
Occurrence of Equine Foamy Virus Infection in Horses from Poland
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Horse Samples
2.2. DNA Preparation
2.3. EFVeca DNA Detection
2.4. EFV Sequence Analysis
2.5. EFVeca Gag Amplification and Selection of a Sequence for Recombinant Antigen Preparation
2.6. Development of Recombinant Gag Protein of EFVeca
2.7. Immunoblotting for the Detection of the Horse Sera Reactivity to EFVeca Recombinant Antigen
2.8. EFVeca Gag Specific ELISA Test
2.9. Detection of Antibodies Specific for EHV-1, EAV and EIV
2.10. Statistical Analysis
3. Results
3.1. Detection of EFVeca DNA in Peripheral Blood Leukocytes of Horses
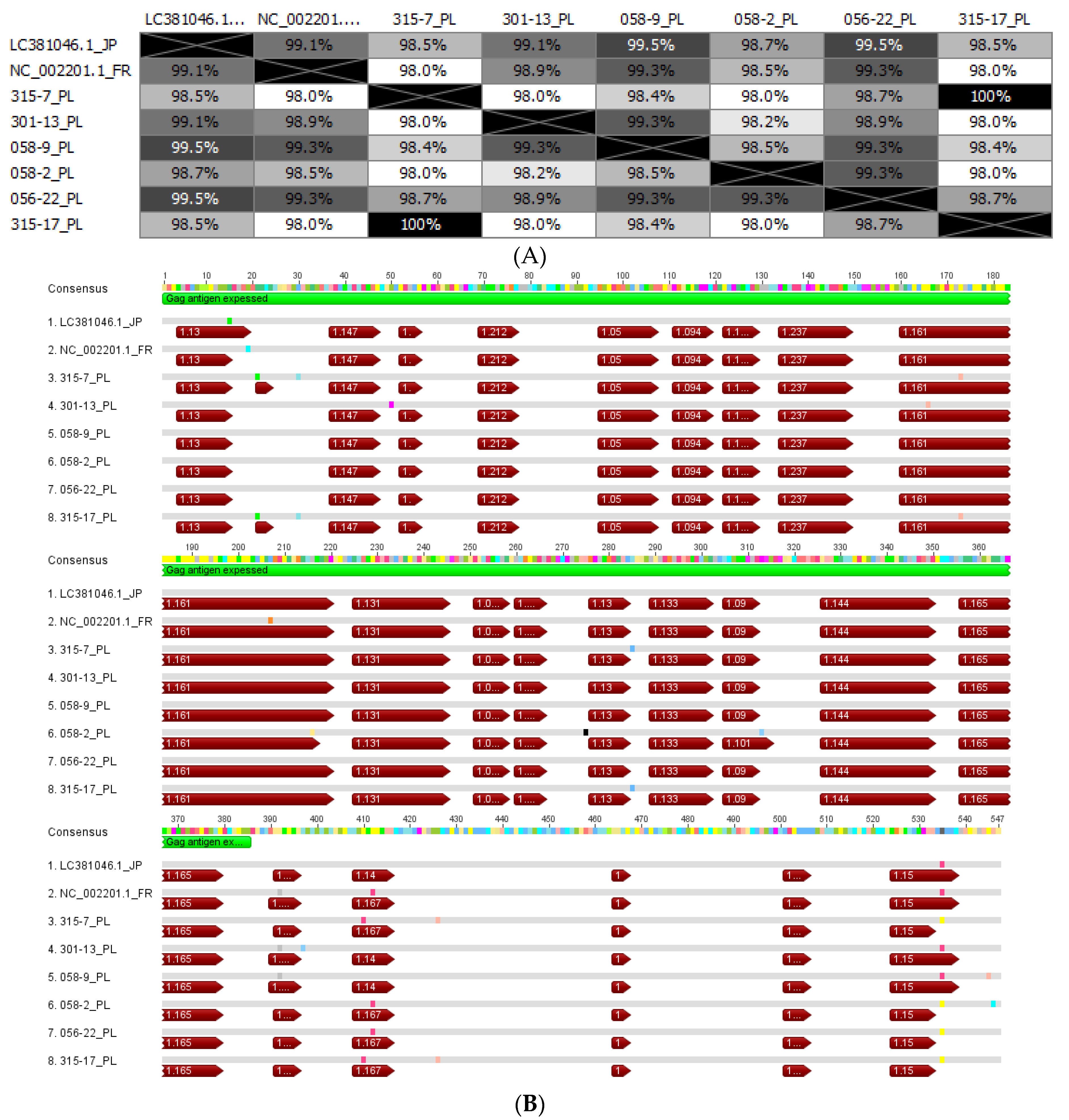
3.2. Analysis of EFVeca Gag Sequences Derived from Horses
3.3. Development and Optimization of EFVeca Specific ELISA
3.4. Serological Testing of Horse Samples Using a Newly-Developed EFVeca ELISA
3.5. Co-Infections of EFVeca and Other Common Equine Viruses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, A.S.; Bodem, J.; Buseyne, F.; Gessain, A.; Johnson, W.; Kuhn, J.H.; Kuzmak, J.; Lindemann, D.; Linial, M.L.; Löchelt, M.; et al. Spumaretroviruses: Updated taxonomy and nomenclature. Virology 2018, 516, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Kehl, T.; Tan, J.; Materniak, M. Non-Simian Foamy Viruses: Molecular Virology, Tropism and Prevalence and Zoonotic/Interspecies Transmission. Viruses 2013, 5, 2169–2209. [Google Scholar] [CrossRef]
- Materniak, M.; Hechler, T.; Lochelt, M.; Kuzmak, J.; Löchelt, M.; Kuźmak, J. Similar Patterns of Infection with Bovine Foamy Virus in Experimentally Inoculated Calves and Sheep. J. Virol. 2013, 87, 3516–3525. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; Picker, L.J.; Axthelm, M.K.; Linial, M.L. Expanded Tissue Targets for Foamy Virus Replication with Simian Immunodeficiency Virus-Induced Immunosuppression. J. Virol. 2006, 80, 663–670. [Google Scholar] [CrossRef]
- Johnson, R.H.; de la Rosa, J.; Abher, I.; Kertayadnya, I.G.; Entwistle, K.W.; Fordyce, G.; Holroyd, R.G. Epidemiological studies of bovine spumavirus. Vet. Microbiol. 1988, 16, 25–33. [Google Scholar] [CrossRef]
- Kertayadnya, I.G.; Johnson, R.H.; Abher, I.; Burgess, G.W. Detection of immunological tolerance to bovine spumavirus (BSV) with evidence for salivary excretion and spread of BSV from the tolerant animal. Vet. Microbiol. 1988, 16, 35–39. [Google Scholar] [CrossRef]
- Flügel, R.M. Spumaviruses: A group of complex retroviruses. J. Acquir. Immune Defic. Syndr. 1991, 4, 739–750. [Google Scholar] [PubMed]
- Malmquisl, W.A.; Van Der Maaten, M.J.; Boothe, A.D. Isolation, Immunodiffusion, Immunofluorescence, and Electron Microscopy of a Syncytial Virus of Lymphosarcomatous and Apparently Normal Cattle. Cancer Res. 1969, 29, 188–200. [Google Scholar]
- Materniak-Kornas, M.; Tan, J.; Heit-Mondrzyk, A.; Hotz-Wagenblatt, A.; Löchelt, M. Bovine Foamy Virus: Shared and Unique Molecular Features In Vitro and In Vivo. Viruses 2019, 11, 1084. [Google Scholar] [CrossRef]
- Calattini, S.; Nerrienet, E.; Mauclère, P.; Georges-Courbot, M.-C.; Saïb, A.; Gessain, A. Natural simian foamy virus infection in wild-caught gorillas, mandrills and drills from Cameroon and Gabon. J. Gen. Virol. 2004, 85, 3313–3317. [Google Scholar] [CrossRef]
- Liu, W.; Worobey, M.; Li, Y.; Keele, B.F.; Bibollet-Ruche, F.; Guo, Y.; Goepfert, P.A.; Santiago, M.L.; Ndjango, J.-B.N.; Neel, C.; et al. Molecular ecology and natural history of simian foamy virus infection in wild-living chimpanzees. PLoS Pathog. 2008, 4, e1000097. [Google Scholar]
- Santos, A.F.; Cavalcante, L.T.F.; Muniz, C.P.; Switzer, W.M.; Soares, M.A. Simian Foamy Viruses in Central and South America: A New World of Discovery. Viruses 2019, 11, 967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romen, F.; Backes, P.; Materniak, M.; Sting, R.; Vahlenkamp, T.W.; Riebe, R.; Pawlita, M.; Kuzmak, J.; Löchelt, M. Serological detection systems for identification of cows shedding bovine foamy virus via milk. Virology 2007, 364, 123–131. [Google Scholar] [PubMed]
- Romen, F.; Pawlita, M.; Sehr, P.; Bachmann, S.; Schröder, J.; Lutz, H.; Löchelt, M. Antibodies against Gag are diagnostic markers for feline foamy virus infections while Env and Bet reactivity is undetectable in a substantial fraction of infected cats. Virology 2006, 345, 502–508. [Google Scholar] [PubMed]
- Tobaly-Tapiero, J.; Bittoun, P.; Neves, M.; Guillemin, M.C.; Lecellier, C.H.; Puvion-Dutilleul, F.; Gicquel, B.; Zientara, S.; Giron, M.L.; de Thé, H.; et al. Isolation and characterization of an equine foamy virus. J. Virol. 2000, 74, 4064–4073. [Google Scholar] [CrossRef]
- Kirisawa, R.; Toishi, Y.; Hashimoto, H.; Tsunoda, N. Isolation of an equine foamy virus and sero-epidemiology of the viral infection in horses in Japan. Viruses 2019, 11, 613. [Google Scholar] [CrossRef]
- Jacobs, R.M.; Pollari, F.L.; McNab, W.B.; Jefferson, B. A serological survey of bovine syncytial virus in Ontario: Associations with bovine leukemia and immunodeficiency-like viruses, production records, and management practices. Can. J. Vet. Res. 1995, 59, 271–278. [Google Scholar]
- Gessain, A.; Montange, T.; Betsem, E.; Bilounga, C.; Njouom, N.R.; Buseyne, F. Case-control study of the immune status of humans infected with zoonotic gorilla simian foamy viruses. J. Infect. Dis. 2020, 221, 1724–1733. [Google Scholar]
- Buseyne, F.; Betsem, E.; Montange, T.; Njouom, R.; Bilounga Ndongo, C.; Hermine, O.; Gessain, A. Clinical Signs and Blood Test Results Among Humans Infected With Zoonotic Simian Foamy Virus: A Case-Control Study. J. Infect. Dis. 2018, 218, 144–151. [Google Scholar] [CrossRef]
- Amborski, G.F.; Lo, J.L.; Seger, C.L. Serological detection of multiple retroviral infections in cattle: Bovine leukemia virus, bovine syncytial virus and bovine visna virus. Vet. Microbiol. 1989, 20, 247–253. [Google Scholar]
- Jacobs, R.M.; Smith, H.E.; Gregory, B.; Valli, V.E.; Whetstone, C.A. Detection of multiple retroviral infections in cattle and cross-reactivity of bovine immunodeficiency-like virus and human immunodeficiency virus type 1 proteins using bovine and human sera in a western blot assay. Can. J. Vet. Res. 1992, 56, 353–359. [Google Scholar] [PubMed]
- Hooks, J.J.; Gibbs, C.J. The foamy viruses. Bacteriol. Rev. 1975, 39, 169–185. [Google Scholar] [PubMed]
- Zenger, E.; Brown, W.C.; Song, W.; Wolf, A.M.; Pedersen, N.C.; Longnecker, M.; Li, J.; Collisson, E.W. Evaluation of cofactor effect of feline syncytium-forming virus on feline immunodeficiency virus infection. Am. J. Vet. Res. 1993, 54, 713–718. [Google Scholar] [PubMed]
- Hood, S.; Mitchell, J.L.; Sethi, M.; Almond, N.M.; Cutler, K.L.; Rose, N.J. Horizontal acquisition and a broad biodistribution typify simian foamy virus infection in a cohort of Macaca fascicularis. Virol. J. 2013, 10, 326. [Google Scholar]
- Jones-Engel, L.; Steinkraus, K.A.; Murray, S.M.; Engel, G.A.; Grant, R.; Aggimarangsee, N.; Lee, B.P.Y.-H.; May, C.; Schillaci, M.A.; Somgird, C.; et al. Sensitive assays for simian foamy viruses reveal a high prevalence of infection in commensal, free-ranging Asian monkeys. J. Virol. 2007, 81, 7330–7337. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, R. DNA sequencing with chain-terminating. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef]
- Parker, J.M.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar]
- Materniak, M.; Bicka, L.; Kuźmak, J. Isolation and partial characterization of bovine foamy virus from Polish cattle. Pol. J. Vet. Sci. 2006, 9, 207–211. [Google Scholar] [PubMed]
- Cullinane, A. Equine influenza (infection with equine influenza virus). In Manual of Diagnositc Tests and Vaccines for Terrestial Animals; World Organisation for Animal Health: Paris, France, 2022. [Google Scholar]
- Elton, D.; Bryant, N. Equine rhinopneumonitis (equine herpesvirus-1 and -4)No Title. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2022. [Google Scholar]
- Timoney, P. Equine viral arteritis (infection with equine arteritis virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2022. [Google Scholar]
- Branscum, A.J.; Gardner, I.A.; Johnson, W.O. Estimation of diagnostic-test sensitivity and specificity through Bayesian modeling. Prev. Vet. Med. 2005, 68, 145–163. [Google Scholar] [PubMed]
- Uriu, K.; Kosugi, Y.; Ito, J.; Sato, K. The Battle between Retroviruses and APOBEC3 Genes: Its Past and Present. Viruses 2021, 13, 124. [Google Scholar] [CrossRef] [PubMed]
- Müllers, E. The foamy virus Gag proteins: What makes them different? Viruses 2013, 5, 1023–1041. [Google Scholar]
- Materniak-Kornas, M.; Osiński, Z.; Rudzki, M.; Kuźmak, J. Development of a recombinant protein-based ELISA for detection of antibodies against bovine foamy virus. J. Vet. Res. 2017, 61, 247–252. [Google Scholar] [PubMed]
- Leendertz, S.A.J.; Junglen, S.; Hedemann, C.; Goffe, A.; Calvignac, S.; Boesch, C.; Leendertz, F.H. High prevalence, coinfection rate, and genetic diversity of retroviruses in wild red colobus monkeys (Piliocolobus badius badius) in Tai National Park, Cote d’Ivoire. J. Virol. 2010, 84, 7427–7436. [Google Scholar] [CrossRef]
- Mouinga-Ondémé, A.; Betsem, E.; Caron, M.; Makuwa, M.; Sallé, B.; Renault, N.; Saib, A.; Telfer, P.; Marx, P.; Gessain, A.; et al. Two distinct variants of simian foamy virus in naturally infected mandrills (Mandrillus sphinx) and cross-species transmission to humans. Retrovirology 2010, 7, 105. [Google Scholar]
- Muniz, C.P.; Troncoso, L.L.; Moreira, M.A.; Soares, E.A.; Pissinatti, A.; Bonvicino, C.R.; Seuánez, H.N.; Sharma, B.; Jia, H.; Shankar, A.; et al. Identification and Characterization of Highly Divergent Simian Foamy Viruses in a Wide Range of New World Primates from Brazil. PLoS ONE 2013, 8, e67568. [Google Scholar]
- Nakamura, K.; Miyazawa, T.; Ikeda, Y.; Sato, E.; Nishimura, Y. Contrastive Prevalence of Feline Retrovirus Infections between Northern and. Vet. Med. Sci. 2000, 62, 921–923. [Google Scholar] [CrossRef]
- Winkler, I.G.; Löchelt, M.; Flower, R.L.P. Epidemiology of feline foamy virus and feline immunodeficiency virus infections in domestic and feral cats: A seroepidemiological study. J. Clin. Microbiol. 1999, 37, 2848–2851. [Google Scholar]
- Hahn, H.; Baunach, G.; Bräutigam, S.; Mergia, A.; Neumann-Haefelin, D.; Daniel, M.D.; McClure, M.O.; Rethwilm, A. Reactivity of primate sera to foamy virus Gag and Bet proteins. J. Gen. Virol. 1994, 75, 2635–2644. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.I.; Shanmugam, V.; Bhullar, V.B.; Beer, B.E.; Vallet, D.; Gautier-Hion, A.; Wolfe, N.D.; Karesh, W.B.; Kilbourn, A.M.; Tooze, Z.; et al. Screening for simian foamy virus infection by using a combined antigen Western blot assay: Evidence for a wide distribution among Old World primates and identification of four new divergent viruses. Virology 2003, 309, 248–257. [Google Scholar] [CrossRef]
- Winkler, I.G.; Löchelt, M.; Levesque, J.P.; Bodem, J.; Flügel, R.M.; Flower, R.L.P. A rapid streptavidin-capture ELISA specific for the detection of antibodies to feline foamy virus. J. Immunol. Methods 1997, 207, 69–77. [Google Scholar] [CrossRef]
- Rua, R.; Betsem, E.; Gessain, A. Viral latency in blood and saliva of simian foamy virus-infected humans. PLoS ONE 2013, 8, e77072. [Google Scholar] [CrossRef]
- Calattini, S.; Nerrienet, E.; Mauclère, P.; Georges-Courbot, M.-C.; Saib, A.; Gessain, A. Detection and molecular characterization of foamy viruses in Central African chimpanzees of the Pan troglodytes troglodytes and Pan troglodytes vellerosus subspecies. J. Med. Primatol. 2006, 35, 59–66. [Google Scholar] [PubMed]
- Hechler, T.; Materniak, M.; Kehl, T.; Kuzmak, J.; Lochelt, M. Complete Genome Sequences of Two Novel European Clade Bovine Foamy Viruses from Germany and Poland. J. Virol. 2012, 86, 10905–10906. [Google Scholar]
- Kechejian, S.R.; Dannemiller, N.; Kraberger, S.; Ledesma-Feliciano, C.; Malmberg, J.; Roelke Parker, M.; Cunningham, M.; McBride, R.; Riley, S.P.D.; Vickers, W.T.; et al. Feline Foamy Virus is Highly Prevalent in Free-Ranging Puma concolor from Colorado, Florida and Southern California. Viruses 2019, 11, 359. [Google Scholar]
- Bleiholder, A.; Mühle, M.; Hechler, T.; Bevins, S.; vandeWoude, S.; Denner, J.; Löchelt, M. Pattern of seroreactivity against feline foamy virus proteins in domestic cats from Germany. Vet. Immunol. Immunopathol. 2011, 143, 292–300. [Google Scholar] [CrossRef]
- Materniak-Kornas, M.; Frymus, T.; Löchelt, M.; Kuźmak, J. Seroprevalence of feline foamy virus in domestic cats in Poland. J. Vet. Res. 2021, 65, 407–413. [Google Scholar]
- Dannemiller, N.G.; Kechejian, S.; Kraberger, S.; Logan, K.; Alldredge, M.; Crooks, K.R.; VandeWoude, S.; Carver, S. Diagnostic Uncertainty and the Epidemiology of Feline Foamy Virus in Pumas (Puma concolor). Sci. Rep. 2020, 10, 1587. [Google Scholar]
- Castaneda, C.; Juras, R.; Khanshour, A.; Randlaht, I.; Wallner, B.; Rigler, D.; Lindgren, G.; Raudsepp, T.; Gus Cothran, E. Population genetic analysis of the Estonian native horse suggests diverse and distinct genetics, ancient origin and contribution from unique patrilines. Genes 2019, 10, 629. [Google Scholar] [CrossRef] [PubMed]
- Hooks, J.J.; Burns, W.; Hayashi, K.; Geis, S.; Notkins, A.L. Viral spread in the presence of neutralizing antibody: Mechanisms of persistence in foamy virus infection. Infect. Immun. 1976, 14, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Ayouba, A.; Duval, L.; Liégeois, F.; Ngin, S.; Ahuka-Mundeke, S.; Switzer, W.M.; Delaporte, E.; Ariey, F.; Peeters, M.; Nerrienet, E. Nonhuman primate retroviruses from Cambodia: High simian foamy virus prevalence, identification of divergent STLV-1 strains and no evidence of SIV infection. Infect. Genet. Evol. 2013, 18, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Feliciano, C.; Troyer, R.M.; Zheng, X.; Miller, C.; Cianciolo, R.; Bordicchia, M.; Dannemiller, N.; Gagne, R.; Beatty, J.; Quimby, J.; et al. Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease. Viruses 2019, 11, 662. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.J.; Moura, L.; Miller, T.; Thurk, J.; Perreault, N.; Weil, A.; Maggio, R.; Lucas, H.; Breitschwerdt, E. Feline immunodeficiency virus, feline leukemia virus and Bartonella species in stray cats on St Kitts, West Indies. J. Feline Med. Surg. 2010, 12, 447–450. [Google Scholar] [CrossRef]
- Olech, M.; Valas, S.; Kuźmak, J. Epidemiological survey in single-species flocks from Poland reveals expanded genetic and antigenic diversity of small ruminant lentiviruses. PLoS ONE 2018, 13, e0193892. [Google Scholar]
- Rola-Łuszczak, M.; Sakhawat, A.; Pluta, A.; Ryło, A.; Bomba, A.; Bibi, N.; Kuźmak, J. Molecular Characterization of the env Gene of Bovine Leukemia Virus in Cattle from Pakistan with NGS-Based Evidence of Virus Heterogeneity. Pathogens 2021, 10, 910. [Google Scholar] [CrossRef]
- Bandecchi, P.; Matteucci, D.; Baldinotti, F.; Guidi, G.; Abramo, F.; Tozzini, F.; Bendinelli, M. Prevalence of feline immunodeficiency virus and other retroviral infections in sick cats in Italy. Vet. Immunol. Immunopathol. 1992, 31, 337–345. [Google Scholar] [CrossRef]
- Choudhary, A.; Galvin, T.A.; Williams, D.K.; Beren, J.; Bryant, M.A.; Khan, A.S. Influence of naturally occurring simian foamy viruses (SFVs) on SIV disease progression in the rhesus macaque (Macaca mulatta) model. Viruses 2013, 5, 1414–1430. [Google Scholar] [CrossRef]
- Switzer, W.M.; Tang, S.; Zheng, H.; Shankar, A.; Sprinkle, P.S.; Sullivan, V.; Granade, T.C.; Heneine, W. Dual Simian Foamy Virus/Human Immunodeficiency Virus Type 1 Infections in Persons from Côte d’Ivoire. PLoS ONE 2016, 11, e0157709. [Google Scholar]
- Switzer, W.M.; Garcia, A.D.; Yang, C.; Wright, A.; Kalish, M.L.; Folks, T.M.; Heneine, W. Coinfection with HIV-1 and simian foamy virus in West Central Africans. J. Infect. Dis. 2008, 197, 1389–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alais, S.; Pasquier, A.; Jegado, B.; Journo, C.; Rua, R.; Gessain, A.; Tobaly-Tapiero, J.; Lacoste, R.; Turpin, J.; Mahieux, R. STLV-1 co-infection is correlated with an increased SFV proviral load in the peripheral blood of SFV/STLV-1 naturally infected non-human primates. PLoS Negl. Trop. Dis. 2018, 12, e0006812. [Google Scholar] [CrossRef] [PubMed]
- Rola, J.; Larska, M.; Rola, J.G.; Belák, S.; Autorino, G.L. Epizotiology and phylogeny of equine arteritis virus in hucul horses. Vet. Microbiol. 2011, 148, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, A.J.; Howell, P.G.; Hedges, J.F.; Bosman, A.M.; Balasuriya, U.B.R.; McCollum, W.H.; Timoney, P.J.; MacLachlan, N.J. Lateral transmission of equine arteritis virus among Lipizzaner stallions in South Africa. Equine Vet. J. 2003, 35, 596–600. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Prior Information | Posterior Information |
|---|---|---|
| Values of Apparent Parameters (Median and the 95% CI) | Values of True Parameters (Median and the 95% CI) | |
| ELISA Se | 1.0 | 0.9731 (0.9172–0.9951) |
| ELISA Sp | 0.935 | 0.9109 (0.8050–0.9743). |
| WB Se | 0.974 (0.935–0.995) | 0.9877 (0.9803–0.9991) |
| WB Sp | 0.944 (0.880–0.983) | 0.9953 (0.9903–0.9998) |
| Samples Origin | Number of Tested Horses | ELISA Results | PCR Results | ||
|---|---|---|---|---|---|
| Negative | Positive | Negative | Positive | ||
| Hucul horses | 58 | 16 (27.6%) | 42 (72.4%) | 31 (55.4%) | 27 (46.6%) |
| The Konik | 25 | 21 (84%) | 4 (16%) | 25 (100%) | 0 (0%) |
| Livestock horses | 66 | 51 (77.3%) | 15 (22.7%) | 57 (86.4%) | 9 (13.6%) |
| Saddle horses | 17 | 6 (35.3%) | 11 (64.7%) | 13 (76.5%) | 4 (23.5%) |
| Purebred horses | 82 | 19 (23.2%) | 63 (76.8%) | 37 (45.1%) | 45 (54.9%) |
| Total | 248 | 113 (45.6%) | 135 (55.6%) | 163 (65.7%) | 85 (34.3%) |
| PCR (+) | PCR (−) | Total | |
|---|---|---|---|
| ELISA (+) | 76 | 59 | 135 |
| ELISA (−) | 9 | 104 | 113 |
| Total | 85 | 163 | 248 |
| EIV A1 | EIV A2 | EAV | EHV-1 | |||||
|---|---|---|---|---|---|---|---|---|
| + | − | + | − | + | − | + | − | |
| EFVeca + | 54 | 6 | 54 | 6 | 52 | 8 | 59 | 1 |
| EFVeca − | 19 | 2 | 18 | 3 | 10 | 11 | 19 | 2 |
| Total | 81 | 81 | 81 | 81 | ||||
| Chi2 Test | chi2 = 0.004 p = 0.9498 | chi2 = 0.289 p = 0.5907 | chi2 = 13.21 p = 0.0003 * | chi2 = 2.693 p = 0.1008 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Materniak-Kornas, M.; Rożek, W.; Rola, J.; Osiński, Z.; Löchelt, M.; Kuźmak, J. Occurrence of Equine Foamy Virus Infection in Horses from Poland. Viruses 2022, 14, 1973. https://doi.org/10.3390/v14091973
Materniak-Kornas M, Rożek W, Rola J, Osiński Z, Löchelt M, Kuźmak J. Occurrence of Equine Foamy Virus Infection in Horses from Poland. Viruses. 2022; 14(9):1973. https://doi.org/10.3390/v14091973
Chicago/Turabian StyleMaterniak-Kornas, Magdalena, Wojciech Rożek, Jerzy Rola, Zbigniew Osiński, Martin Löchelt, and Jacek Kuźmak. 2022. "Occurrence of Equine Foamy Virus Infection in Horses from Poland" Viruses 14, no. 9: 1973. https://doi.org/10.3390/v14091973