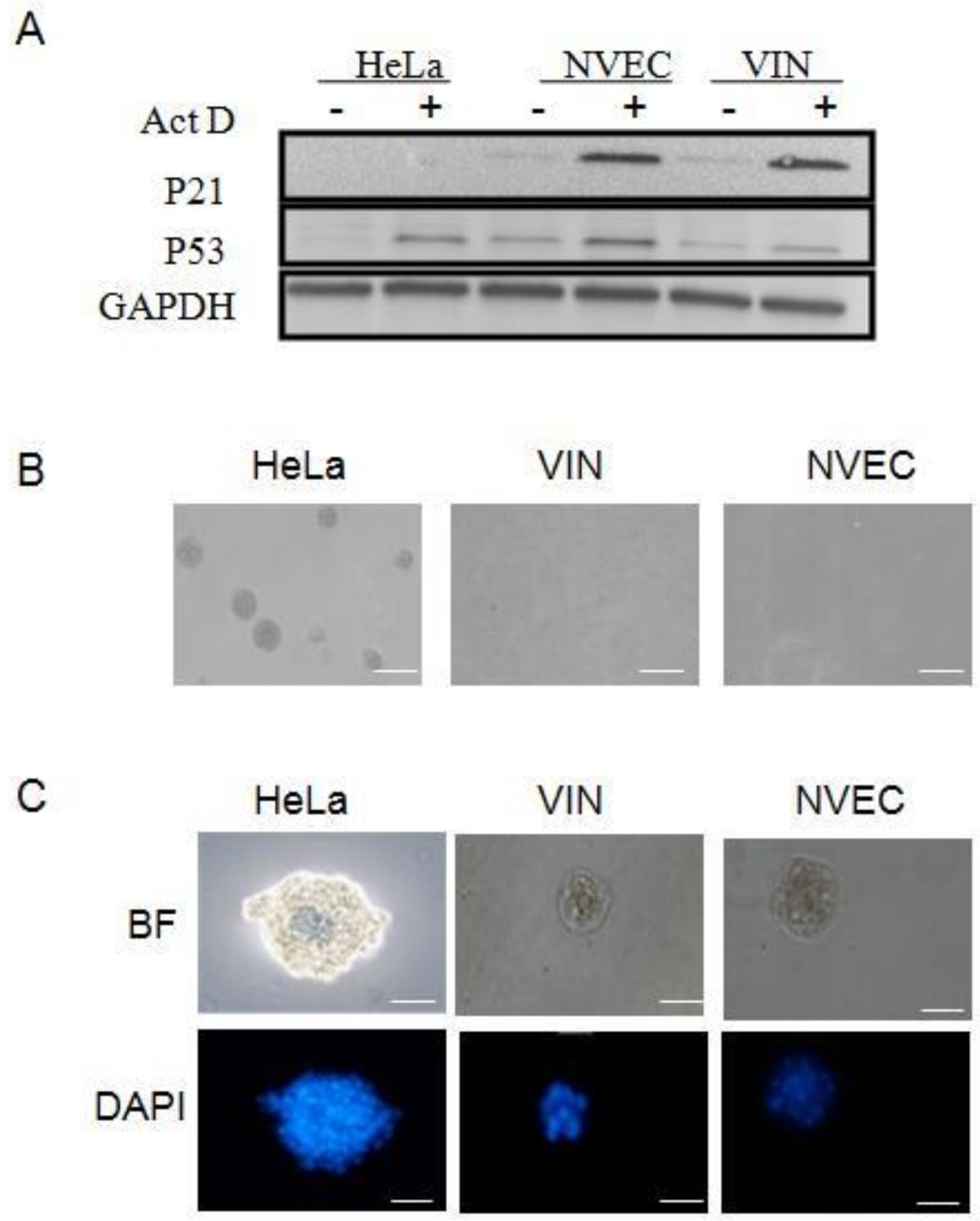
3.2. Different Biological Characteristics in VIN Cells Compared to HPV18 Positive Cancer Cell Line
3.3. Squamous Epithelium and Viral Specific Markers in VIN Cells
3.4. Complex Karyotypes of VIN Cells
3.5. Episomal HPV18 Genome in VIN Cells
3.6. Viral Life Cycle of Naturally Infected HPV18 in VIN Cells under Air-Liquid (ALI) 3D Cultures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bzhalava, D.; Eklund, C.; Dillner, J. International standardization and classification of human papillomavirus types. Virology 2015, 476, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Della Fera, A.N.; Warburton, A.; Coursey, T.L.; Khurana, S.; McBride, A.A. Persistent Human Papillomavirus Infection. Viruses 2021, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.T. Model systems to study the life cycle of human papillomaviruses and HPV-associated cancers. Virol. Sin. 2015, 30, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.S.; Moore, D.W.; Broker, T.R.; Chow, L.T. Vorinostat, a pan-HDAC inhibitor, abrogates productive HPV-18 DNA amplification. Proc. Natl. Acad. Sci. USA 2018, 115, E11138–E11147. [Google Scholar] [CrossRef] [PubMed]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Nooij, L.S.; Ter Haar, N.T.; Ruano, D.; Rakislova, N.; van Wezel, T.; Smit, V.; Trimbos, B.; Ordi, J.; van Poelgeest, M.I.E.; Bosse, T. Genomic Characterization of Vulvar (Pre)cancers Identifies Distinct Molecular Subtypes with Prognostic Significance. Clin. Cancer Res. 2017, 23, 6781–6789. [Google Scholar] [CrossRef]
- Williams, A.; Syed, S.; Velangi, S.; Ganesan, R. New Directions in Vulvar Cancer Pathology. Curr. Oncol. Rep. 2019, 21, 88. [Google Scholar] [CrossRef]
- De Vuyst, H.; Clifford, G.M.; Nascimento, M.C.; Madeleine, M.M.; Franceschi, S. Prevalence and type distribution of human papillomavirus in carcinoma and intraepithelial neoplasia of the vulva, vagina and anus: A meta-analysis. Int. J. Cancer 2009, 124, 1626–1636. [Google Scholar] [CrossRef]
- Bertoli, H.K.; Thomsen, L.T.; Iftner, T.; Dehlendorff, C.; Kjaer, S.K. Risk of vulvar, vaginal and anal high-grade intraepithelial neoplasia and cancer according to cervical human papillomavirus (HPV) status: A population-based prospective cohort study. Gynecol. Oncol. 2020, 157, 456–462. [Google Scholar] [CrossRef]
- Vinokurova, S.; Wentzensen, N.; Einenkel, J.; Klaes, R.; Ziegert, C.; Melsheimer, P.; Sartor, H.; Horn, L.C.; Hockel, M.; von Knebel Doeberitz, M. Clonal history of papillomavirus-induced dysplasia in the female lower genital tract. J. Natl. Cancer Inst. 2005, 97, 1816–1821. [Google Scholar] [CrossRef] [Green Version]
- Stanley, M.A.; Browne, H.M.; Appleby, M.; Minson, A.C. Properties of a non-tumorigenic human cervical keratinocyte cell line. Int. J. Cancer 1989, 43, 672–676. [Google Scholar] [CrossRef]
- Hummel, M.; Hudson, J.B.; Laimins, L.A. Differentiation-induced and constitutive transcription of human papillomavirus type 31b in cell lines containing viral episomes. J. Virol. 1992, 66, 6070–6080. [Google Scholar] [CrossRef]
- Morgan, E.L.; Wasson, C.W.; Hanson, L.; Kealy, D.; Pentland, I.; McGuire, V.; Scarpini, C.; Coleman, N.; Arthur, J.S.C.; Parish, J.L.; et al. STAT3 activation by E6 is essential for the differentiation-dependent HPV18 life cycle. PLoS Pathog. 2018, 14, e1006975. [Google Scholar] [CrossRef]
- Liu, X.; Ory, V.; Chapman, S.; Yuan, H.; Albanese, C.; Kallakury, B.; Timofeeva, O.A.; Nealon, C.; Dakic, A.; Simic, V.; et al. ROCK inhibitor and feeder cells induce the conditional reprogramming of epithelial cells. Am. J. Pathol. 2012, 180, 599–607. [Google Scholar] [CrossRef]
- Liu, X.; Krawczyk, E.; Suprynowicz, F.A.; Palechor-Ceron, N.; Yuan, H.; Dakic, A.; Simic, V.; Zheng, Y.L.; Sripadhan, P.; Chen, C.; et al. Conditional reprogramming and long-term expansion of normal and tumor cells from human biospecimens. Nat. Protoc. 2017, 12, 439–451. [Google Scholar] [CrossRef]
- Suprynowicz, F.A.; Upadhyay, G.; Krawczyk, E.; Kramer, S.C.; Hebert, J.D.; Liu, X.; Yuan, H.; Cheluvaraju, C.; Clapp, P.W.; Boucher, R.C., Jr.; et al. Conditionally reprogrammed cells represent a stem-like state of adult epithelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, 20035–20040. [Google Scholar] [CrossRef]
- Xia, S.; Wu, M.; Chen, S.; Zhang, T.; Ye, L.; Liu, J.; Li, H. Long Term Culture of Human Kidney Proximal Tubule Epithelial Cells Maintains Lineage Functions and Serves as an Ex vivo Model for Coronavirus Associated Kidney Injury. Virol. Sin. 2020, 35, 311–320. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, Y.; Guo, J.; Dai, Y.; Ye, L.; Qiu, J.; Zeng, Z.; Wu, X.; Xing, Y.; Long, X.; et al. Ex vivo 2D and 3D HSV-2 infection model using human normal vaginal epithelial cells. Oncotarget 2017, 8, 15267–15282. [Google Scholar] [CrossRef]
- Lee, G.Y.; Kenny, P.A.; Lee, E.H.; Bissell, M.J. Three-dimensional culture models of normal and malignant breast epithelial cells. Nat. Methods 2007, 4, 359–365. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, L.; Zhong, Y.; Zhu, C.; Zhao, K.; Teng, Y.; Cheng, X.; Chen, Q.; Xia, Y. A Novel Mouse Model Harboring Hepatitis B Virus Covalently Closed Circular DNA. Cell Mol. Gastroenterol. Hepatol. 2022, 13, 1001–1017. [Google Scholar] [CrossRef]
- Banerjee, N.S.; Wang, H.K.; Beadle, J.R.; Hostetler, K.Y.; Chow, L.T. Evaluation of ODE-Bn-PMEG, an acyclic nucleoside phosphonate prodrug, as an antiviral against productive HPV infection in 3D organotypic epithelial cultures. Antiv. Res. 2018, 150, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Wu, M.; Zhou, X.; Zhang, X.; Ye, L.; Zhang, K.; Kang, Y.; Liu, J.; Zhang, Y.; Wu, W.; et al. Treating intrauterine adhesion using conditionally reprogrammed physiological endometrial epithelial cells. Stem Cell Res. Ther. 2022, 13, 178. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.; Liu, X.; Meyers, C.; Schlegel, R.; McBride, A.A. Human keratinocytes are efficiently immortalized by a Rho kinase inhibitor. J. Clin. Investig. 2010, 120, 2619–2626. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.A.; Demers, G.W.; Etscheid, B.G.; Galloway, D.A. The ability of human papillomavirus E6 proteins to target p53 for degradation in vivo correlates with their ability to abrogate actinomycin D-induced growth arrest. J. Virol. 1994, 68, 5698–5705. [Google Scholar] [CrossRef]
- Filho, S.M.A.; Bertoni, N.; Brant, A.C.; Vidal, J.; Felix, S.P.; Cavalcanti, S.M.B.; Carestiato, F.N.; Martins, L.F.L.; Almeida, L.M.; Moreira, M.A.M. Methylation at 3’LCR of HPV16 can be affected by patient age and disruption of E1 or E2 genes. Virus Res. 2017, 232, 48–53. [Google Scholar] [CrossRef]
- Cheung, J.L.; Lo, K.W.; Cheung, T.H.; Tang, J.W.; Chan, P.K. Viral load, E2 gene disruption status, and lineage of human papillomavirus type 16 infection in cervical neoplasia. J. Infect. Dis. 2006, 194, 1706–1712. [Google Scholar] [CrossRef]
- Egawa, N.; Wang, Q.; Griffin, H.M.; Murakami, I.; Jackson, D.; Mahmood, R.; Doorbar, J. HPV16 and 18 genome amplification show different E4-dependence, with 16E4 enhancing E1 nuclear accumulation and replicative efficiency via its cell cycle arrest and kinase activation functions. PLoS Pathog. 2017, 13, e1006282. [Google Scholar] [CrossRef]
- Warburton, A.; Della Fera, A.N.; McBride, A.A. Dangerous Liaisons: Long-Term Replication with an Extrachromosomal HPV Genome. Viruses 2021, 13, 1846. [Google Scholar] [CrossRef]
- Bedell, M.A.; Hudson, J.B.; Golub, T.R.; Turyk, M.E.; Hosken, M.; Wilbanks, G.D.; Laimins, L.A. Amplification of human papillomavirus genomes in vitro is dependent on epithelial differentiation. J. Virol. 1991, 65, 2254–2260. [Google Scholar] [CrossRef]
- Jeon, S.; Allen-Hoffmann, B.L.; Lambert, P.F. Integration of human papillomavirus type 16 into the human genome correlates with a selective growth advantage of cells. J. Virol. 1995, 69, 2989–2997. [Google Scholar] [CrossRef] [Green Version]
- Stanley, M. Host defence and persistent human papillomavirus infection. Curr. Opin. Virol. 2021, 51, 106–110. [Google Scholar] [CrossRef]
- Spriggs, C.C.; Laimins, L.A. Human Papillomavirus and the DNA Damage Response: Exploiting Host Repair Pathways for Viral Replication. Viruses 2017, 9, 232. [Google Scholar] [CrossRef]
- Regan, J.A.; Laimins, L.A. Viral transformation of epithelial cells. Methods Mol. Biol. 2013, 945, 449–465. [Google Scholar] [CrossRef]
- Lambert, P.F.; Ozbun, M.A.; Collins, A.; Holmgren, S.; Lee, D.; Nakahara, T. Using an immortalized cell line to study the HPV life cycle in organotypic “raft” cultures. Methods Mol. Med. 2005, 119, 141–155. [Google Scholar] [CrossRef]
- Galloway, D.A. Human papillomaviruses: A growing field. Genes Dev. 2009, 23, 138–142. [Google Scholar] [CrossRef]
- McBride, A.A. Human papillomaviruses: Diversity, infection and host interactions. Nat. Rev. Microbiol. 2022, 20, 95–108. [Google Scholar] [CrossRef]
- Dzobo, K. The Role of Viruses in Carcinogenesis and Molecular Targeting: From Infection to Being a Component of the Tumor Microenvironment. OMICS 2021, 25, 358–371. [Google Scholar] [CrossRef]
- Kaynarcalidan, O.; Oguzoglu, T.C. The oncogenic pathways of papillomaviruses. Vet. Comp. Oncol. 2021, 19, 7–16. [Google Scholar] [CrossRef]
- Ghittoni, R.; Accardi, R.; Hasan, U.; Gheit, T.; Sylla, B.; Tommasino, M. The biological properties of E6 and E7 oncoproteins from human papillomaviruses. Virus Genes 2010, 40, 1–13. [Google Scholar] [CrossRef]
- McLaughlin-Drubin, M.E.; Munger, K. Oncogenic activities of human papillomaviruses. Virus Res. 2009, 143, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Coursey, T.L.; McBride, A.A. Development of Keratinocyte Cell Lines Containing Extrachromosomal Human Papillomavirus Genomes. Curr. Protoc. 2021, 1, e235. [Google Scholar] [CrossRef] [PubMed]
- Bienkowska-Haba, M.; Luszczek, W.; Myers, J.E.; Keiffer, T.R.; DiGiuseppe, S.; Polk, P.; Bodily, J.M.; Scott, R.S.; Sapp, M. A new cell culture model to genetically dissect the complete human papillomavirus life cycle. PLoS Pathog. 2018, 14, e1006846. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.S.; McBride, A.A. Human Papillomavirus Quasivirus Production and Infection of Primary Human Keratinocytes. Curr. Protoc. Microbiol. 2020, 57, e101. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J. Model systems of human papillomavirus-associated disease. J. Pathol. 2016, 238, 166–179. [Google Scholar] [CrossRef]
- Wang, H.K.; Duffy, A.A.; Broker, T.R.; Chow, L.T. Robust production and passaging of infectious HPV in squamous epithelium of primary human keratinocytes. Genes Dev. 2009, 23, 181–194. [Google Scholar] [CrossRef]
- Xia, S.; Liu, J.; Yang, Y.; Wu, M.; Ye, L.; Chen, S.; Zhang, T.; Zeng, Z.; Zhang, K.; Cai, K.; et al. Coupled CRC 2D and ALI 3D Cultures Express Receptors of Emerging Viruses and Are More Suitable for the Study of Viral Infections Compared to Conventional Cell Lines. Stem Cells Int. 2020, 2020, 2421689. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Sequences (5’-to-3’) | Amplicon Size (bp) |
|---|---|---|
| GP5+/GP6+-F | TTTGTTACTGTGGTAGATACTAC | 150 |
| GP5+/GP6+-R | GAAAAATAAACTGTAAATCATATTC | |
| HPV18-F | CCGAGCACGACAGGAACGACT | 172 |
| HPV18-R | TCGTTTTCTTCCTCTGAGTCGCTT | |
| HPV18 E1-F | ATGGCTGATCCAGAAG | 205 |
| HPV18 E1-R | ATGCCTGTGCTGTCTCTAGC | |
| HPV18 E2 (amino 1)-F | AGACACCGAAGGAAACCCTTT | 223 |
| HPV18 E2 (amino 1)-R | GCTTTATGTGCTTTACTTTTTGA | |
| HPV18 E2 (amino 2)-F | TGCAAGACACATGCGAGGAA | 157 |
| HPV18 E2 (amino 2)-R | CATGTTCCTGCATCAGTCATAT | |
| HPV18 E2 (hinge)-F | AAAATATGGGAACACAGGTACG | 192 |
| HPV18 E2 (hinge)-R | GCCGACGTCTGGCCGTAGGTCT | |
| HPV18 E2 (carboxyl)-F | TACAGGCAACAACAAAAGACG | 177 |
| HPV18 E2 (carboxyl)-R | CCTGTTTTTTCATTGCCTGC | |
| HPV18 E4-F | TGTGCAGTACCAGTGACGAC | 204 |
| HPV18 E4-R | GTGTAGCTGCACCGAGAAGT | |
| HPV18 E5-F | CCGCTTTTGCCATCTGTCTG | 85 |
| HPV18 E5-R | CTGTGGCAGGGGACGTTATT | |
| HPV18 L1-F | GCCGCCACGTCTAATGTTTC | 149 |
| HPV18 L1-R | CCCTGTGATAAAGGACGCGA | |
| HPV18 L2-F | ATGACAACCCGGCCTTTGAG | 165 |
| HPV18 L2-R | ACATAGTTGCCCGTTGACCT | |
| HPV18 LCR-F | AGGGAGTAACCGAAAACGGTC | 70 |
| HPV18 LCR-R | GTATTGTGGTGTGTTTCTCAC | |
| HPV18 E6/E7-F | ACGACAGGAACGACTCCAAC | 291 |
| HPV18 E6/E7-R | AGGTCGTCTGCTGAGCTTTC | |
| β-actin-F | GCACGGCATCGTCACCAACT | 142 |
| β-actin-R | CATCTTCTCGCGGTTGGCCT | |
| HPV11-F | CGCAGAGATATATGCATATGC | 80 |
| HPV11-R | AGTTCTAAGCAACAGGCACAC | |
| HPV16-F | GTCAAAAGCCACTGTGTCCT | 499 |
| HPV16-R | CCATCCATTACATCCCGTAC | |
| HPV16 E6-F | ATGTTTCAGGACCCACAGGA | 453 |
| HPV16 E6-R | CAGCTGGGTTTCTCTACGTGTT | |
| HPV16 E7-F | ATGCATGGAGATACACCTAC | 252 |
| HPV16 E7-R | CATTAACAGGTCTTCCAAAG | |
| HPV33-F | AACGCCATGAGAGGACACAAG | 211 |
| HPV33-R | ACACATAAACGAACTGTGGTG | |
| HPV53 E6-F | GGGTATCCGTATGGAGTGTGC | 263 |
| HPV53 E6-R | GTTGTGTGTCTCCAGCATGTC | |
| HPV18 E6-F | CGCTTTGAGGATCCAACACG | 439 |
| HPV18 E6-R | GTTCCTGTCGTGCTCGGTTG | |
| HPV18 E7-F | GTCACGAGCAATTAAGCGAC | 212 |
| HPV18 E7-R | CACAAAGGACAGGGTGTTCA | |
| GAPDH-F | CTGGGCTACACTGAGCACC | 101 |
| GAPDH-R | AAGTGGTCGTTGAGGGCAATG | |
| HPV18 E2 (fl)-F | GTTGTAAAACGACGGCCAGTATGCAGACACCGAAGGAA | 1098 |
| HPV18 E2 (fl)-R | TTACATTGTCATGTATCCCACC | |
| HPV18 E7 (fl)-F | TGGGATACATGACAATGTAAATGCATGGACCTAAGGCA | 318 |
| HPV18 E7 (fl)-R | GATCCCCGGGTACCGAGCTCTTACTGCTGGGATGCACA |
| Antibody | Species | Vendor/Source | Catalogue # |
|---|---|---|---|
| CK14 | mouse | Santa Cruz Biotech | sc-23878 |
| CK18 P21 P53 P63 Ki67 Filaggrin Involucrin Involucrin HPV18 E7 HPV18 L1 | mouse Rabbit mouse Rabbit mouse mouse Rabbit mouse mouse Rabbit | Santa Cruz Biotech Abcam Santa Cruz Biotech Abcam Cell Signaling Santa Cruz Biotech Abcam Santa Cruz Biotech Santa Cruz Biotech biorbyt | sc-32329 ab-109199 sc-126 ab-124762 9449S sc-66192 ab-227530 sc-21748 sc-365035 p06794 |
| GAPDH | mouse | Santa Cruz Biotech | sc-365062 |
| HeLa | VIN | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene | Ct | Log Copies | Copies | E2/E7 | Ct | Log Copies | Copies | E2/E7 |
| E2 amino1 | 22.48 | 7.6 | 39,959,130.32 | 0.62 | 18.56 | 9.01 | 1,014,918,805 | 1.94 |
| E2 amino2 | 28.75 | 5.23 | 169,033.01 | 0.00 | 18.90 | 8.74 | 555,874,925.75 | 1.06 |
| E2 hinge | 27.80 | 5.57 | 371,777.79 | 0.01 | 18.64 | 8.84 | 691,655,509.02 | 1.32 |
| E2 carboxy1 | 26.95 | 5.90 | 795,092.58 | 0.01 | 18.73 | 8.84 | 693,065,846.55 | 1.32 |
| E7 | 20.64 | 7.82 | 65,874,356.7 | - | 18.13 | 8.72 | 524,162,224 | - |
| Enzymes | Number of Cutting Fragments | Size of Cutting Fragment (bp) |
|---|---|---|
| EcoRⅠ | 1 | 7857 |
| BamHⅠ | 2 | 1047, 6810 |
| NdeⅠ | 3 | 1075, 2342, 4440 |
| EcoRⅠ+ EcoRⅤ | 2 | 2229, 5628 |
| EcoR Ⅴ+ SphⅠ | 2 | 3486, 4371 |
| EcoRⅠ+ SphⅠ | 2 | 2142, 5715 |
| EcoRⅠ+ BamHⅠ | 3 | 1047, 2321, 4489 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.; Zhang, X.; Kang, Y.; Zhu, Y.; Su, Z.; Liu, J.; Zhang, W.; Chen, H.; Li, H. The First Human Vulvar Intraepithelial Neoplasia Cell Line with Naturally Infected Episomal HPV18 Genome. Viruses 2022, 14, 2054. https://doi.org/10.3390/v14092054
Wu M, Zhang X, Kang Y, Zhu Y, Su Z, Liu J, Zhang W, Chen H, Li H. The First Human Vulvar Intraepithelial Neoplasia Cell Line with Naturally Infected Episomal HPV18 Genome. Viruses. 2022; 14(9):2054. https://doi.org/10.3390/v14092054
Chicago/Turabian StyleWu, Ming, Xiu Zhang, Yiyi Kang, Yaqi Zhu, Zhaoyu Su, Jun Liu, Wei Zhang, Hong Chen, and Hui Li. 2022. "The First Human Vulvar Intraepithelial Neoplasia Cell Line with Naturally Infected Episomal HPV18 Genome" Viruses 14, no. 9: 2054. https://doi.org/10.3390/v14092054