
Differential Expression of Genes between a Tolerant and a Susceptible Maize Line in Response to a Sugarcane Mosaic Virus Infection



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Virus and Plant Materials
2.2. Experimental Design and Sequencing
2.3. Mapping and Differential Expression Analysis
2.4. Construction of CTEPs
2.5. Functional Enrichment
2.6. Gene Expression Validation
3. Results
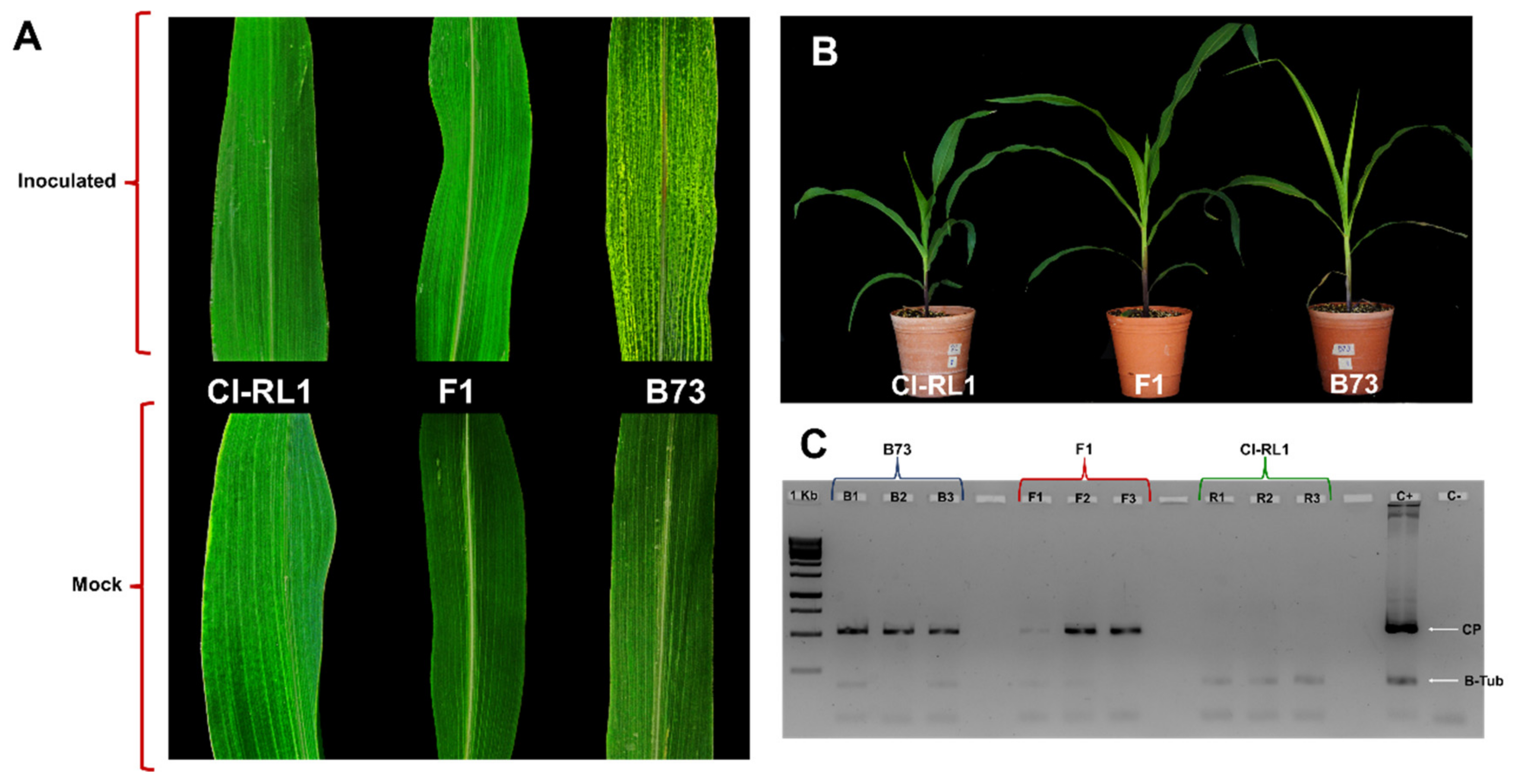
3.1. Symptom Development in B73, CI-RL 1, and F1 Progeny
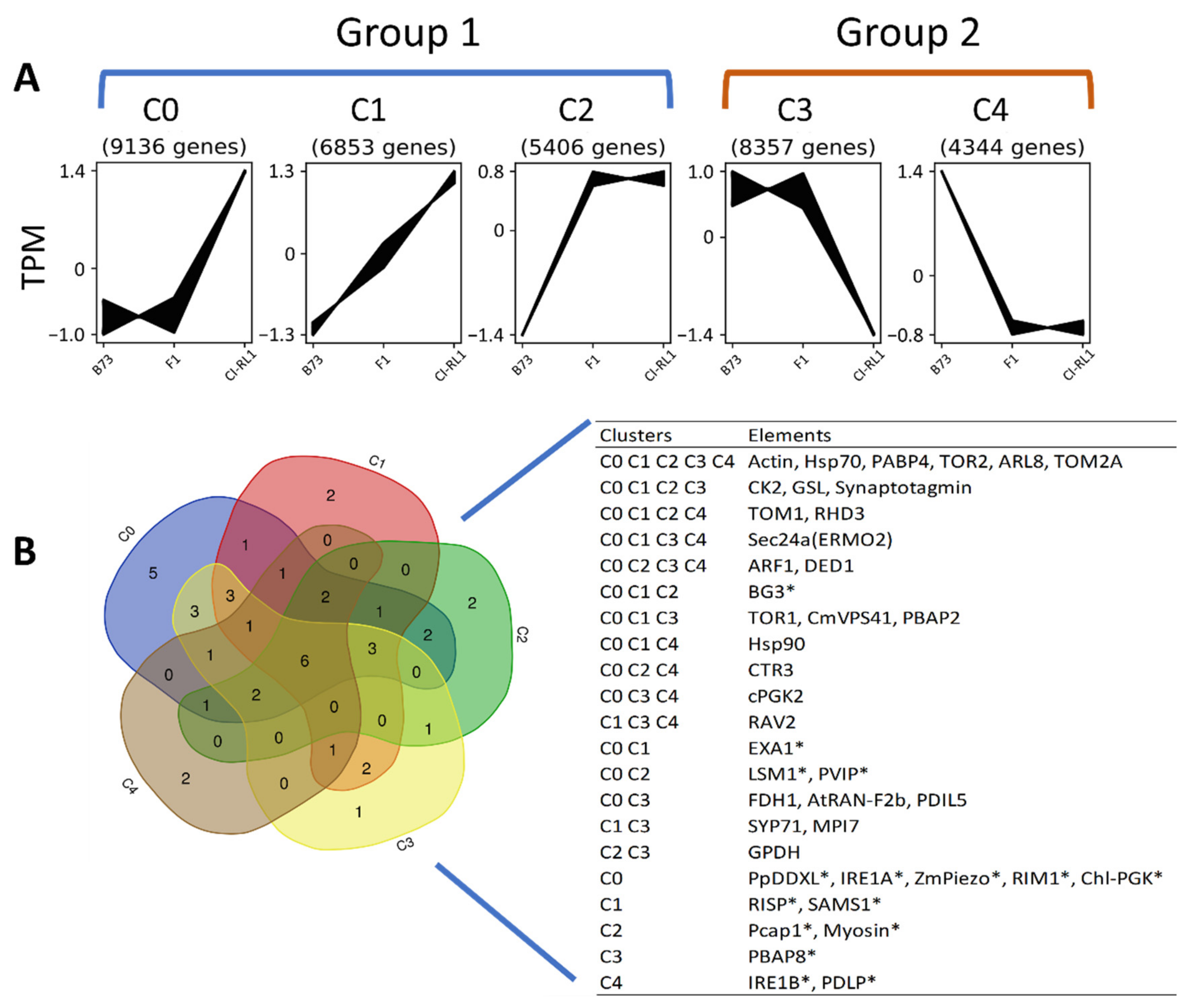
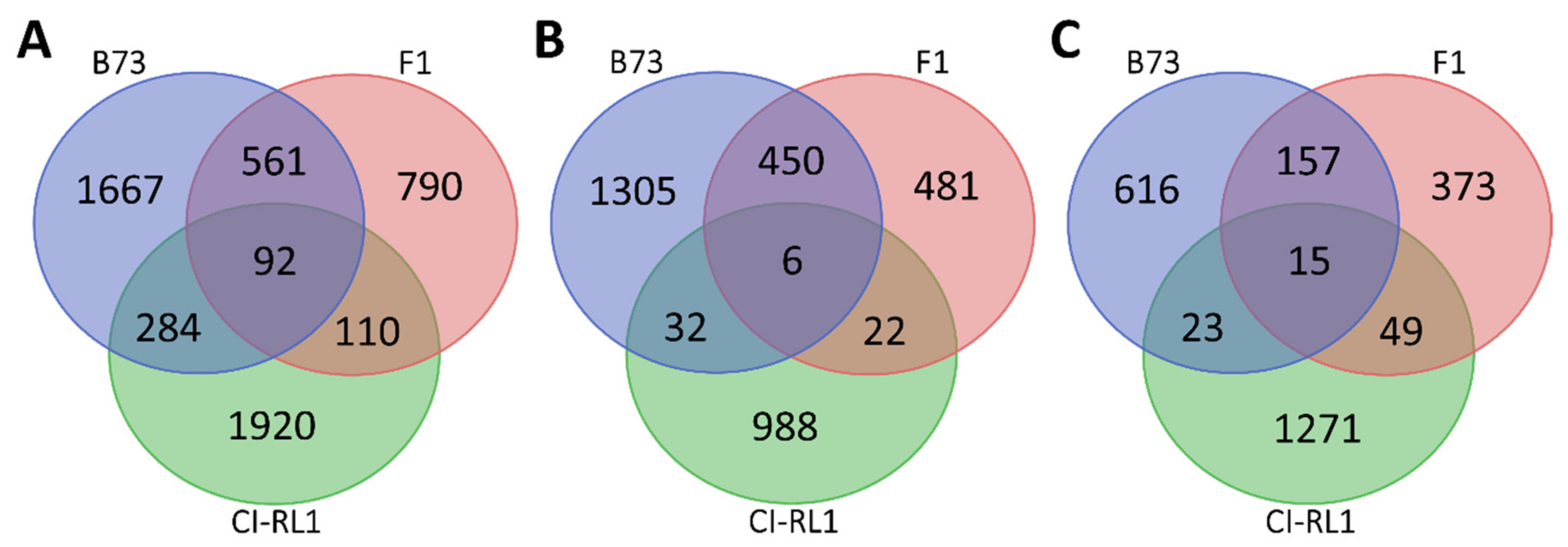
3.2. Gene Expression Profiling Is Similar between Susceptible B73 and F1 Progeny
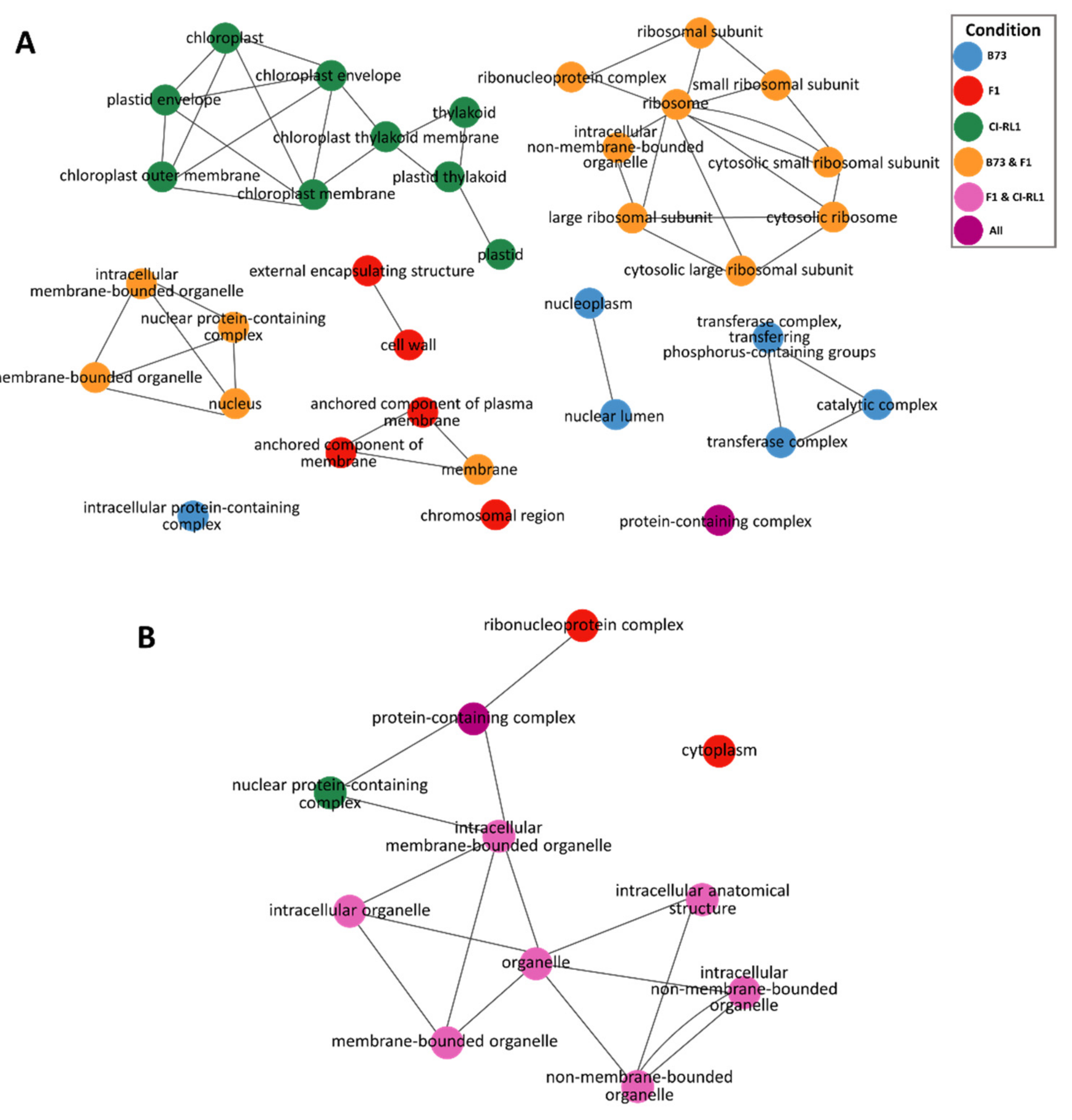
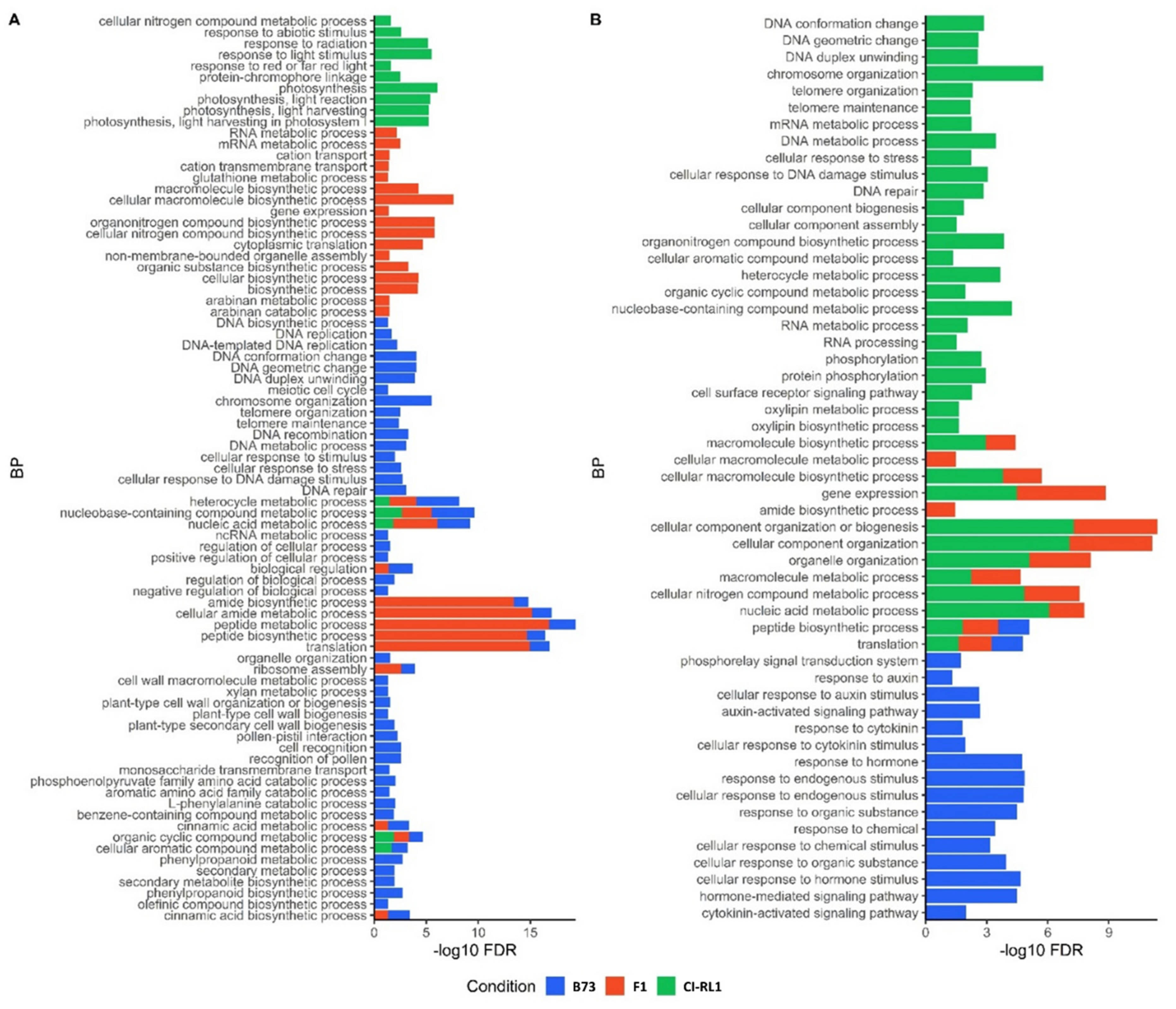
3.3. Up- and Downregulation of Enriched Cellular Components after SCMV Infection
3.4. Enriched Biological Process Shows the Predominance of Downregulated CI-RL1 Transcripts
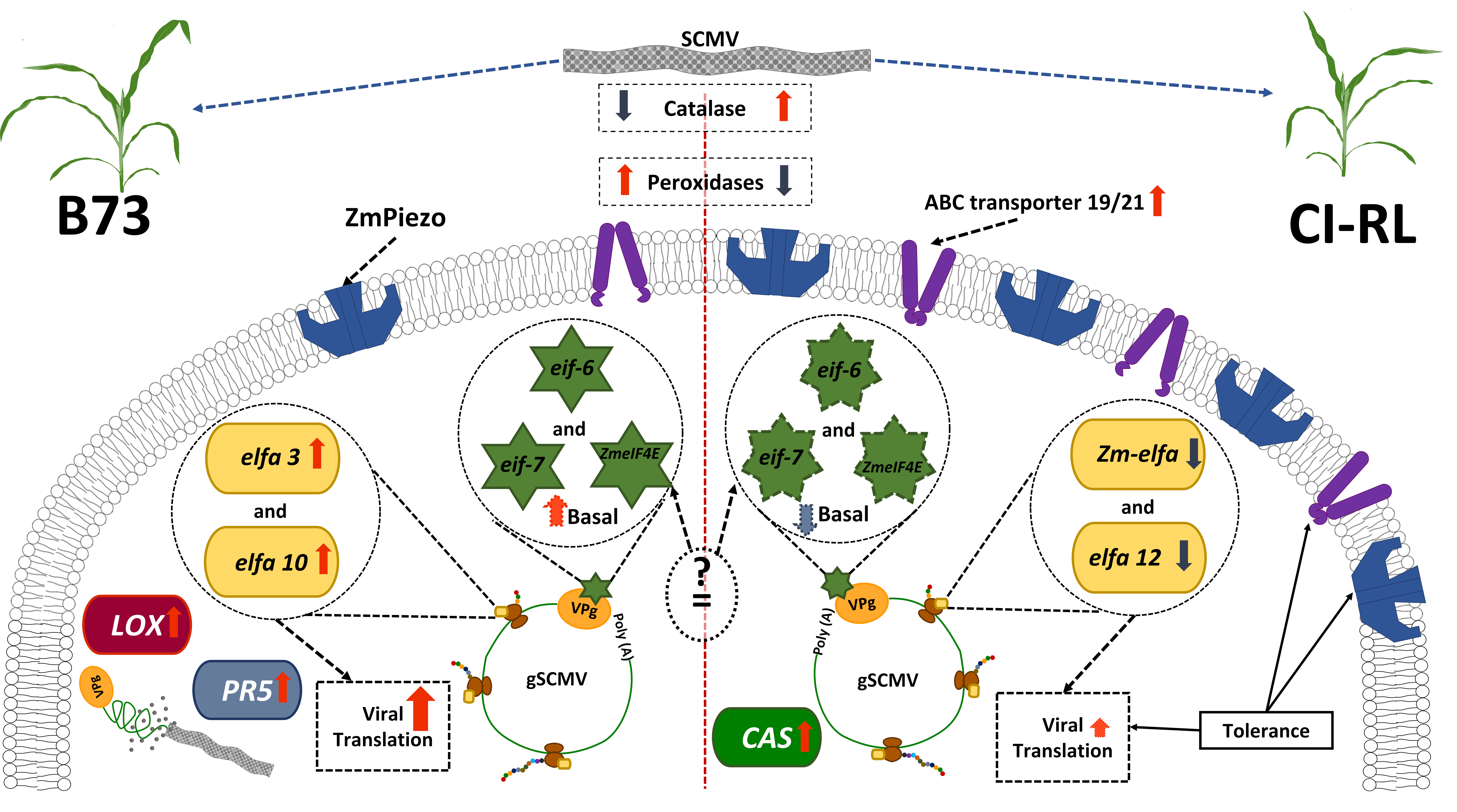
3.5. Candidate Genes Associated with the CI-RL 1 Tolerance/Resistance Response to SCMV Were Selected with Different Criteria
3.6. Thirteen Selected Candidate Genes Have High Basal Expression in CI-RL1
3.7. Differences in Translation and Elongation Factor Coding Transcripts in the CI-RL1 Line
4. Discussion
4.1. Resistance or Tolerance of the CI-RL1 Line
4.2. DEG Analysis Pinpoints Multi-Genic Tolerance
4.3. PR5 and LDOX: Two Putative Susceptibility Factors in B73?
4.4. Genes Involved in Virus Replication Found in CTEP Analyses
4.5. Cell-to-Cell Movement-Related Genes
4.6. Long-Distance Movement-Related Genes
4.7. The Presence of Diverse Transcripts for eIFs and eEFs Implies the Complex Regulation of Translation in SCMV Interaction with Maize Transcripts in Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva-Rosales, L.; Alcalá-Briseño, R.I.; Espejel, F. Sugarcane Mosaic. In Virus Diseases of Tropical and Subtropical Crops; Tennant, P., Fermin, G., Eds.; CABI: Wallingford, UK, 2015; pp. 131–143. ISBN 978-1-78064-426-4. [Google Scholar]
- Xie, L.; Zhang, J.; Wang, Q.; Meng, C.; Hong, J.; Zhou, X. Characterization of Maize Chlorotic Mottle Virus Associated with Maize Lethal Necrosis Disease in China. J. Phytopathol. 2011, 159, 191–193. [Google Scholar] [CrossRef]
- Fatma, H.K.; Tileye, F.; Patrick, A.N. Insights of Maize Lethal Necrotic Disease: A Major Constraint to Maize Production in East Africa. Afr. J. Microbiol. Res. 2016, 10, 271–279. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Kuntze, L.; Gumber, R.K.; Lübberstedt, T.; Fuchs, E. Genetic Basis of Resistance to Sugarcane Mosaic Virus in European Maize Germplasm. Theor. Appl. Genet. 1998, 96, 1151–1161. [Google Scholar] [CrossRef]
- Lubberstedt, T.; Ingvardsen, C.; Melchinger, A.E.; Xing, Y.; Salomon, R.; Redinbaugh, M.G. Two Chromosome Segments Confer Multiple Potyvirus Resistance in Maize. Plant Breed. 2006, 125, 352–356. [Google Scholar] [CrossRef]
- Wu, J.; Ding, J.; Du, Y.; Xu, Y.; Zhang, X. Genetic Analysis and Molecular Mapping of Two Dominant Complementary Genes Determining Resistance to Sugarcane Mosaic Virus in Maize. Euphytica 2007, 156, 355–364. [Google Scholar] [CrossRef]
- Pokorný, R.; Porubová, M. Movement of Sugarcane Mosaic Virus in Plants of Resistant and Susceptible Maize Lines. Cereal Res. Commun. 2006, 34, 1109–1116. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An Atypical Thioredoxin Imparts Early Resistance to Sugarcane Mosaic Virus in Maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.V.; Rey, P. Plant Thioredoxins Are Key Actors in the Oxidative Stress Response. Trends Plant Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Hisabori, T.; Motohashi, K.; Hosoya-Matsuda, N.; Ueoka-Nakanishi, H.; Romano, P.G.N. Towards a Functional Dissection of Thioredoxin Networks in Plant Cells. Photochem. Photobiol. 2006, 83, 145–151. [Google Scholar] [CrossRef]
- Chaves-Bedoya, G.; Espejel, F.; Alcalá-Briseño, R.I.; Hernández-Vela, J.; Silva-Rosales, L. Short Distance Movement of Genomic Negative Strands in a Host and Nonhost for Sugarcane Mosaic Virus (SCMV). Virol. J. 2011, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Dangl, J.L.; Jones, J.D.G. Plant Pathogens and Integrated Defence Responses to Infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Ausubel, F.M. Are Innate Immune Signaling Pathways in Plants and Animals Conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe Interactions: Shaping the Evolution of the Plant Immune Response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Nurnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate Immunity in Plants and Animals: Striking Similarities and Obvious Differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling Mechanisms in Pattern-Triggered Immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef]
- DeYoung, B.J.; Innes, R.W. Plant NBS-LRR Proteins in Pathogen Sensing and Host Defense. Nat. Immunol. 2006, 7, 1243–1249. [Google Scholar] [CrossRef]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Liu, L.; Sonbol, F.-M.; Huot, B.; Gu, Y.; Withers, J.; Mwimba, M.; Yao, J.; He, S.Y.; Dong, X. Salicylic Acid Receptors Activate Jasmonic Acid Signalling through a Non-Canonical Pathway to Promote Effector-Triggered Immunity. Nat. Commun. 2016, 7, 13099. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant–Pathogen Warfare under Changing Climate Conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef]
- Saleem, M.; Fariduddin, Q.; Castroverde, C.D.M. Salicylic Acid: A Key Regulator of Redox Signalling and Plant Immunity. Plant Physiol. Biochem. 2021, 168, 381–397. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J.; Nürnberger, T.; Joosten, M.H.A.J. Of PAMPs and Effectors: The Blurred PTI-ETI Dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef]
- Nejat, N.; Mantri, N. Plant Immune System: Crosstalk Between Responses to Biotic and Abiotic Stresses the Missing Link in Understanding Plant Defence. Curr. Issues Mol. Biol. 2017, 23, 1–16. [Google Scholar] [CrossRef]
- Zvereva, A.; Pooggin, M. Silencing and Innate Immunity in Plant Defense Against Viral and Non-Viral Pathogens. Viruses 2012, 4, 2578–2597. [Google Scholar] [CrossRef]
- Teixeira, R.M.; Ferreira, M.A.; Raimundo, G.A.S.; Loriato, V.A.P.; Reis, P.A.B.; Fontes, E.P.B. Virus Perception at the Cell Surface: Revisiting the Roles of Receptor-like Kinases as Viral Pattern Recognition Receptors. Mol. Plant Pathol. 2019, 20, 1196–1202. [Google Scholar] [CrossRef]
- Wang, M.-B.; Masuta, C.; Smith, N.A.; Shimura, H. RNA Silencing and Plant Viral Diseases. Mol. Plant-Microbe Interact. 2012, 25, 1275–1285. [Google Scholar] [CrossRef]
- Waterhouse, P.M.; Wang, M.-B.; Lough, T. Gene Silencing as an Adaptive Defence against Viruses. Nature 2001, 411, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.B.; Santos, A.A.; Fontes, E.P.B. Immune Receptors and Co-Receptors in Antiviral Innate Immunity in Plants. Front. Microbiol. 2017, 7, 2139. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, M. Plant Virus Replication and Movement. Virology 2015, 479–480, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, K. Plant Susceptibility Genes as a Source for Potyvirus Resistance. Ann. Appl. Biol. 2020, 176, 122–129. [Google Scholar] [CrossRef]
- Wang, A. Dissecting the Molecular Network of Virus-Plant Interactions: The Complex Roles of Host Factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef]
- Bastet, A.; Robaglia, C.; Gallois, J.-L. EIF4E Resistance: Natural Variation Should Guide Gene Editing. Trends Plant Sci. 2017, 22, 411–419. [Google Scholar] [CrossRef]
- Nicaise, V.; Gallois, J.-L.; Chafiai, F.; Allen, L.M.; Schurdi-Levraud, V.; Browning, K.S.; Candresse, T.; Caranta, C.; Le Gall, O.; German-Retana, S. Coordinated and Selective Recruitment of EIF4E and EIF4G Factors for Potyvirus Infection in Arabidopsis thaliana. FEBS Lett. 2007, 581, 1041–1046. [Google Scholar] [CrossRef]
- Hafrén, A.; Hofius, D.; Rönnholm, G.; Sonnewald, U.; Mäkinen, K. HSP70 and Its Cochaperone CPIP Promote Potyvirus Infection in Nicotiana benthamiana by Regulating Viral Coat Protein Functions. Plant Cell 2010, 22, 523–535. [Google Scholar] [CrossRef]
- Lubkowska, A.; Pluta, W.; Strońska, A.; Lalko, A. Role of Heat Shock Proteins (HSP70 and HSP90) in Viral Infection. Int. J. Mol. Sci. 2021, 22, 9366. [Google Scholar] [CrossRef]
- Gorovits, R.; Czosnek, H. The Involvement of Heat Shock Proteins in the Establishment of Tomato Yellow Leaf Curl Virus Infection. Front. Plant Sci. 2017, 8, 355. [Google Scholar] [CrossRef]
- Sullivan, C.S.; Pipas, J.M. The Virus–Chaperone Connection. Virology 2001, 287, 1–8. [Google Scholar] [CrossRef]
- Park, C.-J.; Seo, Y.-S. Heat Shock Proteins: A Review of the Molecular Chaperones for Plant Immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef]
- Beris, D.; Kotsaridis, K.; Vakirlis, N.; Termentzi, A.; Theologidis, I.; Moury, B.; Vassilakos, N. The Plasma Membrane Cation Binding Protein 1 Affects Accumulation of Potato Virus Y in Pepper Both at the Systemic Level and in Protoplasts. Virus Res. 2020, 280, 197899. [Google Scholar] [CrossRef]
- Dunoyer, P.; Thomas, C.; Harrison, S.; Revers, F.; Maule, A. A Cysteine-Rich Plant Protein Potentiates Potyvirus Movement through an Interaction with the Virus Genome-Linked Protein VPg. J. Virol. 2004, 78, 2301–2309. [Google Scholar] [CrossRef]
- Clarke, D. Tolerance of Parasites and Disease in Plants and Its Significance in Host-Parasite Interactions. Adv. Plant Pathol. 1986, 5, 161–197. [Google Scholar]
- Strauss, S.Y.; Agrawal, A.A. The Ecology and Evolution of Plant Tolerance to Herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Pagán, I.; García-Arenal, F. Tolerance to Plant Pathogens: Theory and Experimental Evidence. Int. J. Mol. Sci. 2018, 19, 810. [Google Scholar] [CrossRef]
- Little, T.J.; Shuker, D.M.; Colegrave, N.; Day, T.; Graham, A.L. The Coevolution of Virulence: Tolerance in Perspective. PLoS Pathog. 2010, 6, e1001006. [Google Scholar] [CrossRef]
- Råberg, L. How to Live with the Enemy: Understanding Tolerance to Parasites. PLoS Biol. 2014, 12, e1001989. [Google Scholar] [CrossRef] [PubMed]
- Del Fabbro, C.; Scalabrin, S.; Morgante, M.; Giorgi, F. An Extensive Evaluation of Read Trimming Effects on Illumina NGS Data Analysis. PLoS ONE 2013, 8, e85024. [Google Scholar] [CrossRef]
- Vargas-Mejía, P.; Vega-Arreguín, J.; Chávez-Calvillo, G.; Ibarra-Laclette, E.; Silva-Rosales, L. Differential Accumulation of Innate- and Adaptive-Immune-Response-Derived Transcripts during Antagonism between Papaya Ringspot Virus and Papaya Mosaic Virus. Viruses 2020, 12, 230. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Pimentel, H.; Bray, N.L.; Pachter, L. Gene-Level Differential Analysis at Transcript-Level Resolution. Genome Biol. 2018, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Abu-Jamous, B.; Kelly, S. Clust: Automatic Extraction of Optimal Co-Expressed Gene Clusters from Gene Expression Data. Genome Biol. 2018, 19, 172. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.-H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl De Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef]
- Kang, B.-C.; Yeam, I.; Jahn, M.M. Genetics of Plant Virus Resistance. Annu. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Susceptibility Genes to Plant Viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef]
- Lellis, A.D.; Kasschau, K.D.; Whitham, S.A.; Carrington, J.C. Loss-of-Susceptibility Mutants of Arabidopsis thaliana Reveal an Essential Role for EIF(Iso)4E during Potyvirus Infection. Curr. Biol. 2002, 12, 1046–1051. [Google Scholar] [CrossRef]
- Lei, J.D. Mechanisms of Resistance in Corn to Maize Dwarf Mosaic Virus. Phytopathology 1986, 76, 1034. [Google Scholar] [CrossRef]
- Nagy, P.D.; Pogany, J. The Dependence of Viral RNA Replication on Co-Opted Host Factors. Nat. Rev. Microbiol. 2012, 10, 137–149. [Google Scholar] [CrossRef]
- Mäkinen, K.; Hafrén, A. Intracellular Coordination of Potyviral RNA Functions in Infection. Front. Plant Sci. 2014, 5, 110. [Google Scholar] [CrossRef]
- Wei, T.; Huang, T.-S.; McNeil, J.; Laliberté, J.-F.; Hong, J.; Nelson, R.S.; Wang, A. Sequential Recruitment of the Endoplasmic Reticulum and Chloroplasts for Plant Potyvirus Replication. J. Virol. 2010, 84, 799–809. [Google Scholar] [CrossRef]
- Schaad, M.C.; Jensen, P.E.; Carrington, J.C. Formation of Plant RNA Virus Replication Complexes on Membranes: Role of an Endoplasmic Reticulum-Targeted Viral Protein. EMBO J. 1997, 16, 4049–4059. [Google Scholar] [CrossRef]
- Kräutler, B. Breakdown of Chlorophyll in Higher Plants—Phyllobilins as Abundant, Yet Hardly Visible Signs of Ripening, Senescence, and Cell Death. Angew. Chem. Int. Ed. 2016, 55, 4882–4907. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S. Chlorophyll Degradation During Senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Tamary, E.; Nevo, R.; Naveh, L.; Levin-Zaidman, S.; Kiss, V.; Savidor, A.; Levin, Y.; Eyal, Y.; Reich, Z.; Adam, Z. Chlorophyll Catabolism Precedes Changes in Chloroplast Structure and Proteome during Leaf Senescence. Plant Direct 2019, 3, e00127. [Google Scholar] [CrossRef]
- Bianchi, M.E. DAMPs, PAMPs and Alarmins: All We Need to Know about Danger. J. Leukoc. Biol. 2007, 81, 1–5. [Google Scholar] [CrossRef]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in Plant Innate Immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s That Spur Autophagy and Immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef]
- Kadota, Y.; Sklenar, J.; Derbyshire, P.; Stransfeld, L.; Asai, S.; Ntoukakis, V.; Jones, J.D.; Shirasu, K.; Menke, F.; Jones, A.; et al. Direct Regulation of the NADPH Oxidase RBOHD by the PRR-Associated Kinase BIK1 during Plant Immunity. Mol. Cell 2014, 54, 43–55. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of Oxidative Stress in Plants: From Classical Chemistry to Cell Biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Clarke, S.F.; Guy, P.L.; Burritt, D.J.; Jameson, P.E. Changes in the Activities of Antioxidant Enzymes in Response to Virus Infection and Hormone Treatment. Physiol. Plant. 2002, 114, 157–164. [Google Scholar] [CrossRef]
- Ellis, J. Proteins as Molecular Chaperones. Nature 1987, 328, 378–379. [Google Scholar] [CrossRef]
- Tyedmers, J.; Mogk, A.; Bukau, B. Cellular Strategies for Controlling Protein Aggregation. Nat. Rev. Mol. Cell Biol. 2010, 11, 777–788. [Google Scholar] [CrossRef]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular Chaperones and Protein Quality Control. Cell 2006, 125, 443–451. [Google Scholar] [CrossRef]
- Verchot, J. Cellular Chaperones and Folding Enzymes Are Vital Contributors to Membrane Bound Replication and Movement Complexes during Plant RNA Virus Infection. Front. Plant Sci. 2012, 3, 275. [Google Scholar] [CrossRef]
- Lu, R.; Malcuit, I.; Moffett, P.; Ruiz, M.T.; Peart, J.; Wu, A.-J.; Rathjen, J.P.; Bendahmane, A.; Day, L.; Baulcombe, D.C. High Throughput Virus-Induced Gene Silencing Implicates Heat Shock Protein 90 in Plant Disease Resistance. EMBO J. 2003, 22, 5690–5699. [Google Scholar] [CrossRef]
- Higgins, C.F. ABC Transporters: From Microorganisms to Man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef]
- Moons, A. Ospdr9, Which Encodes a PDR-Type ABC Transporter, Is Induced by Heavy Metals, Hypoxic Stress and Redox Perturbations in Rice Roots. FEBS Lett. 2003, 553, 370–376. [Google Scholar] [CrossRef]
- Stukkens, Y.; Bultreys, A.; Grec, S.; Trombik, T.; Vanham, D.; Boutry, M. NpPDR1, a Pleiotropic Drug Resistance-Type ATP-Binding Cassette Transporter from Nicotiana plumbaginifolia, Plays a Major Role in Plant Pathogen Defense. Plant Physiol. 2005, 139, 341–352. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, X.; Li, S.; Jiang, C.-Z.; Zhang, Y.; Niu, L. LrABCF1, a GCN-Type ATP-Binding Cassette Transporter from Lilium Regale, Is Involved in Defense Responses against Viral and Fungal Pathogens. Planta 2016, 244, 1185–1199. [Google Scholar] [CrossRef]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC Transporters: The Power to Change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Dean, M.; Hamon, Y.; Chimini, G. The Human ATP-Binding Cassette (ABC) Transporter Superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Klein, I.; Sarkadi, B.; Váradi, A. An Inventory of the Human ABC Proteins. Biochim. Biophys. Acta-Biomembr. 1999, 1461, 237–262. [Google Scholar] [CrossRef]
- Tusnády, G.E.; Sarkadi, B.; Simon, I.; Váradi, A. Membrane Topology of Human ABC Proteins. FEBS Lett. 2006, 580, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Aller, S.G.; Beis, K.; Carpenter, E.P.; Chang, G.; Chen, L.; Dassa, E.; Dean, M.; Van Hoa, F.D.; Ekiert, D.; et al. Structural and Functional Diversity Calls for a New Classification of ABC Transporters. FEBS Lett. 2020, 594, 3767–3775. [Google Scholar] [CrossRef]
- Ellis, J.G.; Lagudah, E.S.; Spielmeyer, W.; Dodds, P.N. The Past, Present and Future of Breeding Rust Resistant Wheat. Front. Plant Sci. 2014, 5, 641. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Kang, J.; Bräunlich, S.; Boni, R.; Chauhan, H.; Selter, L.L.; Robinson, M.D.; Schmid, M.W.; Wiederhold, E.; Hensel, G.; et al. Abscisic Acid Is a Substrate of the ABC Transporter Encoded by the Durable Wheat Disease Resistance Gene Lr34. New Phytol. 2019, 223, 853–866. [Google Scholar] [CrossRef]
- Sucher, J.; Boni, R.; Yang, P.; Rogowsky, P.; Büchner, H.; Kastner, C.; Kumlehn, J.; Krattinger, S.G.; Keller, B. The Durable Wheat Disease Resistance Gene Lr34 Confers Common Rust and Northern Corn Leaf Blight Resistance in Maize. Plant Biotechnol. J. 2017, 15, 489–496. [Google Scholar] [CrossRef]
- Gas-Pascual, E.; Simonovik, B.; Schaller, H.; Bach, T.J. Inhibition of Cycloartenol Synthase (CAS) Function in Tobacco BY-2 Cells. Lipids 2015, 50, 761–772. [Google Scholar] [CrossRef]
- Hartmann, M. Plant Sterols and the Membrane Environment. Trends Plant Sci. 1998, 3, 170–175. [Google Scholar] [CrossRef]
- Tao, Y.; Jiang, L.; Liu, Q.; Zhang, Y.; Zhang, R.; Ingvardsen, C.R.; Frei, U.K.; Wang, B.; Lai, J.; Lübberstedt, T.; et al. Combined Linkage and Association Mapping Reveals Candidates for Scmv1, a Major Locus Involved in Resistance to Sugarcane Mosaic Virus (SCMV) in Maize. BMC Plant Biol. 2013, 13, 162. [Google Scholar] [CrossRef]
- Cao, J.; Lv, Y.; Hou, Z.; Li, X.; Ding, L. Expansion and Evolution of Thaumatin-like Protein (TLP) Gene Family in Six Plants. Plant Growth Regul. 2016, 79, 299–307. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Liang, F.; Zhang, Y.; Ma, L.; Wang, H.; Liu, D. Functional Analysis of a Pathogenesis-Related Thaumatin-like Protein Gene TaLr35PR5 from Wheat Induced by Leaf Rust Fungus. BMC Plant Biol. 2018, 18, 76. [Google Scholar] [CrossRef]
- Cornelissen, B.J.C.; Van Huijsduijnen, R.A.M.H.; Bol, J.F. A Tobacco Mosaic Virus-Induced Tobacco Protein Is Homologous to the Sweet-Tasting Protein Thaumatin. Nature 1986, 321, 531–532. [Google Scholar] [CrossRef]
- Gollop, R.; Farhi, S.; Perl, A. Regulation of the Leucoanthocyanidin Dioxygenase Gene Expression in Vitis Vinifera. Plant Sci. 2001, 161, 579–588. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering Anthocyanin Biosynthesis in Plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef]
- Dufresne, P.J.; Ubalijoro, E.; Fortin, M.G.; Laliberté, J.-F. Arabidopsis thaliana Class II Poly(A)-Binding Proteins Are Required for Efficient Multiplication of Turnip Mosaic Virus. J. Gen. Virol. 2008, 89, 2339–2348. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, H.; Brandizzi, F.; Verchot, J.; Wang, A. The UPR Branch IRE1-BZIP60 in Plants Plays an Essential Role in Viral Infection and Is Complementary to the Only UPR Pathway in Yeast. PLoS Genet. 2015, 11, e1005164. [Google Scholar] [CrossRef]
- Reagan, B.C.; Burch-Smith, T.M. Viruses Reveal the Secrets of Plasmodesmal Cell Biology. Mol. Plant-Microbe Interact. 2020, 33, 26–39. [Google Scholar] [CrossRef]
- Anjanappa, R.B.; Mehta, D.; Okoniewski, M.J.; Szabelska-Berȩsewicz, A.; Gruissem, W.; Vanderschuren, H. Molecular Insights into Cassava Brown Streak Virus Susceptibility and Resistance by Profiling of the Early Host Response. Mol. Plant Pathol. 2018, 19, 476–489. [Google Scholar] [CrossRef]
- Kauss, H. Callose Biosynthesis as a Ca2+-Regulated Process and Possible Relations to the Induction of Other Metabolic Changes. J. Cell Sci. 1985, 1985, 89–103. [Google Scholar] [CrossRef]
- Zavaliev, R.; Ueki, S.; Epel, B.L.; Citovsky, V. Biology of Callose (β-1,3-Glucan) Turnover at Plasmodesmata. Protoplasma 2011, 248, 117–130. [Google Scholar] [CrossRef]
- Levy, A.; Guenoune-Gelbart, D.; Epel, B.L. β-1,3-Glucanases: Plasmodesmal Gate Keepers for Intercellular Communication. Plant Signal. Behav. 2007, 2, 404–407. [Google Scholar] [CrossRef]
- Pastor, V.; Luna, E.; Ton, J.; Cerezo, M.; García-Agustín, P.; Flors, V. Fine Tuning of Reactive Oxygen Species Homeostasis Regulates Primed Immune Responses in Arabidopsis. Mol. Plant-Microbe Interact. 2013, 26, 1334–1344. [Google Scholar] [CrossRef]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the Trans-Frame Potyvirus Protein P3N-PIPO with Host Protein PCaP1 Facilitates Potyvirus Movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef]
- Harries, P.A.; Park, J.-W.; Sasaki, N.; Ballard, K.D.; Maule, A.J.; Nelson, R.S. Differing Requirements for Actin and Myosin by Plant Viruses for Sustained Intercellular Movement. Proc. Natl. Acad. Sci. USA 2009, 106, 17594–17599. [Google Scholar] [CrossRef]
- Amari, K.; Boutant, E.; Hofmann, C.; Schmitt-Keichinger, C.; Fernandez-Calvino, L.; Didier, P.; Lerich, A.; Mutterer, J.; Thomas, C.L.; Heinlein, M.; et al. A Family of Plasmodesmal Proteins with Receptor-Like Properties for Plant Viral Movement Proteins. PLoS Pathog. 2010, 6, e1001119. [Google Scholar] [CrossRef]
- Thomas, C.L.; Bayer, E.M.; Ritzenthaler, C.; Fernandez-Calvino, L.; Maule, A.J. Specific Targeting of a Plasmodesmal Protein Affecting Cell-to-Cell Communication. PLoS Biol. 2008, 6, e7. [Google Scholar] [CrossRef]
- Ridone, P.; Vassalli, M.; Martinac, B. Piezo1 Mechanosensitive Channels: What Are They and Why Are They Important. Biophys. Rev. 2019, 11, 795–805. [Google Scholar] [CrossRef]
- Zhang, Z.; Tong, X.; Liu, S.-Y.; Chai, L.-X.; Zhu, F.-F.; Zhang, X.-P.; Zou, J.-Z.; Wang, X.-B. Genetic Analysis of a Piezo-like Protein Suppressing Systemic Movement of Plant Viruses in Arabidopsis thaliana. Sci. Rep. 2019, 9, 3187. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez Sanchez, P.A.; Babujee, L.; Jaramillo Mesa, H.; Arcibal, E.; Gannon, M.; Halterman, D.; Jahn, M.; Jiang, J.; Rakotondrafara, A.M. Overexpression of a Modified EIF4E Regulates Potato Virus Y Resistance at the Transcriptional Level in Potato. BMC Genom. 2020, 21, 18. [Google Scholar] [CrossRef]
- Contreras-Paredes, C.A.; Silva-Rosales, L.; Daròs, J.-A.; Alejandri-Ramírez, N.D.; Dinkova, T.D. The Absence of Eukaryotic Initiation Factor EIF(Iso)4E Affects the Systemic Spread of a Tobacco Etch Virus Isolate in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2013, 26, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Cristiano, L. The Pseudogenes of Eukaryotic Translation Elongation Factors (EEFs): Role in Cancer and Other Human Diseases. Genes Dis. 2021, 9, 941–958. [Google Scholar] [CrossRef] [PubMed]
- Mateyak, M.K.; Kinzy, T.G. EEF1A: Thinking Outside the Ribosome. J. Biol. Chem. 2010, 285, 21209–21213. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Shine, M.B.; Cui, X.; Chen, X.; Ma, N.; Kachroo, P.; Zhi, H.; Kachroo, A. The Potyviral P3 Protein Targets Eukaryotic Elongation Factor 1A to Promote the Unfolded Protein Response and Viral Pathogenesis. Plant Physiol. 2016, 172, 221–234. [Google Scholar] [CrossRef]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The Many Roles of the Eukaryotic Elongation Factor 1 Complex. Wiley Interdiscip. Rev. RNA 2012, 3, 543–555. [Google Scholar] [CrossRef]
- Cimarelli, A.; Luban, J. Translation Elongation Factor 1-Alpha Interacts Specifically with the Human Immunodeficiency Virus Type 1 Gag Polyprotein. J. Virol. 1999, 73, 5388–5401. [Google Scholar] [CrossRef]
- Blackwell, J.L.; Brinton, M.A. Translation Elongation Factor-1 Alpha Interacts with the 3′ Stem-Loop Region of West Nile Virus Genomic RNA. J. Virol. 1997, 71, 6433–6444. [Google Scholar] [CrossRef]
- Wei, T.; Li, D.; Marcial, D.; Khan, M.; Lin, M.-H.; Snape, N.; Ghildyal, R.; Harrich, D.; Spann, K. The Eukaryotic Elongation Factor 1A Is Critical for Genome Replication of the Paramyxovirus Respiratory Syncytial Virus. PLoS ONE 2014, 9, e114447. [Google Scholar] [CrossRef]
- Dreher, T.W. Role of TRNA-like Structures in Controlling Plant Virus Replication. Virus Res. 2009, 139, 217–229. [Google Scholar] [CrossRef]
- Li, D.; Wei, T.; Abbott, C.M.; Harrich, D. The Unexpected Roles of Eukaryotic Translation Elongation Factors in RNA Virus Replication and Pathogenesis. Microbiol. Mol. Biol. Rev. 2013, 77, 253–266. [Google Scholar] [CrossRef]
- Matsuda, D.; Dreher, T.W. The TRNA-like Structure of Turnip Yellow Mosaic Virus RNA Is a 3′-Translational Enhancer. Virology 2004, 321, 36–46. [Google Scholar] [CrossRef]
- Choi, Y.G.; Dreher, T.W.; Rao, A.L.N. TRNA Elements Mediate the Assembly of an Icosahedral RNA Virus. Proc. Natl. Acad. Sci. USA 2002, 99, 655–660. [Google Scholar] [CrossRef]
- Annamalai, P.; Rao, A.L.N. In Vivo Packaging of Brome Mosaic Virus RNA3, but Not RNAs 1 and 2, Is Dependent on a cis-Acting 3′ TRNA-Like Structure. J. Virol. 2007, 81, 173–181. [Google Scholar] [CrossRef]
- Helderman, T.A.; Deurhof, L.; Bertran, A.; Boeren, S.; Fokkens, L.; Kormelink, R.; Joosten, M.H.A.J.; Prins, M.; van den Burg, H.A. An Isoform of the Eukaryotic Translation Elongation Factor 1A (EEF1a) Acts as a Pro-Viral Factor Required for Tomato Spotted Wilt Virus Disease in Nicotiana benthamiana. Viruses 2021, 13, 2190. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | 2 dpi | 7 dpi | 12 dpi | 17 dpi | 30 dpi |
|---|---|---|---|---|---|
| B73 | 2.2 | 34.3 | 7090.5 | 43,588.7 | 93,975.7 |
| F1 | 5.0 | 0.9 | 57.5 | 0.2 | 32,616.9 |
| CI-RL1 | 1.8 | 1.5 | 0.8 | 0.8 | 72.8 |
| Go Term | ID | V3 Identifier | log2 FC | Description | ||
|---|---|---|---|---|---|---|
| B73 | F1 | CI-RL1 | ||||
| Nucleic acid metabolic process | Zm00001d008223 | GRMZM2G304121 | −1.2 | 3.4 | Eukaryotic translation initiation factor 6 | |
| Zm00001d035823 | GRMZM2G036525 | 2.9 | Tetratricopeptide repeat (TPR)-like superfamily protein | |||
| Zm00001d004415 | GRMZM2G327247 | 1.6 | RNA-dependent RNA polymerase | |||
| Zm00001d026398 | GRMZM2G445575 | −1.3 | 1.4 | Transcription factor TGA4 | ||
| Response to abiotic stimulus | Zm00001d031332 | GRMZM2G024668 | 2.0 | Heat shock protein 90-2 | ||
| Zm00001d027511 | GRMZM2G090568 | −1.3 | −1.2 | 2.0 | Catalase/Catalase isozyme 2 | |
| Zm00001d031325 | GRMZM2G080724 | 1.9 | 25.3 kDa heat shock protein chloroplastic | |||
| Translation | Zm00001d034776 | GRMZM2G040369 | 2.5 | 1.4 | Elongation factor 2 | |
| Regulation of biological process | Zm00001d044242 | GRMZM2G081816 | 2.1 | 1.6 | Transcription factor bHLH87 | |
| Negative regulation of biological process | Zm00001d011096 | GRMZM2G162525 | 1.7 | 1.4 | Argonaute1 | |
| Zm00001d050259 | GRMZM2G091478 | 3.7 | −1.5 | ABC transporter A family member 7 | ||
| Zm00001d043762 | GRMZM5G808836 | 1.9 | 1.3 | −2.8 | ABC transporter B family member 9 | |
| Zm00001d004442 | GRMZM2G167658 | −2.1 | 1.7 | ABC transporter B family member 19 | ||
| Zm00001d008178 | GRMZM2G085111 | 1.0 | ABC transporter B family member 21 | |||
| Zm00001d048329 | GRMZM2G361256 | 1.8 | −1.7 | ABC transporter C family member 3 | ||
| Zm00001d025043 | 1.5 | −1.2 | ABC transporter C family member 4 | |||
| Zm00001d024497 | GRMZM2G165757 | −2.5 | ABC transporter G family member 14 | |||
| Cellular response to stress | Zm00001d053554 | GRMZM2G117706 | 4.3 | Peroxidase/Peroxidase 52 | ||
| Zm00001d052335 | 2.8 | Peroxidase/Peroxidase 67 | ||||
| Zm00001d040705 | GRMZM2G107228 | 2.6 | −2.2 | Peroxidase/Peroxidase 64 | ||
| Zm00001d008173 | GRMZM2G122853 | 1.8 | Peroxidase/Peroxidase 2 | |||
| Zm00001d042022 | GRMZM2G103342 | 1.6 | −1.8 | Peroxidase/Peroxidase 12 | ||
| Zm00001d014341 | GRMZM2G150893 | 1.5 | 1.1 | Peroxidase/Peroxidase 53 | ||
| Zm00001d040364 | GRMZM2G089982 | 1.3 | Peroxidase/Peroxidase 72 | |||
| Zm00001d007161 | GRMZM2G450717 | 1.3 | Peroxidase/Peroxidase 52 | |||
| Zm00001d038599 | GRMZM2G089895 | 1.2 | Peroxidase/Peroxidase 1 | |||
| Zm00001d029604 | GRMZM2G020523 | 1.2 | Peroxidase/Peroxidase 2 | |||
| Response to light stimulus | Zm00001d027511 | GRMZM2G090568 | −1.3 | −1.2 | 2.0 | Catalase/Catalase isozyme 2 |
| Response to chemical | Zm00001d037894 | GRMZM2G079440 | −4.8 | −4.3 | RAB17 protein/Dehydrin DHN1 | |
| Hormone-mediated signaling pathway | Zm00001d023659 | GRMZM2G006042 | −1.1 | Auxin response factor 2 | ||
| Zm00001d018178 | GRMZM2G479760 | −1.0 | ABSCISIC ACID-INSENSITIVE 5-like protein 5 | |||
| Cellular nitrogen compound metabolic process | Zm00001d031953 | GRMZM5G882446 | −1.2 | Glutathione S-transferase | ||
| Response to hormone | Zm00001d014606 | GRMZM2G133434 | −1.2 | Peroxidase/Peroxidase 45 | ||
| Zm00001d020808 | GRMZM5G843748 | −1.1 | −1.6 | Peroxidase/Peroxidase 17 | ||
| Zm00001d009373 | GRMZM2G023840 | −1.5 | Peroxidase/Peroxidase 72 | |||
| Zm00001d037873 | GRMZM2G154218 | 0.8 | 0.9 | Elongation factor 1-alpha (elfa3) | ||
| Zm00001d036904 | GRMZM2G343543 | 0.6 | Elongation factor 1-alpha (elfa10) | |||
| Zm00001d037877 | GRMZM2G001327 | 0.7 | 0.7 | −0.5 | Elongation factor 1-alpha (elfa12) | |
| Zm00001d025100 | GRMZM5G801457 | −0.7 | Translation elongation factor EF1A (Zm-elfa) | |||
| ID | V3 Identifier | log2 FC | Description | ||
|---|---|---|---|---|---|
| B73 | F1 | CI-RL1 | |||
| Zm00001d021205 | GRMZM2G069082 | −2.0 | −0.9 | 2.7 | AP2-EREBP transcription factor/Dehydration-responsive element-binding protein 1B |
| Zm00001d006169 | GRMZM2G124037 | −3.0 | −1.2 | 2.2 | CBF3-like protein; CRT/DRE binding factor; DREB-like protein |
| Zm00001d021208 | GRMZM2G069146 | −2.0 | −0.8 | 2.0 | AP2-EREBP transcription factor/Dehydration-responsive element-binding protein 1B |
| Zm00001d027511 | GRMZM2G090568 | −1.3 | −1.2 | 2.0 | Catalase/Catalase isozyme 2 |
| Zm00001d031745 | GRMZM2G142962 | −1.5 | −0.6 | 1.5 | Homeobox-leucine zipper protein HAT3 |
| Zm00001d016361 | GRMZM2G031983 | −1.3 | 0.9 | 1.5 | C2C2-GATA transcription factor/Putative GATA transcription factor 22 |
| Zm00001d038850 | GRMZM2G111017 | −1.1 | −0.7 | 1.3 | Probable ADPATP carrier protein |
| Zm00001d053911 | GRMZM2G042664 | −1.2 | −0.4 | 1.3 | expressed protein |
| Zm00001d043060 | GRMZM2G059562 | −1.7 | −1.1 | 1.3 | Probable WRKY transcription factor 30 |
| Zm00001d044117 | GRMZM2G150260 | −1.1 | 1.9 | 1.2 | DNA binding protein |
| Zm00001d043505 | GRMZM2G039246 | −1.5 | −1.4 | 1.2 | Histidine-containing phosphotransfer protein 4 |
| Zm00001d033510 | GRMZM2G146616 | −1.5 | −0.5 | 1.2 | expressed protein |
| Zm00001d018107 | AC220927.3_FG007 | −1.3 | −0.9 | 1.2 | Protein EXORDIUM |
| Zm00001d033786 | GRMZM2G155767 | −1.7 | −0.8 | 1.2 | histidine kinase4 |
| Zm00001d016982 | GRMZM2G176253 | −1.4 | −1.2 | 1.2 | Protein NRT1/PTR FAMILY 6.4 |
| Zm00001d025470 | GRMZM2G158097 | −1.2 | −0.5 | 1.2 | expressed protein |
| Zm00001d010490 | GRMZM2G084489 | −1.3 | −0.6 | 1.1 | CW-type Zinc Finger |
| Zm00001d002256 | GRMZM5G897776 | −1.5 | −0.6 | 1.0 | Starch synthase 3 chloroplastic/amyloplastic |
| Zm00001d019217 | GRMZM2G031780 | −1.0 | −0.7 | 1.0 | ABC2 homolog 13 |
| Zm00001d031158 | GRMZM2G402631 | 7.0 | 4.0 | −2.7 | Pathogenesis-related protein5 |
| Zm00001d030548 | GRMZM2G162158 | 6.0 | 2.8 | −2.0 | Leucoanthocyanidin dioxygenase |
| Zm00001d002707 | GRMZM2G120016 | 4.8 | 5.2 | −3.6 | Glycosyltransferase/Cis-zeatin O-glucosyltransferase 1 |
| Zm00001d052690 | AC233883.1_FG006 | 3.8 | 2.2 | −3.4 | UPF0481 protein |
| Zm00001d049632 | GRMZM2G007587 | 3.7 | 1.6 | −1.4 | Kelch motif family protein isoform 1\F-box/kelch-repeat protein |
| Zm00001d044904 | GRMZM2G017244 | 3.6 | 2.5 | −1.5 | Transcription factor EMB1444 |
| Zm00001d002436 | GRMZM2G125669 | 3.2 | 2.8 | −3.6 | Ubiquinol oxidase/alternative oxidase2 |
| Zm00001d048683 | GRMZM2G106377 | 3.2 | 1.6 | −2.1 | Putative RING zinc finger domain superfamily protein |
| Zm00001d052989 | GRMZM2G130389 | 3.1 | 1.4 | −1.1 | Cysteine-rich receptor-like protein kinase 6 |
| Zm00001d024963 | GRMZM2G330635 | 2.9 | 2.1 | −1.7 | Glutathione S-transferase GSTU6 |
| Zm00001d032869 | GRMZM5G896540 | 2.8 | 1.1 | −2.0 | L-type lectin-domain containing receptor kinase V.9 |
| Zm00001d039077 | GRMZM5G846057 | 2.8 | 0.9 | −2.5 | AP2-EREBP transcription factor/Ethylene-responsive transcription factor ERF061 |
| Zm00001d005028 | GRMZM2G018436 | 2.7 | 1.1 | −2.0 | NAC domain-containing protein 77 |
| Zm00001d040743 | GRMZM2G091088 | 2.7 | 1.1 | −1.0 | Os01g23380-like protein |
| Zm00001d044874 | GRMZM2G135960 | 2.6 | 1.4 | −4.0 | 23.6 kDa heat shock protein mitochondrial |
| Zm00001d048949 | GRMZM2G117971 | 2.6 | −2.6 | −1.9 | Hevein-like preproprotein |
| Zm00001d015623 | GRMZM2G417945 | 2.5 | 1.1 | −1.8 | Glycosyltransferase/Limonoid UDP-glucosyltransferase |
| Zm00001d034501 | GRMZM2G137321 | 2.5 | 1.1 | −2.7 | AAA-ATPase ASD mitochondrial |
| Zm00001d038451 | GRMZM2G169966 | 2.4 | 1.0 | −2.7 | Putative WRKY DNA-binding domain superfamily protein |
| C0 | C1 | C2 | C3 | C4 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Transcript ID | Gene | Transcript ID | Gene | Transcript ID | Gene | Transcript ID | Gene | Transcript ID |
| elfa1 | Zm00001d009868_T004 | elfa1 | Zm00001d009868_T009 | elfa1 | Zm00001d009868_T003 | ZmeIF4E | Zm00001d041973_T001 | eif-6 | Zm00001d041682_T001 |
| elfa2 | Zm00001d037875_T005 | elfa2 | Zm00001d037875_T008 | elfa1 | Zm00001d009868_T012 | eif-7 | Zm00001d014065_T001 | elfa2 | Zm00001d037875_T006 |
| elfa3 | Zm00001d037873_T011 | elfa3 | Zm00001d037873_T009 | elfa2 | Zm00001d037875_T003 | elfa1 | Zm00001d009868_T002 | elfa3 | Zm00001d037873_T001 |
| elfa9 | Zm00001d046449_T004 | elfa10 | Zm00001d036904_T002 | elfa11 | Zm00001d037905_T003 | elfa12 | Zm00001d037877_T005 | elfa7 | Zm00001d009870_T013 |
| elfa11 | Zm00001d037905_T002 | elfa10 | Zm00001d036904_T005 | elfa12 | Zm00001d037877_T008 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Gómez, G.; Vargas-Mejía, P.; Silva-Rosales, L. Differential Expression of Genes between a Tolerant and a Susceptible Maize Line in Response to a Sugarcane Mosaic Virus Infection. Viruses 2022, 14, 1803. https://doi.org/10.3390/v14081803
Rodríguez-Gómez G, Vargas-Mejía P, Silva-Rosales L. Differential Expression of Genes between a Tolerant and a Susceptible Maize Line in Response to a Sugarcane Mosaic Virus Infection. Viruses. 2022; 14(8):1803. https://doi.org/10.3390/v14081803
Chicago/Turabian StyleRodríguez-Gómez, Gustavo, Pablo Vargas-Mejía, and Laura Silva-Rosales. 2022. "Differential Expression of Genes between a Tolerant and a Susceptible Maize Line in Response to a Sugarcane Mosaic Virus Infection" Viruses 14, no. 8: 1803. https://doi.org/10.3390/v14081803