1. Introduction
Recent decades have seen the emergence and reemergence of a number of significant arboviral diseases such as dengue, Zika, West Nile, yellow fever, and chikungunya [
1,
2,
3]. Arboviruses, which are transmitted through the bite of an infected arthropod vector such as a tick or mosquito, are expected to become more significant in the future and it is predicted that climate change will increase incidence of disease by impacting vector geographical range, feeding behavior, and survival [
4,
5,
6]. New ways of protection from these diseases are needed as vaccines are not available for many of these diseases and there is increasing insecticide resistance in some vectors [
7,
8,
9].
An alternative to traditional means of controlling arboviral diseases is to prevent productive infection of the vector. Tissue barriers in the vector such as the midgut present an obstacle for viruses to overcome and inhibiting escape from midgut tissue would prevent disseminated infection and thus the spread of these viruses [
10]. Several pathways and processes may be considered which when altered could prevent disseminated infection, and improved knowledge of these pathways could lead to new strategies of vector infection control. One cellular process which shows potential promise in preventing disseminated infection is apoptosis, a specific type of programmed cell death that has been shown to be an important antiviral pathway in many organisms including insects [
11,
12]. The insect apoptosis pathway has been best studied in
Drosophila melanogaster, but a similar pathway has been demonstrated in the disease vector
Aedes aegypti [
13,
14,
15,
16]. During apoptosis, activated initiator caspases cleave and activate effector caspases, which are responsible for cleaving target proteins in the cell, leading to death. Activation of initiator caspases is prevented by inhibitor of apoptosis (IAP) proteins, the action of which can be overcome by IAP antagonists such as
Drosophila Reaper, which can bind directly to IAP proteins via the IAP binding motif (IBM) found at its amino terminus [
17]. The result is a carefully controlled mechanism which prevents unnecessary cell death but promotes cell death in response to activating stimuli such as viral infection.
The role of apoptosis in protecting insects against viral infections brings up the question of what role it plays in vector competence for arboviruses. Several studies have implicated apoptosis as a significant factor in preventing viral escape from the midgut. For example, enhanced midgut apoptosis has been associated with a
Culex pipiens pipiens strain of mosquitoes that were refractory to West Nile virus infection [
18]. Additionally, previous work by our group showed that a SINV construct that expressed Reaper from a duplicated subgenomic promoter was initially less able to infect and disseminate from the
Ae. aegypti midgut, although viruses with deletions in the
reaper insert rapidly predominated [
19]. Consistent with these results, one study showed that some pro-apoptotic genes were more highly expressed in a refractory strain compared to a susceptible strain of
Ae. aegypti [
20], while another study found increased rapid induction of apoptosis in mosquitoes that were less susceptible to dengue virus serotype 2 (DENV-2) compared to a more susceptible strain [
21]. However, it has also been suggested that excessive apoptosis may weaken the barrier that the midgut provides and thus allow viruses to pass through more easily. One study found that knocking down AeIAP1 expression in
Ae. aegypti and then feeding them with SINV led to increased midgut infection and virus dissemination [
22]. Due to a high rate of mosquito mortality in these AeIAP1 knockdown mosquitoes, it was hypothesized that the level of apoptosis was drastically increased, greatly reducing the structural integrity of the midgut. It is possible that some level of apoptosis is critical to preventing virus passage through tissues but if apoptosis levels are too high, viral spread is promoted through gaps in the structure.
A previous series of studies by our group aimed to determine how inserting the
Drosophila pro-apoptotic gene
reaper into SINV and then infecting
Ae. aegypti would affect rates of disseminated infection [
19,
23]. The construct used in these earlier studies, called MRE/rpr, expressed Reaper via a duplicated subgenomic promoter located between the nonstructural and structural genes. MRE/rpr strongly induced apoptosis and had decreased virus yield compared to control viruses in cultured mosquito cells [
23]. When mosquitoes were fed MRE/rpr, apoptosis was observed in midgut cells and it was found that there was less disseminated infection compared to control virus at early time points, but by 7 days post-blood meal (PBM), there was no significant difference between MRE/rpr and control. The reason for this was found to be that a significant proportion of the MRE/rpr population by 7 days PBM had deletions in the
reaper insert, rendering it non-functional [
19]. This result indicated strong negative selection against Reaper expression and supported a role for apoptosis as an antiviral response. However, the lack of stability of Reaper expression from MRE/rpr complicates the interpretation of these results.
To potentially deliver more stable expression of Reaper protein, we decided to insert the
reaper gene into the structural ORF of SINV, a method which has been previously shown to allow more stable insertions into the SINV genome [
24,
25]. By inserting the sequence into the structural ORF as a continuous open reading frame between the capsid gene and PE2, as opposed to the duplicated subgenomic promoter region, we hoped to increase selective pressure for retaining the insert because deletions in
reaper would be more likely to negatively impact the expression of critical viral structural proteins. To allow proper synthesis of the viral structural proteins, we utilized the autoproteolytic function of the SINV capsid protein and the ribosomal skipping function of foot and mouth disease virus (FMDV) 2A [
26] to cotranslationally release the Reaper protein. Additionally, we employed a ubiquitin fusion strategy which has been used successfully to generate proteins with precise N-terminal sequences [
27,
28]. This ensured that we would not impact the N-terminus of Reaper, which has been shown to be critical for binding IAPs [
17].
Using this SINV construct, called MRE/rprORF, we then re-examined the effect of Reaper expression on establishment of midgut infection and dissemination from the midgut. Our results provide deeper insights into the effects of apoptosis on SINV infection in Ae. aegypti.
2. Materials and Methods
2.1. Cells
BHK-21 cells were maintained at 37 °C with 5% CO2 in Dulbecco modified Eagle medium (DMEM, Gibco, Waltham, MA, USA) plus 10% fetal bovine serum (FBS, Atlanta Biologicals, Minneapolis, MN, USA). C6/36 cells were maintained at 27 °C in Liebovitz’s medium (Gibco) plus 10% FBS.
2.2. Insect Rearing
Orlando strain Ae. aegypti mosquitoes (obtained in 2008 from James Becnel, USDA ARS, Gainesville, FL, USA) were reared in a 27 °C incubator with 80% humidity and a 12-h light-dark cycle. To obtain eggs used in experiments, females were allowed to feed on defibrinated sheep’s blood (Colorado Serum Company, Denver, CO, USA) using a Hemotek feeding system (Hemotek Ltd., Blackburn, UK). Adult mosquitoes were maintained on raisins and water.
2.3. Plasmid Design and Construction
A previously described plasmid containing a fragment of 5′dsMRE16ic extending from the NotI site to the AvrII site as well as microRNA target sites and a 2A self-cleaving peptide sequence ligated into a pGEM-T backbone was used as the starting plasmid [
25]. This plasmid was digested with BstEII and AfIII and the intervening sequence containing the miRNA target sites was replaced with the following sequence (synthesized by Genewiz, South Plainfield NJ) containing ubiquitin (human K48R), the
Drosophila reaper sequence (lacking the initiator methionine), and an HA tag: (TGGAATAGCAAGGGAAAGACCATCAAGACGACGCCCGAAGGGACAGAGGAATGGTCAGCAGCACTCGAGATGCAGATCTTCGTCAAGACGTTAACCGGTAAAACCATAACTCTAGAAGTTGAACCATCCGATACCATCGAAAACGTTAAGGCTAAAATTCAAGACAAGGAAGGCATTCCACCTGATCAACAAAGATTGATCTTTGCCGGTAGGCAGCTTGAGGACGGTAGAACGCTGTCTGATTACAACATTCAGAAGGAGTCCACCCTGCACCTGGTCCTCCGTCTCAGAGGTGGTGCAGTGGCATTCTACATACCCGATCAGGCGACTCTGTTGCGGGAGGCGGAGCAGAAGGAGCAGCAGATCCTTCGCTTGCGGGAGTCACAGTGGAGATTCCTGGCCACCGTCGTCCTGGAAACCCTGCGCCAGTACACTTCATGTCATCCGAAGACCGGAAGAAAGTCCGGCAAATATCGCAAGCCATCGCAATACCCATACGATGTTCCAGATTACGCTGGATCCCAGCTGTTGAATTTTGACCTT). A control plasmid was generated in the same way, but the intervening sequence was replaced with a synthesized sequence not containing
Drosophila reaper but still containing ubiquitin and an HA tag: (TGGAATAGCAAGGGAAAGACCATCAAGACGACGCCCGAAGGGACAGAGGAATGGTCAGCAGCACTCGAGATGCAGATCTTCGTCAAGACGTTAACCGGTAAAACCATAACTCTAGAAGTTGAACCATCCGATACCATCGAAAACGTTAAGGCTAAAATTCAAGACAAGGAAGGCATTCCACCTGATCAACAAAGATTGATCTTTGCCGGTAGGCAGCTTGAGGACGGTAGAACGCTGTCTGATTACAACATTCAGAAGGAGTCCACCCTGCACCTGGTCCTCCGTCTCAGAGGTGGTTATCCATACGATGTTCCAGATTACGCTGGATCCCAGCTGTTGAATTTTGACCTT). Both plasmids were then digested with NotI and AvrII and the purified fragments were ligated into p5′dsMRE16ic which had been digested with the same restriction enzymes. Proper insertion was verified by Sanger sequencing. The generation of MRE/rpr has previously been described [
23].
2.4. Virus Production
Infectious clone plasmids were linearized using AscI (New England Biolabs, Ipswich, MA, USA) and then in vitro transcribed using the MEGAscript SP6 transcription kit (Thermo Fisher Scientific, Waltham, MA, USA) with added cap analog (New England Biolabs). Following transcription, RNA was transfected into BHK-21 cells using Lipofectamine 3000 (Thermo Fisher Scientific). After two days, the media was removed and used to infect a T75 flask of C6/36 cells. After 5 days, the resulting P2 virus stock was frozen in aliquots and titer was determined using TCID50 assay.
2.5. TCID50 Assay
BHK-21 cells were plated at a density of 1 × 10
4 cells per well in a 96-well tissue culture plate in 100 µL of DMEM plus 10% FBS and supplemented with 15 µg per ml of penicillin/streptomycin (Invitrogen, Waltham MA). Mosquito and cell samples were removed from −80 °C and thawed on ice. Samples were then spun to remove debris and DMEM was used to make serial dilutions of each sample. Each dilution was transferred to five wells containing BHK-21 cells. After 5 days, each well was scored for cytopathic effects. The number of wells of each dilution scored as positive was used to determine TCID
50 mL−1 and this was converted to PFU mL
−1 by multiplying by 0.69 [
29].
2.6. Replication Curves
For replication curves in C6/36 cells, the cells were plated at a density of 1 × 106 cells per well in a 6-well plate in 2 mL of Leibovitz’s medium containing 10% FBS. For replication curves in BHK-21 cells, the cells were plated at a density of 5 × 105 cells per well in a 6-well plate in 2 mL DMEM containing 10% FBS. Cells were allowed to recover for 2 h and then were infected with MRE/rprORF, MRE/rpr, or MRE/control at a multiplicity of infection (MOI) of 0.1. The cells were placed on a rocker and the virus was allowed to adsorb for 1 h. The media was then removed, and the cells were rinsed before replacing the media. For the cumulative replication curves, 100 μL of media was sampled for analysis at 1, 2, 3, and 4 dpi. For the non-cumulative replication curves, 100 μL of media was sampled at each time point for analysis and then the remaining media was removed, the cells were rinsed twice, and then 2 mL of media were added. All samples were frozen and stored at −80 °C until analysis by TCID50 assay.
2.7. DNA Fragmentation Assay
C6/36 cells were plated at a density of 2 × 106 cells per well in a 6-well plate in 2 mL Leibovitz’s medium containing 10% FBS. Cells were allowed to recover for 2 h and then were infected with MRE/rprORF, MRE/control, or 5′dsMRE16ic at an MOI of 1. After 48 h, cells were removed from the 6-well plates and pelleted at 500× g. They were washed twice with phosphate buffered saline (140 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4) and then resuspended in 200 μL lysis buffer (0.1 M SDS, 0.1 M Tris pH 8.0, 0.05 M EDTA pH 8.0, 200 mg ml−1 Proteinase K) and incubated at room temperature for 1 h. The samples were then extracted twice with phenol/chloroform and then ethanol precipitated. The pelleted DNA was resuspended in 100 μL Tris-EDTA buffer (10 mM Tris-Cl, 1 mM EDTA) containing 100 μg mL−1 RNase A (Thermo Fisher Scientific) and incubated at room temperature for 5 min. Twenty μL of each sample was loaded into a 1.2% agarose gel containing 0.5 μg mL−1 ethidium bromide. The gel was visualized using an AlphaImager gel imaging system (Alpha Innotech, San Leandro, CA, USA). Sizes of the bands were compared to a Versaladder DNA ladder (Gold Bio, Olivette, MO, USA).
2.8. Mosquito Infection for TCID50 and Caspase Assay
Prior to blood feeding, mosquitoes were placed in cups containing 20–30 females and 10% males and were only provided water for 24 h. MRE/rprORF, MRE/rpr, and MRE/control stocks were diluted to 1.45 × 107 PFU mL−1 with Liebovitz’s medium. The diluted virus stocks were then mixed 1:1 with defibrinated sheep blood and the mosquitoes were then allowed to feed on one of the virus blood mixtures using a Hemotek feeding system for 90 min. Fully engorged females were separated from unfed and partially fed females and males and were given water and raisins to feed ad libitum. At 3, 5, and 7 days PBM, the mosquitoes were cold anesthetized, and the midguts were dissected from the rest of the mosquito (the carcass). The midguts and carcasses used for TCID50 assays were placed in 1.5 mL tubes containing 200 μL DMEM media containing 10% FBS and homogenized using disposable pestles. The samples were then frozen at −80 °C. The midguts used for caspase assays were collected in 30 μL of caspase reaction buffer (20 mM Hepes-KOH, pH 7.5, 50 mM KCl, 1.5 mM MgCl2, 1 mM EDTA, 1 mM EGTA, 1 mM DTT), homogenized with disposable pestles, and stored at −80 °C.
2.9. Immunoblotting
C6/36 cells were plated at a density of 2 × 106 cells per well in a 6-well plate containing 2 mL Leibovitz’s media plus 10% FBS. They were then infected with MRE/rprORF, MRE/control, or 5′dsMRE16ic at an MOI of 10. After 24 or 48 h, the cells were rinsed three times with cold PBS. The plate was placed on ice and 100 μL of cold Laemmli sample buffer (Bio-Rad, Hercules CA) was added to each well. The wells were scraped, and the lysate was collected in a 1.5 mL tube. The samples were then heated to 100 °C for 5 min, centrifuged at 4 °C, and the supernatant was transferred to a new tube. SDS-PAGE was performed using 30 μL of sample with 4–20% Bis-Tris gels (Genscript, Piscataway, NJ, USA) in Tris-MOPS-SDS running buffer (Genscript), and proteins were transferred to PVDF membrane. After blocking for 1 h in 5% dried skim milk in TBST, a 1:1000 dilution of the anti-HA (Biolegend, San Diego, CA, USA) or anti-β-actin (Santa Cruz Biotechnology, Dallas, TX, USA) primary antibody was incubated with the membrane overnight at 4 °C with constant agitation. After incubation, the membrane was washed three times with TBST. A 1:15,000 dilution of goat anti-mouse IgG-horseradish peroxidase secondary antibody (Thermo Fisher Scientific) was added and was incubated for 1 h at room temperature with agitation. Bands were detected using SuperSignal West Pico PLUS Chemiluminescent Substrate (Thermo Fisher Scientific) and visualized using a LI-COR Western Blot Imager.
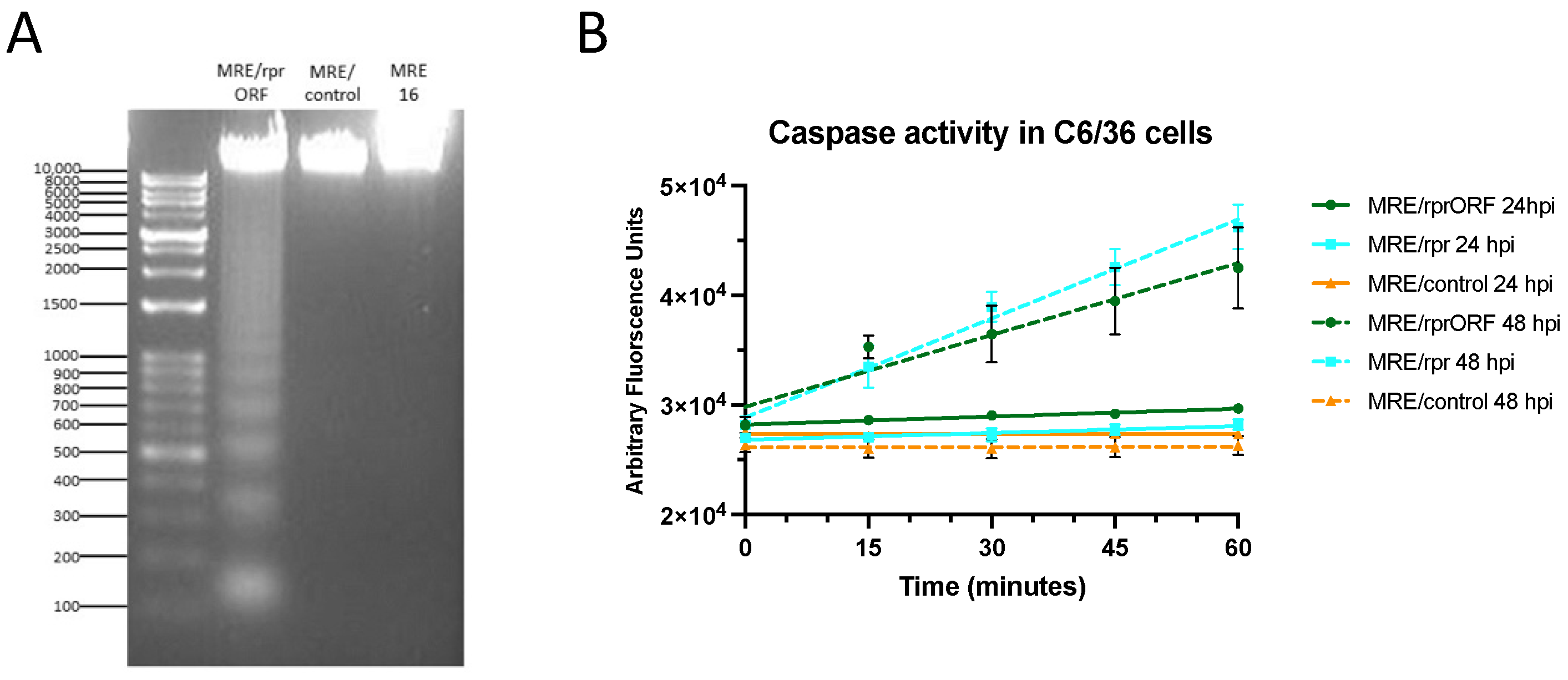
2.10. Caspase Assay
In the cell experiments, C6/36 cells were plated at a density of 2 × 106 cells per well in a 6-well plate in 2 mL Leibovitz’s media plus 10% FBS. The cells were infected with MRE/rprORF, MRE/rpr, or MRE/control at an MOI of 1. At 24 and 48 hpi, the cells were rinsed with PBS and then collected in 400 µL of caspase reaction buffer. Mosquito midgut samples were collected in pools of 8 in caspase reaction buffer and the tissues were disrupted by sonication. The tubes were centrifuged, and the supernatant was moved to a new tube. All samples were frozen at −80 °C until testing. Bradford protein assay (Bio-Rad) was used to determine protein concentration, and cell and mosquito samples were diluted to 100 µg ml−1. A total of 50 µL of each sample were added to white 96-well plates (Costar; Sigma Aldrich, St. Louis, MO, USA) and incubated at 37 °C for 15 min. Then, 10 µL of Ac-DEVD-AFC (ApexBio, Houston, TX, USA) was added to each well at a final concentration of 20 µM. Cleavage of this fluorogenic substrate was monitored at an excitation wavelength of 405 nm and an emission wavelength of 535 nm using a Victor3 1420 Multilabel Counter (Perkin-Elmer, Waltham, MA, USA). Readings were taken at 15 min intervals for a period of 1 h.
2.11. Deep Sequencing of Virus Populations
Total RNA was isolated using TRIzol (Invitrogen) from the carcasses of 10 mosquitoes that became infected with MRE/rprORF and were harvested at 5 or 7 days PBM, as well as from the same MRE/rprORF stock virus that was used in mosquito infections. Samples C6, C9, C20, C24, C36, and C37 were from 5 days PBM, while C11, C17, C18, and C19 were from 7 days PBM. RNA was converted to cDNA using the Improm-II reverse transcription system (Promega) and oligo dT primer. The resulting cDNA was subjected to PCR using Q5 high fidelity polymerase (New England Biolabs) with primers that amplified a ~450 bp region of the MRE/rprORF genome containing the reaper sequence (forward primer: 5′- TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGTACAACATTCAGAAGGAGTCCAC-3′; reverse primer: 5′- GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGTCATATTTCCCAGGATGCAC-3′). The primers also included Illumina overhang adaptor sequences. PCR products were purified and used for index PCR. Following validation and quantification, amplicons were purified, pooled, and sequenced via 2 × 300 bp Illumina MiSeq at the K-State Integrated Genomics Facility.
Paired end reads were remapped to the 5′dsMRE16ic reference genome using BWA 0.7.17-r1198-dirty under the BWA-MEM algorithm [
30]. The resulting SAM file was converted to binary format, sorted, and indexed using SAMtools v1.10 [
31]. Variant calling was done using bam-readcount 0.8.0-unstable-7-625eea2 [
32] with filtering and removal of false-positive calls having minimum base and mapping qualities of less than 30 and 20, respectively. Output from bam-readcount was converted to tab-delimited files which are presented as
Data Set S1. Sequence data have been deposited in the Sequence Read Archive under BioProject ID PRJNA875102.
4. Discussion
In order to better understand the effects of apoptosis on the ability of SINV to infect
Ae. aegypti, we generated a SINV construct that expresses Reaper as an in-frame insertion in the structural ORF and tested how this virus behaved in cell culture and in mosquitoes. Previous work using a SINV construct that expressed Reaper from a duplicated subgenomic promoter (MRE/rpr) found that less mosquitoes developed disseminated infection than a control virus at early time points, but this difference disappeared over time due to deletions that accumulated in the inserted
reaper sequence [
19]. Therefore, we were interested in learning what affect apoptosis would have on SINV infection if the Reaper protein was expressed in a potentially more stable fashion.
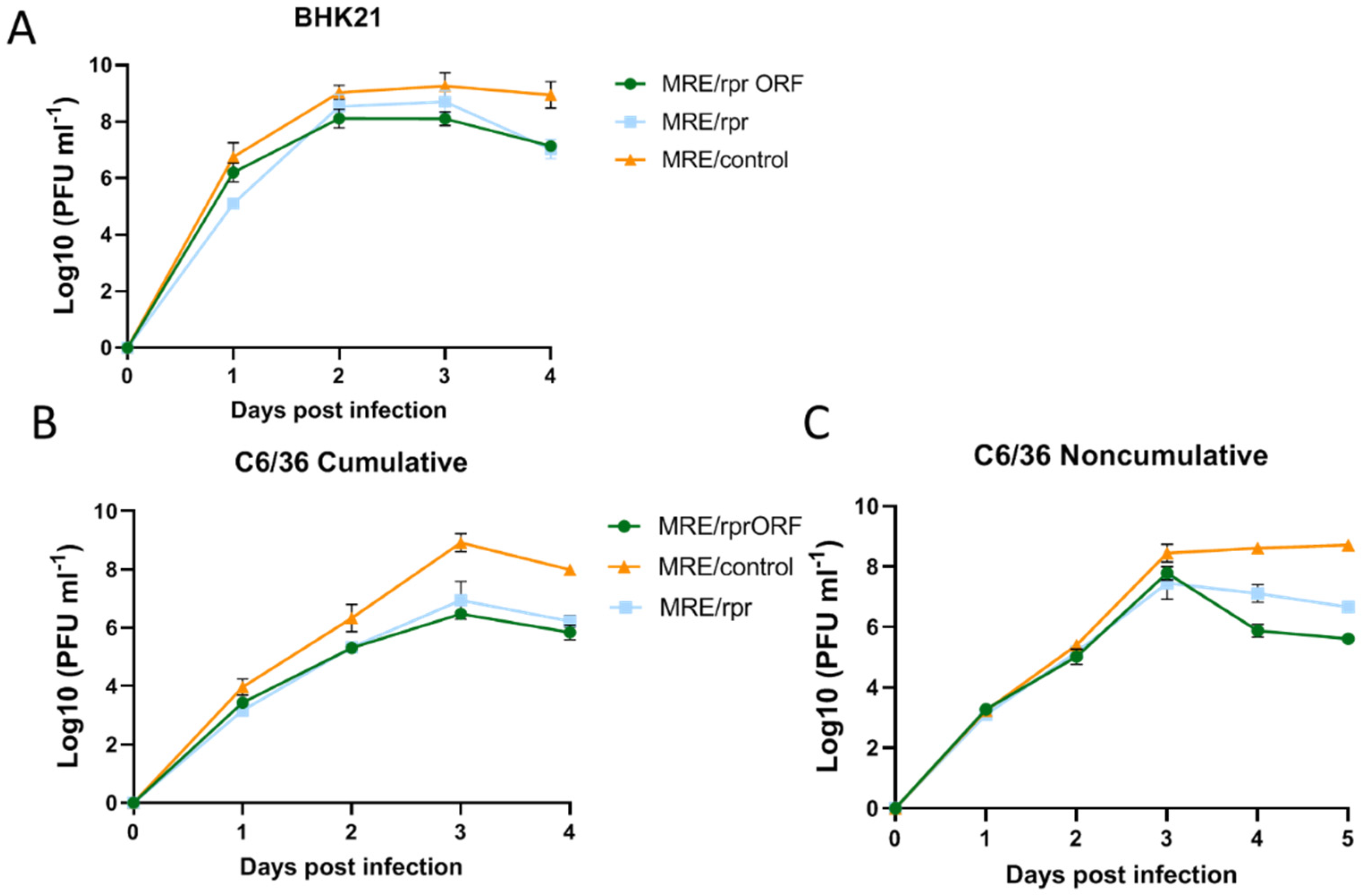
The results of our study showed that expressing the Reaper protein from the structural ORF of SINV using a ubiquitin fusion strategy was successful. We were able to detect Reaper protein in MRE/rprORF-infected cells by immunoblotting, as well as increased markers of apoptosis in both infected cells and mosquito midgut. We detected what was likely the free Reaper protein at 48 hpi but also detected what was most likely the uncleaved ubiquitin-Reaper fusion protein at both 24 hpi and 48 hpi. Thus, it appears that cleavage of ubiquitin fusion proteins in this system is relatively inefficient, although our ability to detect the cleaved, free Reaper protein may have been negatively impacted by its ability to induce cell death. We also observed a larger band of unknown origin in the MRE/control virus, which suggests that the ubiquitin expressed from these constructs may be utilized as a substrate by cellular ubiquitin ligases. We cannot conclude with certainty whether the expression of ubiquitin from the virus has any effect on virus structure or replication. However, the results of the replication curve experiments indicated that the ubiquitin fusion strategy did not result in any significant replication defects. In BHK-21 cells, MRE/control replicated to the highest level, with MRE/rprORF and MRE/rpr replicating to modestly lower levels before decreasing after 3 dpi. Although SINV infection alone causes apoptosis in BHK-21 cells [
40,
41], Reaper expression also induces apoptosis in mammalian cells, at least partially due to global suppression of protein synthesis [
42], which might explain why the Reaper-expressing viruses replicated to lower titers than MRE/control in these cells.
In the cumulative replication curve in C6/36 cells, the differences between MRE/control and the Reaper-expressing viruses were more pronounced. This was expected since SINV does not itself cause very much death in these cells, instead establishing a persistent infection [
23]. In this experiment, the titers of MRE/rprORF and MRE/rpr closely matched each other, suggesting that neither the location of the
reaper insert nor the expression of ubiquitin affected replication. In the C6/36 noncumulative replication curve, the titers of MRE/control, MRE/rprORF, and MRE/rpr closely aligned until 3 dpi, after which it appears that enough Reaper protein had accumulated to cause significant cell death and a decrease in titer was seen. MRE/rprORF titers decreased after 3 dpi to a greater extent than those of MRE/rpr, and these two replication curves were found to be significantly different. The reason for this is unknown but it is consistent with the higher level of caspase activity in MRE/rprORF-infected midgut compared to MRE/rpr. Thus, instability of the
reaper insert in MRE/rpr may have decreased its effectiveness in causing cell death both in cultured cells and in mosquito midgut. It is important to note that C6/36 cells have an impaired RNAi response and therefore results in these cells may not completely reflect the in vivo situation [
43].
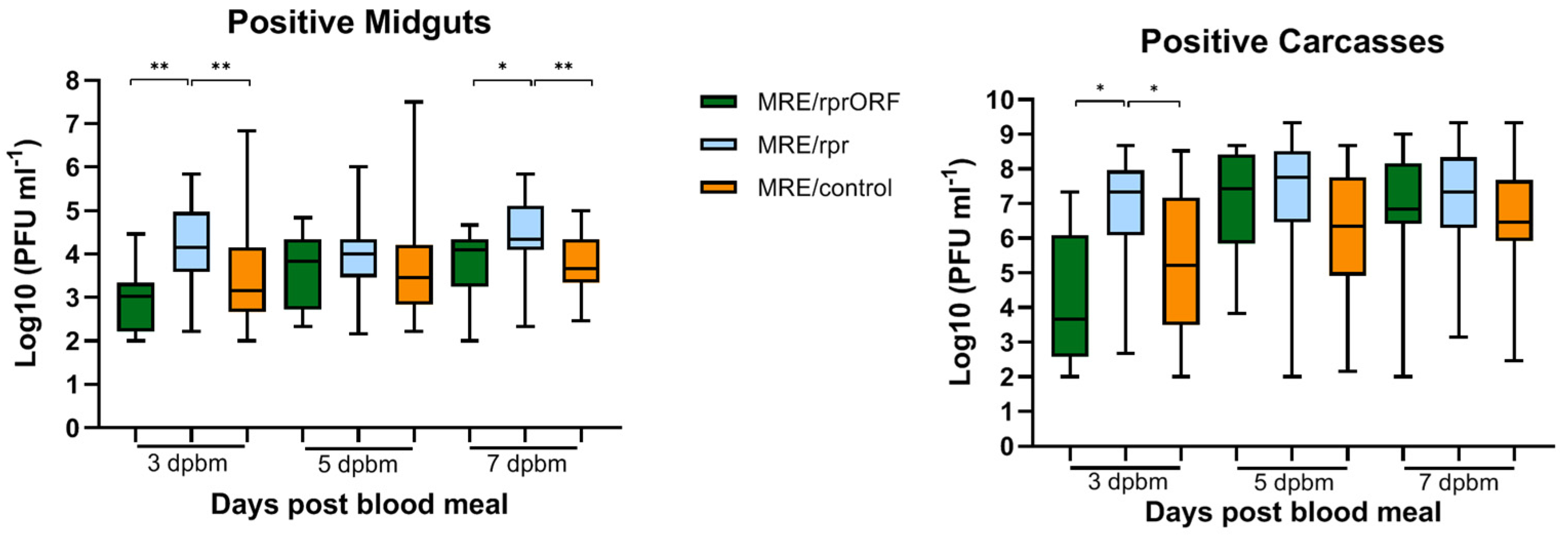
In mosquitoes, MRE/rprORF had a decreased ability to establish both midgut and carcass infection compared to MRE/rpr. At all time points tested, MRE/rprORF caused a lower infection prevalence compared to both MRE/control and MRE/rpr, while the prevalence of MRE/rpr infection did not significantly differ from MRE/control at any of the time points. This latter result differed from our previous study, which found that the percentage of mosquitoes infected with MRE/rpr was lower than the control virus used in that study at early time points in both the midgut and the carcass [
19]. Although we observed that MRE/rpr infection prevalence was lower than MRE/control at 3 dpi (65% vs. 81%), this difference did not reach statistical significance. Perhaps additional replicates would accentuate this difference. It is also possible that this difference could be due to the fact that different control viruses were used in the two studies. Regardless, the most important result for the purposes of this study was that MRE/rprORF showed significantly less ability to establish infection in the midgut and disseminate compared to MRE/rpr. We conclude that this is likely due to selective pressure to retain the
reaper insert in the structural ORF, since deletions within the
reaper insert would theoretically have a 2 out of 3 chance of altering the reading frame and eliminating expression of the downstream envelope proteins, resulting in defective virus. In contrast, any deletions within the
reaper insert of MRE/rpr virus would not be expected to impact virus viability. Indeed, our deep sequencing data demonstrated that the
reaper insert was more stably retained in MRE/rprORF than in MRE/rpr [
19], and all of our results are consistent with this being the case.
The results of our study also differed from previous published results [
19] when we examined the virus titers in mosquitoes that did become infected. We found that when MRE/rprORF was able to establish infection in the midgut, the titers were similar to MRE/control and this pattern continued in the carcass. It is possible that if the virus is able to accumulate to a certain level in midgut cells of some mosquitoes, apoptosis may no longer be able to effectively limit viral replication and spread. Another unexpected result was the observation that the titers of mosquitoes that became infected with MRE/rpr were higher compared to MRE/rprORF and MRE/control in the midguts at 3 and 7 days PBM and at 3 days PBM in the carcass. Deep sequencing data from our 2015 study indicated that most MRE/rpr-infected mosquitoes contain a high frequency of viruses with deletions in
reaper [
19], so this would be expected to allow it to replicate to higher levels compared to MRE/rprORF, but it is unclear why the titers were higher than MRE/control. In any case, none of the three viruses appeared to have a large replication advantage over the others in infected mosquitoes.
Deep sequencing of MRE/rprORF-infected mosquitoes demonstrated that
reaper was much more stably maintained during infection compared to MRE/rpr [
19], with no evidence of any deletions accumulating at >1% frequency in MRE/rprORF. However, in two mosquitoes, a point mutation in Reaper, T8937C, became dominant in the population. This point mutation changes an amino acid (L40) in the GH3 domain of Reaper. There is published evidence indicating that the Reaper GH3 domain plays a supportive role in inducing apoptosis, although it appears to be less important than the IBM [
38]. While position L40 is located within the GH3 domain, it is not well conserved in the GH3 domains of Reaper-like proteins from Drosophilids, mosquitoes, or lepidoptera [
38,
39], so the effect of this mutation on Reaper-induced apoptosis is unclear. Nonetheless, the observation that this mutation became highly dominant in two mosquitoes makes it tempting to speculate that the T8937C mutation may confer a selective advantage compared to MRE/rprORF. However, the T8937C mutation was also detected in all other sequenced samples (at 1.4% in one mosquito and at <1% in the MRE/rprORF stock and the other seven mosquitoes;
Data Set S1), so if it provides a selective advantage, it is unclear why it did not predominate in more than two of the ten mosquitoes. Overall, despite this single point mutation becoming dominant in two mosquitoes, the sequencing results indicate remarkably little change in the
reaper insert of MRE/rprORF during infection, especially when compared to the extensive loss of
reaper that was observed in MRE/rpr.
Our results demonstrate that expression of the pro-apoptotic Reaper protein negatively affects midgut infection and dissemination by SINV. While we cannot completely rule out other explanations for this effect, it is most likely due to induction of apoptosis by Reaper. This conclusion is supported by our previous work with MRE/rpr (showing increased midgut TUNEL staining) as well as several other studies [
18,
19,
20,
21]. However, there have been studies which suggest a seemingly opposite conclusion. In one study where expression of AeIAP1, which inhibits apoptosis, was knocked down in mosquitoes, both midgut infection and dissemination increased [
22]. However, AeIAP1 knockdown caused a high mortality rate and extensive pathology of the midgut tissue was observed. The systemic induction of apoptosis thus likely greatly decreased the structural integrity of the midgut, which may have allowed viral passage through the tissue. In the present study as well as in O’Neill et al. [
19], the pro-apoptotic gene was inserted into the viral genome, ensuring that apoptosis was stimulated only in infected cells. Additionally, two studies have shown that knockdown of the caspase AeDronc has the opposite effect than would be expected if apoptosis was strictly antiviral, as it reduces infection with SINV or DENV-2 [
22,
44]. While Eng et al [
44] provided evidence that AeDronc may play a role in autophagy, a process that may be important for arbovirus infection, another possible explanation for this result is that caspases are involved in remodeling of midgut basal lamina [
45], which appears to be necessary for viral midgut escape [
46]. Thus, either a low basal level of midgut caspase activity or extensive, widespread stimulation of apoptosis may be advantageous for a virus, while stimulation of apoptosis specifically in infected cells has a negative effect.
There are several limitations of our study which could be explored and improved upon in future studies. In this study, we only looked at the effect of Reaper expression on SINV in Ae. aegypti. This should be explored in other virus/vector combinations to determine if the results of this study are generalizable to other situations. Additionally, while less than 50% of mosquitoes exposed to MRE/rprORF developed disseminated infection by 7 days PBM, we did see this percentage increase from 3 to 7 days PBM. It is possible that at further time points the percentage of infection of MRE/rprORF would become equivalent to the control. The durability of this effect should be further explored in the future.
Overall, this study provides additional evidence that expression of the pro-apoptotic protein Reaper has a negative effect on the ability of SINV to establish infection in the midgut and disseminate to the carcass. Expressing Reaper by inserting the gene into the structural ORF caused a more robust and durable reduction in infection prevalence than expression via the duplicated subgenomic promoter, with the differences in both midgut and carcass infection prevalence between MRE/rprORF and MRE/control being significant at all of the time points tested. These results provide additional evidence that the apoptotic pathway is antiviral in mosquitoes and possibly could be exploited to prevent transmission of arboviruses. Additionally, this study is consistent with previous findings that inserting genes into a viral ORF is a useful strategy for expression of genes that are subject to negative selection.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}