
Seed Transmission of Tomato Spotted Wilt Orthotospovirus in Peppers

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
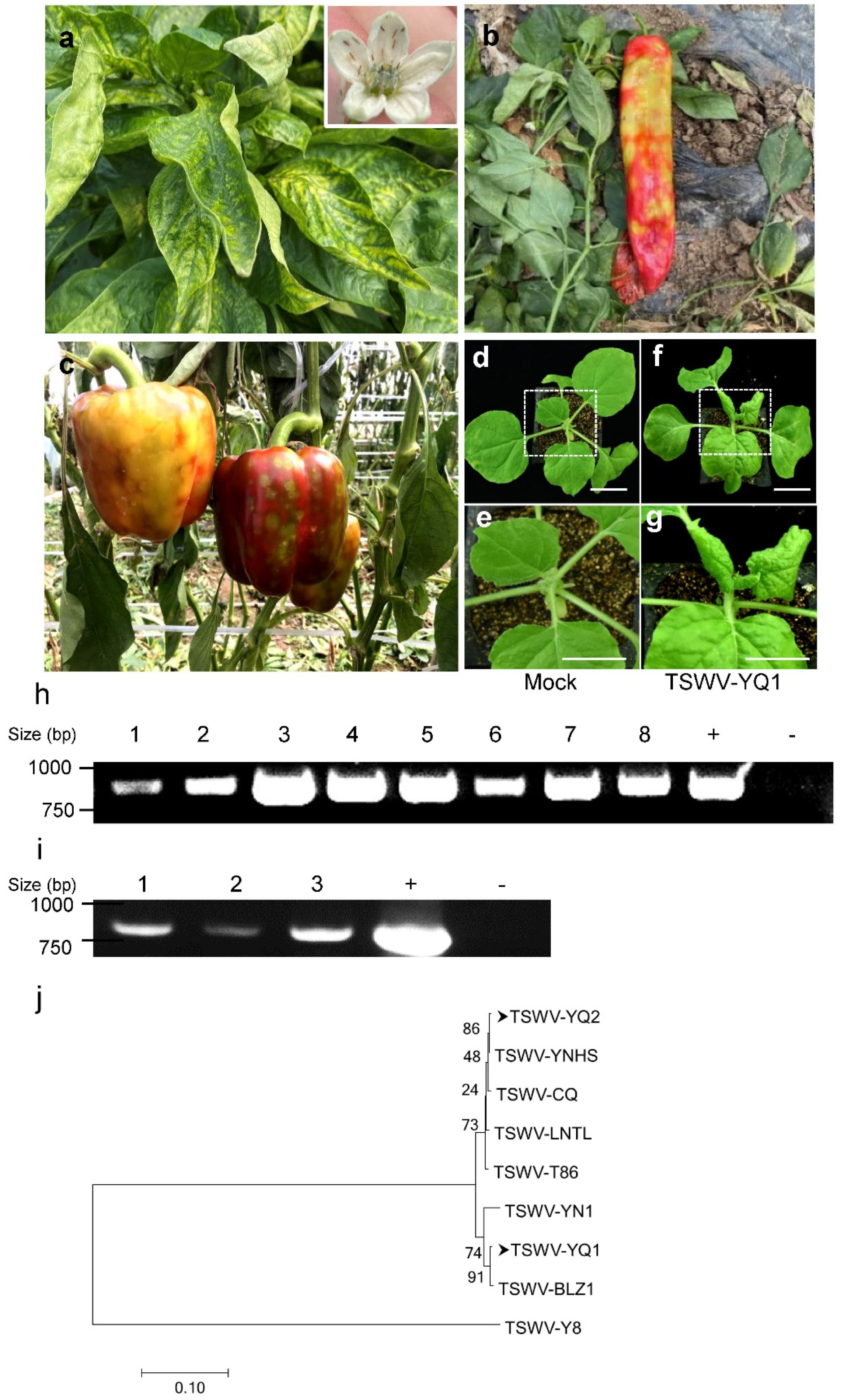
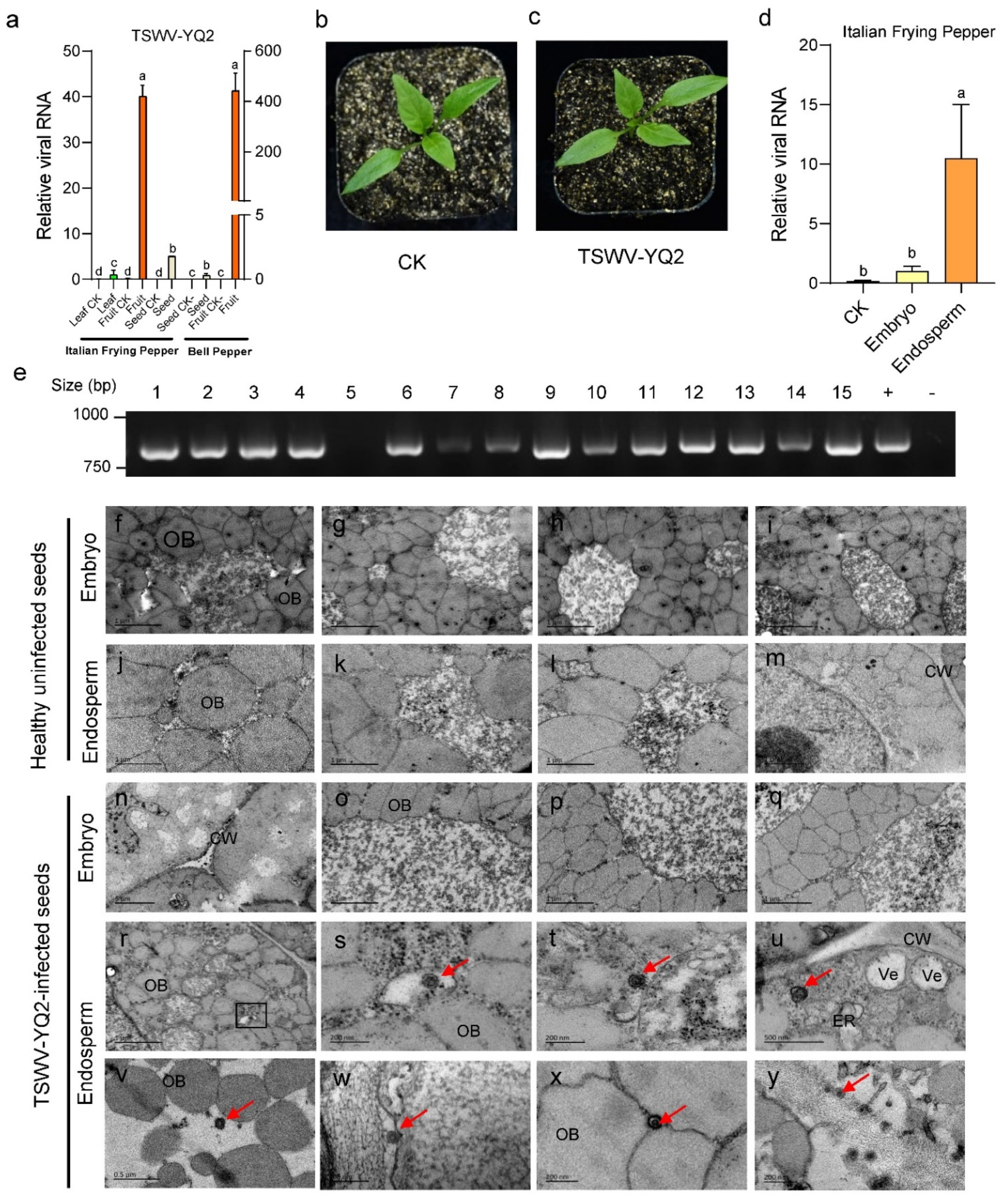
2.1. The Identification and Detection of TSWV in Peppers
2.2. TSWV Is Seed-Transmissible
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Inoculation of Plants
4.3. Dissection of Pepper Seeds
4.4. RT-PCR and RT-qPCR
4.5. Transmission Electron Microscopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allard, H.A. The mosaic disease of tobacco. Science 1912, 36, 875–876. [Google Scholar] [CrossRef] [PubMed]
- Hull, R.; Matthews, R.E.F. Matthews’ Plant Virology, 4th ed.; Elsevier Academic Press: San Diego, CA, USA, 2002; pp. 533–581. [Google Scholar]
- Sastry; Subramanya, K. Seed-Borne Plant Virus Diseases; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Natsuhara, D.; Takishita, K.; Tanaka, K.; Kage, A.; Suzuki, R.; Mizukami, Y.; Saka, N.; Nagai, M.; Shibata, T. A Microfluidic Diagnostic Device Capable of Autonomous Sample Mixing and Dispensing for the Simultaneous Genetic Detection of Multiple Plant Viruses. Micromachines 2020, 11, 540. [Google Scholar] [CrossRef] [PubMed]
- Pooggin, M.M. Small RNA-Omics for Plant Virus Identification, Virome Reconstruction, and Antiviral Defense Characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; van Grinsven, I.L.; Kormelink, R.; Tao, X. Paving the Way to Tospovirus Infection: Multilined Interplays with Plant Innate Immunity. Annu. Rev. Phytopathol. 2019, 57, 41–62. [Google Scholar] [CrossRef] [PubMed]
- Turina, M.; Kormelink, R.; Resende, R.O. Resistance to Tospoviruses in Vegetable Crops: Epidemiological and Molecular Aspects. Annu. Rev. Phytopathol. 2016, 54, 347–371. [Google Scholar] [CrossRef] [PubMed]
- Roselló, S.; Díez, M.; Nuez, F. Viral diseases causing the greatest economic losses to the tomato crop. I. The Tomato Spotted Wilt Virus-a review. Sci. Hortic. 1996, 67, 117–150. [Google Scholar] [CrossRef]
- Wijkamp, I.; Almarza, N.; Goldbach, R.; Peters, D. Distinct levels of specificity in thrips transmission of Tospoviruses. Phytopathology 1995, 85, 1069–1074. [Google Scholar] [CrossRef]
- Jones, L. Streak and mosaic of Cineraria. Phytopathology 1944, 34, 941–953. [Google Scholar]
- Crowley, N.C. Studies on the seed transmission of plant virus diseases. Aust. J. Biol. Sci. 1957, 10, 449–464. [Google Scholar] [CrossRef]
- Antignus, Y.; Lapidot, M.; Ganaim, N.; Cohen, J.; Lachman, O.; Pearlsman, M.; Raccah, B.; Gera, A.J.P. Biological and molecular characterization of Tomato spotted wilt Virus in Israel. Phytoparasitica 1997, 25, 319–330. [Google Scholar] [CrossRef]
- Groves, C.; German, T.; Dasgupta, R.; Mueller, D.; Smith, D.L. Seed Transmission of Soybean vein necrosis virus: The First Tospovirus Implicated in Seed Transmission. PLoS ONE 2016, 11, e0147342. [Google Scholar] [CrossRef]
- Scholthof, K.B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, L.; Zhang, X.; Wu, X.; Fang, R. Plant Defense Networks against Insect-Borne Pathogens. Trends Plant Sci. 2021, 26, 272–287. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ye, J. Manipulation of Jasmonate Signaling by Plant Viruses and Their Insect Vectors. Viruses 2020, 12, 148. [Google Scholar] [CrossRef]
- Pappu, H.R.; Jones, R.A.; Jain, R.K. Global status of tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res. 2009, 141, 219–236. [Google Scholar] [CrossRef]
- French, J.M.; Goldberg, N.P.; Randall, J.J.; Hanson, S.F. New Mexico and the southwestern US are affected by a unique population of tomato spotted wilt virus (TSWV) strains. Arch. Virol. 2016, 161, 993–998. [Google Scholar] [CrossRef]
- Sevik, M.A.; Arli-Sokmen, M. Estimation of the effect of Tomato spotted wilt virus (TSWV) infection on some yield components of tomato. Phytoparasitica 2012, 40, 87–93. [Google Scholar] [CrossRef]
- Adkins, S. Tomato spotted wilt virus-positive steps towards negative success. Mol. Plant Pathol. 2000, 1, 151–157. [Google Scholar] [CrossRef]
- de Haan, P.; Kormelink, R.; de Oliveira Resende, R.; van Poelwijk, F.; Peters, D.; Goldbach, R. Tomato spotted wilt virus L RNA encodes a putative RNA polymerase. J. Gen. Virol. 1991, 72, 2207–2216. [Google Scholar] [CrossRef]
- Adkins, S.; Quadt, R.; Choi, T.J.; Ahlquist, P.; German, T. An RNA-dependent RNA polymerase activity associated with virions of tomato spotted wilt virus, a plant-and insect-infecting bunyavirus. Virology 1995, 207, 308–311. [Google Scholar] [CrossRef]
- Lee, J.S.; Cho, W.K.; Kim, M.K.; Kwak, H.R.; Choi, H.S.; Kim, K.H. Complete genome sequences of three tomato spotted wilt virus isolates from tomato and pepper plants in Korea and their phylogenetic relationship to other TSWV isolates. Arch. Virol. 2011, 156, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Margaria, P.; Bosco, L.; Vallino, M.; Ciuffo, M.; Mautino, G.C.; Tavella, L.; Turina, M. The NSs protein of tomato spotted wilt virus is required for persistent infection and transmission by Frankliniella occidentalis. J. Virol. 2014, 88, 5788–5802. [Google Scholar] [CrossRef] [PubMed]
- de Avila, A.C.; de Haan, P.; Kormelink, R.; Resende Rde, O.; Goldbach, R.W.; Peters, D. Classification of tospoviruses based on phylogeny of nucleoprotein gene sequences. J. Gen. Virol. 1993, 74, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Tsompana, M.; Abad, J.; Purugganan, M.; Moyer, J.W. The molecular population genetics of the Tomato spotted wilt virus (TSWV) genome. Mol. Ecol. 2005, 14, 53–66. [Google Scholar] [CrossRef]
- Oliver, J.E.; Whitfield, A.E. The Genus Tospovirus: Emerging Bunyaviruses that Threaten Food Security. Annu. Rev. Virol. 2016, 3, 101–124. [Google Scholar] [CrossRef]
- Wu, X.; Xu, S.; Zhao, P.; Zhang, X.; Yao, X.; Sun, Y.; Fang, R.; Ye, J. The Orthotospovirus nonstructural protein NSs suppresses plant MYC-regulated jasmonate signaling leading to enhanced vector attraction and performance. PLoS Pathog. 2019, 15, e1007897. [Google Scholar] [CrossRef]
- Zhao, P.; Yao, X.; Cai, C.; Li, R.; Du, J.; Sun, Y.; Wang, M.; Zou, Z.; Wang, Q.; Kliebenstein, D.J.; et al. Viruses mobilize plant immunity to deter nonvector insect herbivores. Sci. Adv. 2019, 5, eaav9801. [Google Scholar] [CrossRef]
- Sandra, N.; Tripathi, A.; Dikshit, H.K.; Mandal, B.; Jain, R.K. Seed transmission of a distinct soybean yellow mottle mosaic virus strain identified from India in natural and experimental hosts. Virus Res. 2020, 280, 197903. [Google Scholar] [CrossRef]
- Johansen, E. Seed Transmission of Viruses: Current Perspectives. Annu. Rev. Phytopathol. 1994, 32, 363–386. [Google Scholar] [CrossRef]
- Schaad, N.W. Inoculum Thresholds of Soilborne Pathogens. Phytopathology 1988, 78, 872. [Google Scholar] [CrossRef]
- Ellis, M.D.; Hoak, J.M.; Ellis, B.W.; Brown, J.A.; Sit, T.L.; Wilkinson, C.A.; Reed, T.D.; Welbaum, G.E. Quantitative Real-Time PCR Analysis of Individual Flue-Cured Tobacco Seeds and Seedlings Reveals Seed Transmission of Tobacco Mosaic Virus. Phytopathology 2020, 110, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Suruthi, V.; Nakkeeran, S.; Renukadevi, P.; Malathi, V.G.; Rajasree, V. Evidence of seed transmission of dolichos yellow mosaic virus, a begomovirus infecting lablab-bean in India. Virusdisease 2018, 29, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, K.; Dong, J.; Fang, Q.; Hong, J.; Wang, X. Clustering and cellular distribution characteristics of virus particles of Tomato spotted wilt virus and Tomato zonate spot virus in different plant hosts. Virol. J. 2016, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Carrington, J.C.; Kasschau, K.D.; Mahajan, S.K.; Schaad, M.C. Cell-to-Cell and Long-Distance Transport of Viruses in Plants. Plant Cell 1996, 8, 1669–1681. [Google Scholar] [CrossRef]
- Karmann, J.; Müller, B.; Hammes, U.Z. The long and winding road: Transport pathways for amino acids in Arabidopsis seeds. Plant Reprod. 2018, 31, 253–261. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, S.; Zhou, Y.; Bai, J.; Huang, G.; Liu, X.; Zhang, Y.; Tang, D.; Lu, D. Transcriptional Regulation of the Immune Receptor FLS2 Controls the Ontogeny of Plant Innate Immunity. Plant Cell 2018, 30, 2779–2794. [Google Scholar] [CrossRef]
- Kothandaraman, S.V.; Devadason, A.; Ganesan, M.V. Seed-borne nature of a begomovirus, Mung bean yellow mosaic virus in black gram. Appl. Microbiol. Biotechnol. 2016, 100, 1925–1933. [Google Scholar] [CrossRef]
- Mandal, B.; Csinos, A.S.; Martinez-Ochoa, N.; Pappu, H.R. A rapid and efficient inoculation method for Tomato spotted wilt tospovirus. J. Virol. Methods 2008, 149, 195–198. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sequence | TSWV-YQ1 | TSWV-YQ2 |
|---|---|---|
| TSWV-YQ1 | — | 96.69% |
| TSWV-YQ2 | 96.69% | — |
| TSWV-BLZ1 | 99.72% | 96.71% |
| TSWV-YNHS | 96.84% | 99.86% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wu, X.; Huang, X.; Wei, S.; Lu, Z.; Ye, J. Seed Transmission of Tomato Spotted Wilt Orthotospovirus in Peppers. Viruses 2022, 14, 1873. https://doi.org/10.3390/v14091873
Wang H, Wu X, Huang X, Wei S, Lu Z, Ye J. Seed Transmission of Tomato Spotted Wilt Orthotospovirus in Peppers. Viruses. 2022; 14(9):1873. https://doi.org/10.3390/v14091873
Chicago/Turabian StyleWang, Hongwei, Xiujuan Wu, Xiande Huang, Shujun Wei, Zhijun Lu, and Jian Ye. 2022. "Seed Transmission of Tomato Spotted Wilt Orthotospovirus in Peppers" Viruses 14, no. 9: 1873. https://doi.org/10.3390/v14091873