
Pro-Inflammatory Profile of Children Exposed to Maternal Chikungunya Virus Infection during the Intrauterine Period: A One-Year Follow-Up Study

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Data Collection
2.2. Patients
2.3. Blood Sampling
2.4. Multiplex Immunoassay
2.5. Detection of Anti-CHIKV IgM and IgG Antibodies
2.6. Statistical Analysis
3. Results
3.1. Study Population Characteristics
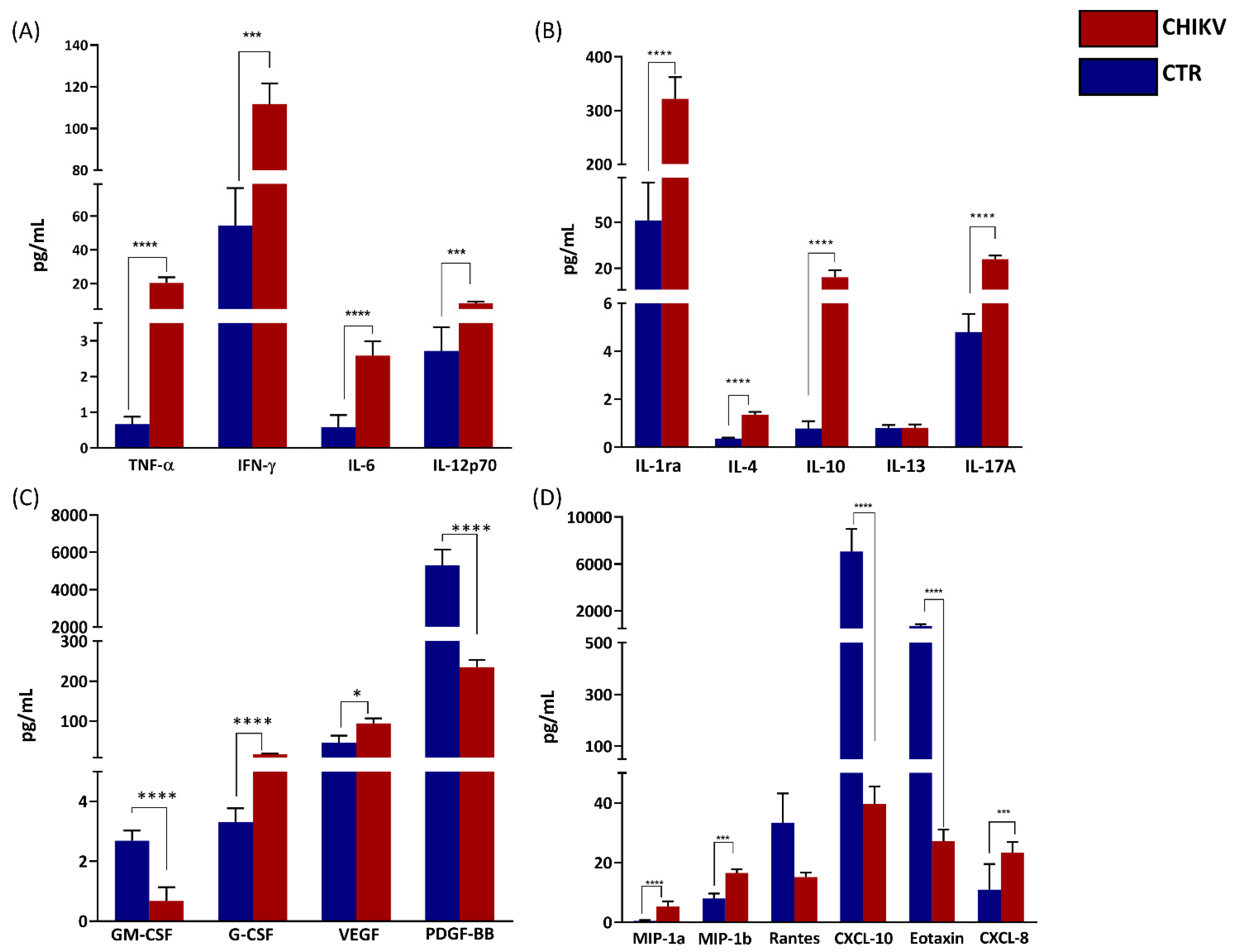
3.2. Analysis of Circulant Inflammatory Mediators
3.3. Principal Component Analysis (PCA) Shows the Relationship between Immune Mediators between Groups and According to Age and Gender
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global Expansion and Redistribution of Aedes-Borne Virus Transmission Risk with Climate Change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef]
- Kendall, C.; Khalid, H.; Müller, M.; Banda, D.H.; Kohl, A.; Merits, A.; Stonehouse, N.J.; Tuplin, A. Structural and Phenotypic Analysis of Chikungunya Virus RNA Replication Elements. Nucleic. Acids. Res. 2019, 47, 9296–9312. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.R.T.; Faria, N.R.; de Vasconcelos, J.M.; Golding, N.; Kraemer, M.U.G.; de Oliveira, L.F.; do Socorro da Silva Azevedo, R.; da Silva, D.E.A.; da Silva, E.V.P.; da Silva, S.P.; et al. Emergence and Potential for Spread of Chikungunya Virus in Brazil. BMC Med. 2015, 13, 102. [Google Scholar] [CrossRef]
- Gutiérrez, L.A. PAHO/WHO Data-Weekly Report | PAHO/WHO. Available online: https://www3.paho.org/data/index.php/en/mnu-topics/chikv-en/550-chikv-weekly-en.html (accessed on 28 November 2021).
- Torres, J.R.; Falleiros-Arlant, L.H.; Dueñas, L.; Pleitez-Navarrete, J.; Salgado, D.M.; Castillo, J.B.-D. Congenital and Perinatal Complications of Chikungunya Fever: A Latin American Experience. Int. J. Infect. Dis. 2016, 51, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.R.R.; Cavalcanti, L.; Zuben, A.P.V.; Donalisio, M.R. Excess Mortality Related to Chikungunya Epidemics in the Context of Co-Circulation of Other Arboviruses in Brazil. PLoS Curr. 2017, PMC5731794. [Google Scholar] [CrossRef]
- Kim, A.S.; Zimmerman, O.; Fox, J.M.; Nelson, C.A.; Basore, K.; Zhang, R.; Durnell, L.; Desai, C.; Bullock, C.; Deem, S.L.; et al. An Evolutionary Insertion in the Mxra8 Receptor-Binding Site Confers Resistance to Alphavirus Infection and Pathogenesis. Cell Host Microbe 2020, 27, 428–440.e9. [Google Scholar] [CrossRef]
- Zhang, R.; Kim, A.S.; Fox, J.M.; Nair, S.; Basore, K.; Klimstra, W.B.; Rimkunas, R.; Fong, R.H.; Lin, H.; Poddar, S.; et al. Mxra8 Is a Receptor for Multiple Arthritogenic Alphaviruses. Nature 2018, 557, 570–574. [Google Scholar] [CrossRef]
- Santhosh, S.R.; Dash, P.K.; Parida, M.M.; Khan, M.; Tiwari, M.; Lakshmana Rao, P.V. Comparative Full Genome Analysis Revealed E1: A226V Shift in 2007 Indian Chikungunya Virus Isolates. Virus Res. 2008, 135, 36–41. [Google Scholar] [CrossRef]
- de Souza, T.; Ribeiro, E.; Corrêa, V.; Damasco, P.; Santos, C.; de Bruycker-Nogueira, F.; Chouin-Carneiro, T.; Faria, N.; Nunes, P.; Heringer, M.; et al. Following in the Footsteps of the Chikungunya Virus in Brazil: The First Autochthonous Cases in Amapá in 2014 and Its Emergence in Rio de Janeiro during 2016. Viruses 2018, 10, 623. [Google Scholar] [CrossRef]
- Manimunda, S.P.; Vijayachari, P.; Uppoor, R.; Sugunan, A.P.; Singh, S.S.; Rai, S.K.; Sudeep, A.B.; Muruganandam, N.; Chaitanya, I.K.; Guruprasad, D.R. Clinical Progression of Chikungunya Fever during Acute and Chronic Arthritic Stages and the Changes in Joint Morphology as Revealed by Imaging. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 392–399. [Google Scholar] [CrossRef]
- Nikitina, E.; Larionova, I.; Choinzonov, E.; Kzhyshkowska, J. Monocytes and Macrophages as Viral Targets and Reservoirs. Int. J. Mol. Sci. 2018, 19, 2821. [Google Scholar] [CrossRef] [PubMed]
- Chirathaworn, C.; Chansaenroj, J.; Poovorawan, Y. Cytokines and Chemokines in Chikungunya Virus Infection: Protection or Induction of Pathology. Pathogens 2020, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Contopoulos-Ioannidis, D.; Newman-Lindsay, S.; Chow, C.; LaBeaud, A.D. Mother-to-Child Transmission of Chikungunya Virus: A Systematic Review and Meta-Analysis. PLOS Negl. Trop. Dis. 2018, 12, e0006510. [Google Scholar] [CrossRef]
- Gérardin, P.; Barau, G.; Michault, A.; Bintner, M.; Randrianaivo, H.; Choker, G.; Lenglet, Y.; Touret, Y.; Bouveret, A.; Grivard, P.; et al. Multidisciplinary Prospective Study of Mother-to-Child Chikungunya Virus Infections on the Island of La Réunion. PLoS Med. 2008, 5, e60. [Google Scholar] [CrossRef] [PubMed]
- Gérardin, P.; Sampériz, S.; Ramful, D.; Boumahni, B.; Bintner, M.; Alessandri, J.-L.; Carbonnier, M.; Tiran-Rajaoefera, I.; Beullier, G.; Boya, I.; et al. Neurocognitive Outcome of Children Exposed to Perinatal Mother-to-Child Chikungunya Virus Infection: The CHIMERE Cohort Study on Reunion Island. PLoS Negl. Trop. Dis. 2014, 8, e2996. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, D.G.; Freddi, T.a.L.; Werner, H.; Lopes, F.P.P.L.; Moreira, M.E.L.; de Almeida Di Maio Ferreira, F.C.P.; de Andrade Lopes, J.M.; Rueda-Lopes, F.C.; da Cruz, L.C.H. Brain MR Imaging of Patients with Perinatal Chikungunya Virus Infection. AJNR Am. J. Neuroradiol 2020, 41, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Schepanski, S.; Buss, C.; Hanganu-Opatz, I.L.; Arck, P.C. Prenatal Immune and Endocrine Modulators of Offspring’s Brain Development and Cognitive Functions Later in Life. Front. Immunol. 2018, 9, 2186. [Google Scholar] [CrossRef]
- Vianna, R.A.d.O.; Lovero, K.L.; de Oliveira, S.A.; Fernandes, A.R.; Santos, T.C.S.D.; de Souza Lima, L.C.S.; Carvalho, F.R.; Quintans, M.D.S.; Bueno, A.C.; Torbey, A.F.M.; et al. Children Born to Mothers with Rash During Zika Virus Epidemic in Brazil: First 18 Months of Life. J. Trop. Pediatr. 2019, 65, 592–602. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya Virus in US Travelers Returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef]
- Costa, J.; Ferreira, E.C.; Santos, C. COVID-19, Chikungunya, Dengue and Zika Diseases: An Analytical Platform Based on MALDI-TOF MS, IR Spectroscopy and RT-QPCR for Accurate Diagnosis and Accelerate Epidemics Control. Microorganisms 2021, 9, 708. [Google Scholar] [CrossRef]
- Sevenoaks, T.; Wedderburn, C.J.; Donald, K.A.; Barnett, W.; Zar, H.J.; Stein, D.J.; Naudé, P.J.W. Association of Maternal and Infant Inflammation with Neurodevelopment in HIV-Exposed Uninfected Children in a South African Birth Cohort. Brain Behav. Immun. 2021, 91, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Zumpf, K.; Sprague, J.; Ciolino, J.; Wisner, K.L.; Clark, C.; Mancebo, M.C.; Eisen, J.L.; Rasmussen, S.A.; Boisseau, C.L. Impact of the Peripartum Period on the Longitudinal Course of Obsessive–Compulsive Disorder. Arch. Womens Ment. Health 2021, 24, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Macedo-da-Silva, J.; Rosa-Fernandes, L.; Barbosa, R.H.; Angeli, C.B.; Carvalho, F.R.; de Oliveira Vianna, R.A.; Carvalho, P.C.; Larsen, M.R.; Cardoso, C.A.; Palmisano, G. Serum Proteomics Reveals Alterations in Protease Activity, Axon Guidance, and Visual Phototransduction Pathways in Infants With In Utero Exposure to Zika Virus Without Congenital Zika Syndrome. Front. Cell. Infect. Microbiol. 2020, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Iosif, R.E.; Ekdahl, C.T.; Ahlenius, H.; Pronk, C.J.H.; Bonde, S.; Kokaia, Z.; Jacobsen, S.-E.W.; Lindvall, O. Tumor Necrosis Factor Receptor 1 Is a Negative Regulator of Progenitor Proliferation in Adult Hippocampal Neurogenesis. J. Neurosci. 2006, 26, 9703–9712. [Google Scholar] [CrossRef] [PubMed]
- von Ehrenstein, O.S.; Neta, G.I.; Andrews, W.; Goldenberg, R.; Goepfert, A.; Zhang, J. Child Intellectual Development in Relation to Cytokine Levels in Umbilical Cord Blood. Am. J. Epidemiol 2012, 175, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, T.; Roth, C.L. Inflammation Markers in Type 2 Diabetes and the Metabolic Syndrome in the Pediatric Population. Curr. Diab. Rep. 2018, 18, 131. [Google Scholar] [CrossRef]
- Semple, B.D.; Kossmann, T.; Morganti-Kossmann, M.C. Role of Chemokines in CNS Health and Pathology: A Focus on the CCL2/CCR2 and CXCL8/CXCR2 Networks. J. Cereb. Blood Flow Metab. 2010, 30, 459–473. [Google Scholar] [CrossRef]
- Kelland, E.E.; Gilmore, W.; Weiner, L.P.; Lund, B.T. The Dual Role of CXCL8 in Human CNS Stem Cell Function: Multipotent Neural Stem Cell Death and Oligodendrocyte Progenitor Cell Chemotaxis. Glia 2011, 59, 1864–1878. [Google Scholar] [CrossRef]
- Stuart, M.J.; Singhal, G.; Baune, B.T. Systematic Review of the Neurobiological Relevance of Chemokines to Psychiatric Disorders. Front. Cell Neurosci. 2015, 9, 357. [Google Scholar] [CrossRef]
- Lima, M.C.; de Mendonça, L.R.; Rezende, A.M.; Carrera, R.M.; Aníbal-Silva, C.E.; Demers, M.; D’Aiuto, L.; Wood, J.; Chowdari, K.V.; Griffiths, M.; et al. The Transcriptional and Protein Profile From Human Infected Neuroprogenitor Cells Is Strongly Correlated to Zika Virus Microcephaly Cytokines Phenotype Evidencing a Persistent Inflammation in the CNS. Front. Immunol. 2019, 10, 1928. [Google Scholar] [CrossRef] [Green Version]
- Naveca, F.G.; Pontes, G.S.; Chang, A.Y.; da Silva, G.A.V.; do Nascimento, V.A.; da Silva Monteiro, D.C.; da Silva, M.S.; Abdalla, L.F.; Santos, J.H.A.; de Almeida, T.A.P.; et al. Analysis of the Immunological Biomarker Profile during Acute Zika Virus Infection Reveals the Overexpression of CXCL10, a Chemokine Linked to Neuronal Damage. Mem. Inst. Oswaldo Cruz 2018, 113, e170542. [Google Scholar] [CrossRef] [PubMed]
- Puccioni-Sohler, M.; da Silva, S.J.; Faria, L.C.S.; Cabral, D.C.B.I.; Cabral-Castro, M.J. Neopterin and CXCL-10 in Cerebrospinal Fluid as Potential Biomarkers of Neuroinvasive Dengue and Chikungunya. Pathogens 2021, 10, 1626. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, S.; Khalfallah, O.; Forhan, A.; Galera, C.; Heude, B.; Glaichenhaus, N.; Davidovic, L. Immune Activity at Birth and Later Psychopathology in Childhood. Brain Behav. Immun. -Health 2020, 8, 100141. [Google Scholar] [CrossRef] [PubMed]
- Nascimento-Carvalho, G.C.; Nascimento-Carvalho, E.C.; Ramos, C.L.; Vilas-Boas, A.-L.; Moreno-Carvalho, O.A.; Vinhaes, C.L.; Barreto-Duarte, B.; Queiroz, A.T.L.; Andrade, B.B.; Nascimento-Carvalho, C.M. Zika-Exposed Microcephalic Neonates Exhibit Higher Degree of Inflammatory Imbalance in Cerebrospinal Fluid. Sci Rep. 2021, 11, 8474. [Google Scholar] [CrossRef]
- Venugopalan, A.; Ghorpade, R.P.; Chopra, A. Cytokines in Acute Chikungunya. PLoS ONE 2014, 9, e111305. [Google Scholar] [CrossRef]
- Renert-Yuval, Y.; Del Duca, E.; Pavel, A.B.; Fang, M.; Lefferdink, R.; Wu, J.; Diaz, A.; Estrada, Y.D.; Canter, T.; Zhang, N.; et al. The Molecular Features of Normal and Atopic Dermatitis Skin in Infants, Children, Adolescents, and Adults. J. Allergy Clin. Immunol. 2021, 148, 148–163. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Luan, B.; Wang, X.-F.; Qiao, J.-Y.; Song, L.; Lei, R.-R.; Gao, W.-X.; Liu, Y. Peripheral Blood MDSCs, IL-10 and IL-12 in Children with Asthma and Their Importance in Asthma Development. PLoS ONE 2013, 8, e63775. [Google Scholar] [CrossRef]
- Kinder, J.M.; Stelzer, I.A.; Arck, P.C.; Way, S.S. Immunological Implications of Pregnancy-Induced Microchimerism. Nat. Rev. Immunol. 2017, 17, 483–494. [Google Scholar] [CrossRef]
- Lamothe, J.; Khurana, S.; Tharmalingam, S.; Williamson, C.; Byrne, C.J.; Lees, S.J.; Khaper, N.; Kumar, A.; Tai, T.C. Oxidative Stress Mediates the Fetal Programming of Hypertension by Glucocorticoids. Antioxidants 2021, 10, 531. [Google Scholar] [CrossRef]
- Fitzgerald, W.; Gomez-Lopez, N.; Erez, O.; Romero, R.; Margolis, L. Extracellular Vesicles Generated by Placental Tissues ex vivo: A Transport System for Immune Mediators and Growth Factors. Am. J. Reprod. Immunol. 2018, 80, e12860. [Google Scholar] [CrossRef]
- Shenoy, S.; Pradeep, G.C.M. Neurodevelopmental Outcome of Neonates with Vertically Transmitted Chikungunya Fever with Encephalopathy. Indian Pediatr 2012, 49, 238–240. [Google Scholar] [PubMed]
- Waechter, R.; Ingraham, E.; Evans, R.; Cudjoe, N.; Krystosik, A.; Isaac, R.; Watts, A.; Noël, T.; Landon, B.; Fernandes, M.; et al. Pre and Postnatal Exposure to Chikungunya Virus Does Not Affect Child Neurodevelopmental Outcomes at Two Years of Age. PLoS Negl. Trop. Dis. 2020, 14, e0008546. [Google Scholar] [CrossRef] [PubMed]
- Decker, M.-L.; Gotta, V.; Wellmann, S.; Ritz, N. Cytokine Profiling in Healthy Children Shows Association of Age with Cytokine Concentrations. Sci Rep. 2017, 7, 17842. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A.; Carlsson, L.; Gordh, T.; Lind, A.-L.; Thulin, M.; Kamali-Moghaddam, M. The Effects of Age and Gender on Plasma Levels of 63 Cytokines. J. Immunol. Methods 2015, 425, 58–61. [Google Scholar] [CrossRef]
- Fordjour, L.; Cai, C.; Bronshtein, V.; Bronshtein, M.; Aranda, J.V.; Beharry, K.D. Growth Factors in the Fetus and Pre-Adolescent Offspring of Hyperglycemic Rats. Diabetes Vasc. Dis. Res. 2021, 18, 147916412110110. [Google Scholar] [CrossRef]
- Ramful, D.; Sampériz, S.; Fritel, X.; Michault, A.; Jaffar-Bandjee, M.-C.; Rollot, O.; Boumahni, B.; Gérardin, P. Antibody Kinetics in Infants Exposed to Chikungunya Virus Infection During Pregnancy Reveals Absence of Congenital Infection. J. Infect. Dis. 2014, 209, 1726–1730. [Google Scholar] [CrossRef]
- Couderc, T.; Chrétien, F.; Schilte, C.; Disson, O.; Brigitte, M.; Guivel-Benhassine, F.; Touret, Y.; Barau, G.; Cayet, N.; Schuffenecker, I.; et al. A Mouse Model for Chikungunya: Young Age and Inefficient Type-I Interferon Signaling Are Risk Factors for Severe Disease. PLoS Pathog. 2008, 4, e29. [Google Scholar] [CrossRef]
- Matusali, G.; Colavita, F.; Bordi, L.; Lalle, E.; Ippolito, G.; Capobianchi, M.R.; Castilletti, C. Tropism of the Chikungunya Virus. Viruses 2019, 11, 175. [Google Scholar] [CrossRef]
- Salomão, N.; Araújo, L.; Rabelo, K.; Avvad-Portari, E.; de Souza, L.; Fernandes, R.; Valle, N.; Ferreira, L.; Basílio-de-Oliveira, C.; Basílio-de-Oliveira, R.; et al. Placental Alterations in a Chikungunya-Virus-Infected Pregnant Woman: A Case Report. Microorganisms 2022, 10, 872. [Google Scholar] [CrossRef]
- Kawabata, H. The pathogenesis of anemia in inflammation. Rinsho Ketsueki 2020, 61, 1105–1111. [Google Scholar] [CrossRef]
- Slaats, J.; ten Oever, J.; van de Veerdonk, F.L.; Netea, M.G. IL-1β/IL-6/CRP and IL-18/Ferritin: Distinct Inflammatory Programs in Infections. PLoS Pathog. 2016, 12, e1005973. [Google Scholar] [CrossRef] [PubMed]
- Valero, N.; Mosquera, J.; Torres, M.; Duran, A.; Velastegui, M.; Reyes, J.; Fernandez, M.; Fernandez, G.; Veliz, T. Increased Serum Ferritin and Interleukin-18 Levels in Children with Dengue. Braz. J. Microbiol. 2019, 50, 649–656. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Clinical Features | CHIK (n = 33) | Control (n = 14) | p-Value |
|---|---|---|---|
| Infants, n (%) | |||
| Age (months. mean ± SD) | 3.0 ± 2.9 | 9.9 ± 7.2 | <0.0001 |
| Male Gender | 19 (57.0) | 8 (57.1) | >0.9 |
| Postnatal sepsis | 1 (3.03) | 0 (0) | 0.9 |
| Neurodevelopment disorders | 1 (3.03) | 0 (0) | 0.9 |
| Mothers, n (%) | |||
| Age (year. mean ± SD) | 26.8 ± 5.6 | 29.9 ± 6.2 | >0.9 |
| Time of maternal rash | |||
| 1th Trimester | 6 (18.2) | 3 (21.4) | 0.7 |
| 2nd Trimester | 13 (39.4) | 6 (42.8) | 0.7 |
| 3rd Trimester | 14 (42.4) | 5 (35.7) | >0.9 |
| Clinical symptoms | |||
| Fever | 26 (78.8) | 7 (50) | 0.08 |
| Myalgia | 33 (100) | 4 (28.6) | <0.0001 |
| Arthralgia | 33 (100) | 7 (50) | <0.0001 |
| Parameters | Reference Range | CHIK (n = 33) | Control (n = 14) | p-Value |
|---|---|---|---|---|
| Alanine aminotransferase (U/L) | 14.0–50.0 | 51.4 ± 46.1 | 41.1 ± 11.5 | 0.9 |
| Aspartate aminotransferase (U/L) | 15.0–37.0 | 36.9 ± 46.0 | 25.1 ± 9.4 | 0.3 |
| Total Bilirrubin (mg/dL) | 0.2–1.0 | 1.5 ± 2.2 | 0.3 ± 0.1 | 0.2 |
| Direct Bilirrubin (mg/dL) | > 0.3 | 0.2 ± 0.1 | 0.1 ± 0.07 | 0.7 |
| Indirect Bilirrubin (mg/dL) | - | 1.4 ± 2.1 | 0.2 ± 0.1 | 0.06 |
| Ferritin (ng/dL) | 26–388.0 | 214.3 ± 219.0 | 89.5 ± 58.1 | 0.1 |
| C-reactive Protein (mg/dL) | 0.1–1 | 0.4 ± 0.1 | 0.1 ± 0.08 | 0.02 |
| Lactate dehydrogenase (U/L) | 85.0–227.0 | 327.2 ± 81.3 | 375.5 ± 173.9 | 0.5 |
| Hemoglobin (g/dL) | 11.0–14.0 | 11.8 ± 1.9 | 11.4 ± 3.3 | 0.05 |
| Hematocrit (%) | 33.0–39.0 | 35.2 ± 5.4 | 34.8 ± 9.0 | 0.1 |
| Platelets (103/mm3) | 150.0–400.0 | 400.6 ± 166.4 | 477.2 ± 162.3 | 0.1 |
| White blood cell count (103/mm3) | 5.0–17.0 | 11.1 ± 5.2 | 11.8 ± 4.6 | 0.8 |
| Neutrophils | 1–7 | 3.6 ± 1.9 | 3.7 ± 2.2 | 0.7 |
| Lymphocytes | 3.5–11 | 5.8 ± 1.9 | 5.4 ± 1.4 | 0.9 |
| Monocytes | 0.2–1 | 0.7 ± 0.4 | 0.9 ± 0.4 | 0.2 |
| Eosinophils | 0.1–1 | 0.3 ± 0.2 | 0.3 ± 0.1 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faustino, R.; Carvalho, F.R.; Medeiros, T.; Familiar-Macedo, D.; Vianna, R.A.d.O.; Leite, P.E.C.; Pereira, I.R.; Cardoso, C.A.A.; De Azeredo, E.L.; Silva, A.A. Pro-Inflammatory Profile of Children Exposed to Maternal Chikungunya Virus Infection during the Intrauterine Period: A One-Year Follow-Up Study. Viruses 2022, 14, 1881. https://doi.org/10.3390/v14091881
Faustino R, Carvalho FR, Medeiros T, Familiar-Macedo D, Vianna RAdO, Leite PEC, Pereira IR, Cardoso CAA, De Azeredo EL, Silva AA. Pro-Inflammatory Profile of Children Exposed to Maternal Chikungunya Virus Infection during the Intrauterine Period: A One-Year Follow-Up Study. Viruses. 2022; 14(9):1881. https://doi.org/10.3390/v14091881
Chicago/Turabian StyleFaustino, Renan, Fabiana Rabe Carvalho, Thalia Medeiros, Débora Familiar-Macedo, Renata Artimos de Oliveira Vianna, Paulo Emílio Côrrea Leite, Isabela Resende Pereira, Claudete Aparecida Araújo Cardoso, Elzinandes Leal De Azeredo, and Andrea Alice Silva. 2022. "Pro-Inflammatory Profile of Children Exposed to Maternal Chikungunya Virus Infection during the Intrauterine Period: A One-Year Follow-Up Study" Viruses 14, no. 9: 1881. https://doi.org/10.3390/v14091881