
The Effects of Cucumber Mosaic Virus and Its 2a and 2b Proteins on Interactions of Tomato Plants with the Aphid Vectors Myzus persicae and Macrosiphum euphorbiae

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Aphids and Plants
2.2. Virus Preparation, Plant Inoculation, and Virus Detection
2.3. Aphid Free-Choice Settling and Preference Assays and Olfactometry
2.4. Volatile Organic Compound Entrainment and Analysis
3. Results
3.1. CMV Alters the Settling Behaviour of Aphids on Tomato Plants
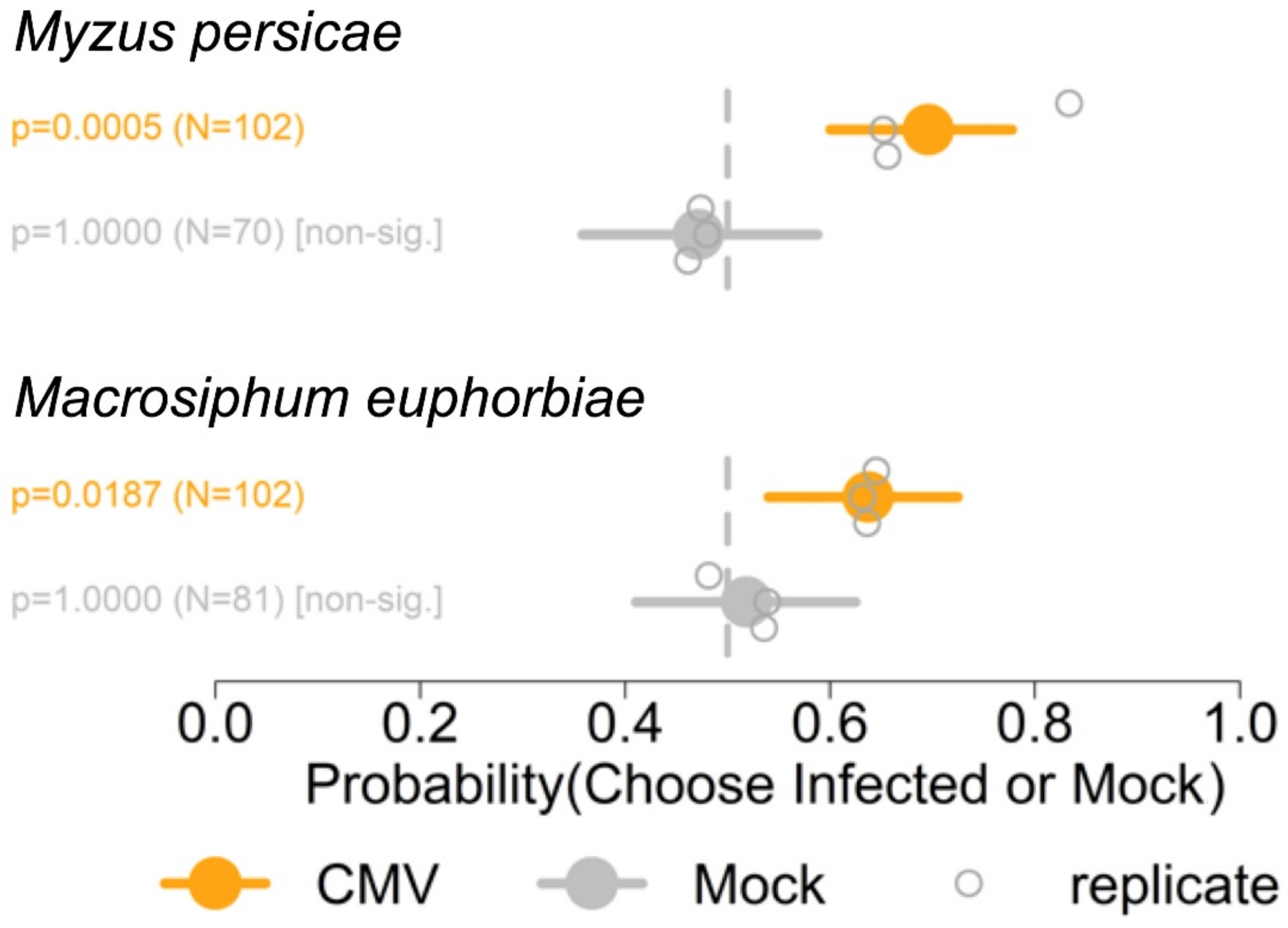
3.2. Myzus persicae and Macrosiphum euphorbiae Preferred Odours Emitted by Plants Infected with Fny-CMV
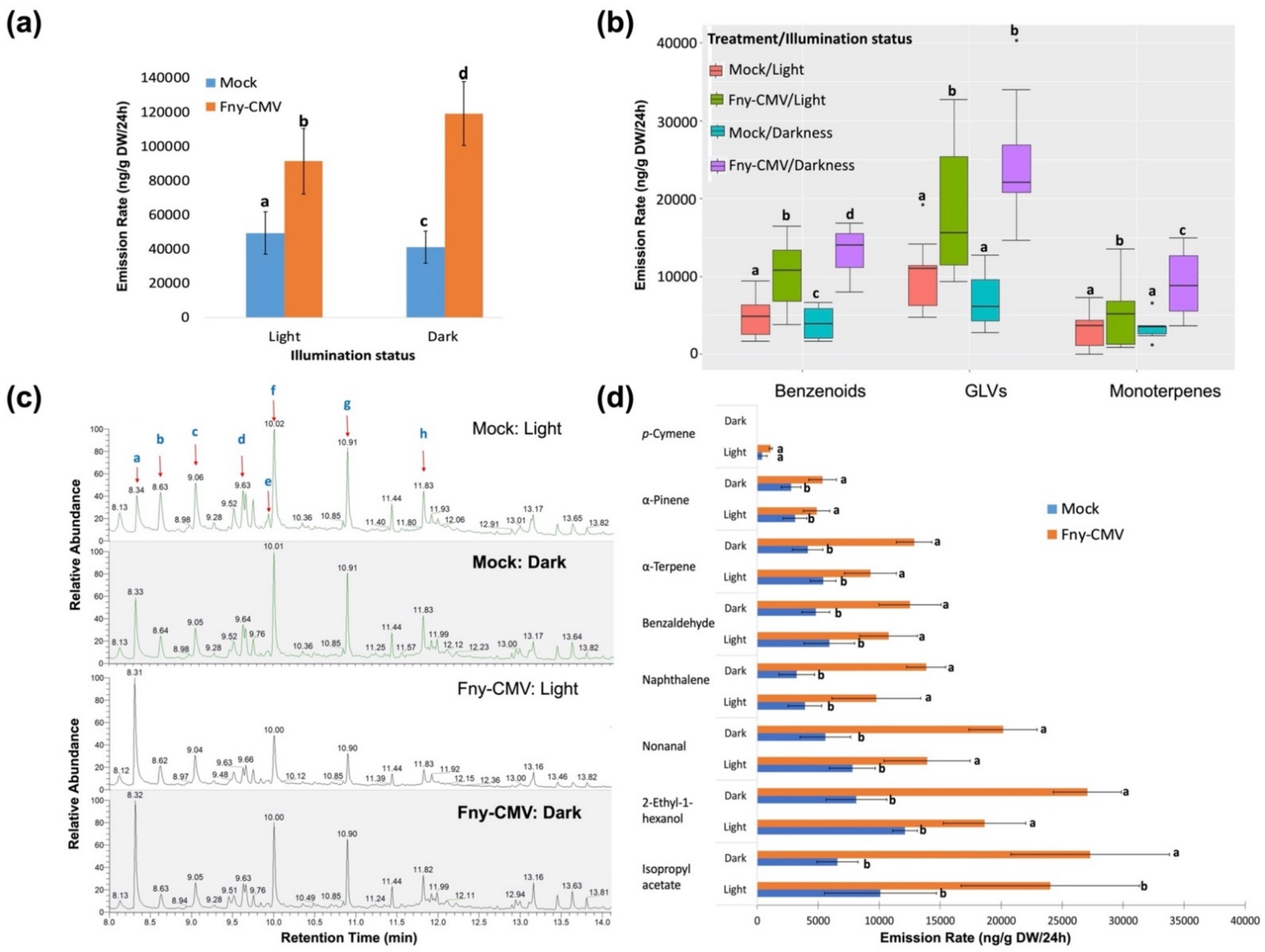
3.3. The Effect of Illumination on CMV-Induced Changes in Plant VOC Emission
3.4. CMV RNA 2 Influences Interactions between Tomato Plants and Aphids
3.5. Investigating the Effect of Salicylic Acid on CMV-Induced Changes in Aphid–Tomato Interactions
4. Discussion
4.1. The Effects of Fny-CMV on Interactions of Tomato with Aphids
4.2. Inter-Strain Differences in CMV-Induced Changes in Aphid–Tomato Interactions and the Role of CMV RNA 2
4.3. To What Cues Are Aphids Responding?
4.4. The Relationship of CMV-Induced Salicylic Acid Accumulation to Aphid Infestation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacquemond, M. Cucumber mosaic virus. Adv. Virus Res. 2012, 84, 439–504. [Google Scholar] [PubMed]
- Palukaitis, P.; García-Arenal, F. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar]
- Ding, S.W.; Anderson, B.J.; Haase, H.R.; Symons, R.H. New overlapping gene encoded by the cucumber mosaic virus genome. Virology 1994, 198, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.N.; Palukaitis, P.; Carr, J.P. Subcellular distribution analysis of the cucumber mosaic virus 2b protein. J. Gen. Virol. 2000, 81, 219–226. [Google Scholar] [CrossRef]
- Ng, J.C.K.; Josefsson, C.; Clark, A.J.; Franz, A.W.E.; Perry, K.L. Virion stability and aphid vector transmissibility of cucumber mosaic virus mutants. Virology 2005, 332, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Fereres, A.; Perry, K.L. Movement between plants: Horizontal transmission. In Cucumber Mosaic Virus; Palukaitis, P., García-Arenal, F., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2019; pp. 173–184. [Google Scholar]
- Suzuki, M.; Kuwata, S.; Kataoka, J.; Masuta, C.; Nitta, N.; Takanami, Y. Functional analysis of deletion mutants of cucumber mosaic virus RNA3 using an in vitro transcription system. Virology 1991, 183, 106–113. [Google Scholar] [CrossRef]
- Canto, T.; Prior, D.A.; Hellwald, K.H.; Oparka, K.J.; Palukaitis, P. Characterization of cucumber mosaic virus. IV. Movement protein and coat protein are both essential for cell-to-cell movement of cucumber mosaic virus. Virology 1997, 237, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.K.; Palukaitis, P.; Min, B.E.; Lee, M.Y.; Choi, J.K.; Ryu, K.H. Cucumber mosaic virus 2a polymerase and 3a movement proteins independently affect both virus movement and the timing of symptom development in zucchini squash. J. Gen. Virol. 2005, 86, 1213–1222. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Palukaitis, P. Cucumber mosaic virus 1a protein interacts with the tobacco she1 transcription factor and partitions between the nucleus and the tonoplast membrane. Plant Pathol. J. 2021, 37, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.A.; Roberts, F.M. A comparative study of the transmission of Hyoscyamus virus 3, potato virus Y and cucumber virus 1 by the vectors Myzus persicae (Sulz), Myzus circumflexus (Buckton), and Macrosiphum gei (Koch). Proc. R. Soc. B 1939, 127, 543–576. [Google Scholar]
- Kennedy, J.S.; Day, M.F.; Eastop, V.F. A Conspectus of Aphids as Vectors of Plant Viruses; Commonwealth Institute of Entomology: London, UK, 1962. [Google Scholar]
- Nalam, V.; Louis, J.; Shah, J. Plant defense against aphids, the pest extraordinaire. Plant Sci. 2019, 279, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Matthews’ Plant Virology, 5th ed.; Academic Press: London, UK, 2013. [Google Scholar]
- Liang, Y.; Gao, X.W. The cuticle protein gene MPCP4 of Myzus persicae (Homoptera: Aphididae) plays a critical role in cucumber mosaic virus acquisition. J. Econ. Entomol. 2017, 110, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Deshoux, M.; Masson, V.; Arafah, K.; Voisin, S.; Guschinskaya, N.; van Munster, M.; Cayrol, B.; Webster, C.G.; Rahbé, Y.; Blanc, S.; et al. Cuticular structure proteomics in the pea aphid Acyrthosiphon pisum reveals new plant virus receptor candidates at the tip of maxillary stylets. J. Proteome Res. 2020, 19, 1319–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenz, B.; Bronikowski, A.; Lu, X.; Ziebell, H.; Thompson, J.R.; Perry, K.L. Visual monitoring of cucumber mosaic virus infection in Nicotiana benthamiana following transmission by the aphid vector Myzus persicae. J. Gen. Virol. 2015, 96, 2904–2912. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, R.; Cunniffe, N.J.; Carr, J.P.; Gilligan, C.A. Pathogenic modification of plants enhances long-distance dispersal of non-persistently transmitted viruses to new hosts. Ecology 2019, 100, e02725. [Google Scholar] [CrossRef] [Green Version]
- Cunniffe, N.J.; Taylor, N.P.; Hamelin, F.M.; Jeger, M.J. Epidemiological and ecological consequences of virus manipulation of host and vector in plant virus transmission. PLoS Comput. Biol. 2021, 17, e1009759. [Google Scholar] [CrossRef] [PubMed]
- Castle, S.J.; Berger, P.H. Rates of growth and increase of Myzus persicae on virus-infected potatoes according to type of virus-vector relationship. Entomol. Exp. Appl. 1993, 69, 51–60. [Google Scholar] [CrossRef]
- Hodge, S.; Powell, G. Do plant viruses facilitate their aphid vectors by inducing symptoms that alter behaviour and performance? Environ. Entomol. 2008, 37, 1573–1581. [Google Scholar] [CrossRef]
- Ziebell, H.; Murphy, A.M.; Groen, S.C.; Tungadi, T.; Westwood, J.H.; Lewsey, M.G.; Moulin, M.; Kleczkowski, A.; Smith, A.G.; Stevens, M.; et al. Cucumber mosaic virus and its 2b RNA silencing suppressor modify plant-aphid interactions in tobacco. Sci. Rep. 2011, 1, 187. [Google Scholar] [CrossRef]
- Mauck, K.; Bosque-Pérez, N.A.; Eigenbrode, S.D.; De Moraes, C.M.; Mescher, M.C. Transmission mechanisms shape pathogen effects on host-vector interactions: Evidence from plant viruses. Funct. Ecol. 2012, 26, 1162–1175. [Google Scholar] [CrossRef]
- Westwood, J.H.; Groen, S.C.; Du, Z.; Murphy, A.M.; Anggoro, D.T.; Tungadi, T.; Luang-In, V.; Lewsey, M.G.; Rossiter, J.T.; Powell, G.; et al. A trio of viral proteins tunes aphid-plant interactions in Arabidopsis thaliana. PLoS ONE 2013, 8, e0083066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H.; Lewsey, M.G.; Murphy, A.M.; Tungadi, T.; Bates, A.; Gilligan, C.A.; Carr, J.P. Interference with jasmonic acid-regulated gene expression is a general property of viral suppressors of RNA silencing but only partly explains virus-induced changes in plant-aphid interactions. J. Gen. Virol. 2014, 95, 733–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteel, C.L.; Yang, C.; Nanduri, A.C.; De Jong, H.N.; Whitham, S.A.; Jander, G. The NIa-Pro protein of Turnip mosaic virus improves growth and reproduction of the aphid vector, Myzus persicae (green peach aphid). Plant J. 2014, 77, 653–663. [Google Scholar] [CrossRef]
- Carr, J.P.; Tungadi, T.; Donnelly, R.; Bravo-Cazar, A.; Rhee, S.J.; Watt, L.G.; Mutuku, J.M.; Wamonje, F.O.; Murphy, A.M.; Arinaitwe, W.; et al. Modelling and manipulation of aphid-mediated spread of non-persistently transmitted viruses. Virus Res. 2020, 277, 197845. [Google Scholar] [CrossRef] [PubMed]
- Wamonje, F.O.; Donnelly, R.; Tungadi, T.D.; Murphy, A.M.; Pate, A.E.; Woodcock, C.; Caulfield, J.; Mutuku, J.M.; Bruce, T.; Gilligan, C.A.; et al. Different plant viruses induce changes in feeding behavior of specialist and generalist aphids on common bean that are likely to enhance virus transmission. Front. Plant Sci. 2020, 10, 1811. [Google Scholar] [CrossRef] [Green Version]
- Wamonje, F.O.; Tungadi, T.D.; Murphy, A.M.; Pate, A.E.; Woodcock, C.; Caulfield, J.C.; Mutuku, J.M.; Cunniffe, N.J.; Bruce, T.J.A.; Gilligan, C.A.; et al. Three aphid-transmitted viruses encourage vector migration from infected common bean (Phaseolus vulgaris) plants through a combination of volatile and surface cues. Front. Plant Sci. 2020, 11, 613772. [Google Scholar] [CrossRef]
- Jayasinghe, W.H.; Akhter, M.S.; Nakahara, K.; Maruthi, M.N. Effect of aphid biology and morphology on plant virus transmission. Pest Manag. Sci. 2022, 78, 416–427. [Google Scholar] [CrossRef]
- Ray, S.; Casteel, C.L. Effector-mediated plant-virus-vector interactions. Plant Cell 2022, 34, 1514–1531. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. USA 2010, 107, 3600–3605. [Google Scholar] [CrossRef] [Green Version]
- Tungadi, T.; Donnelly, R.; Qing, L.; Iqbal, J.; Murphy, A.M.; Pate, A.E.; Cunniffe, N.J.; Carr, J.P. Cucumber mosaic virus 2b proteins inhibit virus-induced aphid resistance in tobacco. Mol. Plant Pathol. 2020, 21, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Carmo-Sousa, M.; Moreno, A.; Garzo, E.; Fereres, A. A non-persistently transmitted-virus induces a pull-push strategy in its aphid vector to optimise transmission and spread. Virus Res. 2014, 186, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Tungadi, T.; Groen, S.C.; Murphy, A.M.; Pate, A.E.; Iqbal, J.; Bruce, T.J.; Cunniffe, N.J.; Carr, J.P. Cucumber mosaic virus and its 2b protein alter emission of host volatile organic compounds but not aphid vector settling in tobacco. Virol. J. 2017, 14, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tungadi, T.; Watt, L.G.; Groen, S.C.; Murphy, A.M.; Du, Z.; Pate, A.E.; Westwood, J.H.; Fennell, T.G.; Powell, G.; Carr, J.P. Infection of Arabidopsis by cucumber mosaic virus triggers jasmonate-dependent resistance to aphids that relies partly on the pattern-triggered immunity factor BAK1. Mol. Plant Pathol. 2021, 22, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Watt, L.G.; Crawshaw, S.; Rhee, S.J.; Murphy, A.M.; Canto, T.; Carr, J.P. The cucumber mosaic virus 1a protein regulates interactions between the 2b protein and ARGONAUTE 1 while maintaining the silencing suppressor activity of the 2b protein. PLoS Pathog. 2020, 16, e1009125. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.J.; Watt, L.G.; Bravo-Cazar, A.; Murphy, A.M.; Carr, J.P. Effects of the cucumber mosaic virus 2a protein on aphid–plant interactions in Arabidopsis thaliana. Mol. Plant Pathol. 2020, 21, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Groen, S.C.; Jiang, S.; Murphy, A.M.; Cunniffe, N.J.; Westwood, J.H.; Davey, M.P.; Bruce, T.J.A.; Caulfield, J.C.; Furzer, O.J.; Reed, A.; et al. Virus infection of plants alters pollinator preference: A payback for susceptible hosts? PLoS Pathog. 2016, 12, e1005790. [Google Scholar] [CrossRef] [Green Version]
- Mhlanga, N.M.; Murphy, A.M.; Wamonje, F.O.; Cunniffe, N.J.; Caulfield, J.C.; Glover, B.J.; Carr, J.P. An innate preference of bumblebees for volatile organic compounds emitted by Phaseolus vulgaris plants infected with three different viruses. Front. Ecol. Evol. 2021, 9, 626851. [Google Scholar] [CrossRef]
- Devonshire, A.L.; Sawicki, R.M. Insecticide-resistant Myzus persicae as an example of evolution by gene duplication. Nature 1979, 280, 140–141. [Google Scholar] [CrossRef]
- Sobhy, I.S.; Woodcock, C.M.; Powers, S.J.; Caulfield, J.C.; Pickett, J.A.; Birkett, M.A. cis-Jasmone elicits aphid-induced stress signalling in potatoes. J. Chem. Ecol. 2017, 43, 39–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brading, P.A.; Hammond-Kosack, K.E.; Parr, A.; Jones, J.D.G. Salicylic acid is not required for Cf-2- and Cf-9-dependent resistance of tomato to Cladosporium fulvum. Plant J. 2000, 23, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hanada, K.; Palukaitis, P. Mapping local and systemic symptom determinants of Cucumber mosaic cucumovirus in tobacco. J. Gen. Virol. 1994, 75, 3185–3191. [Google Scholar] [CrossRef]
- Rizzo, T.M.; Palukaitis, P. Construction of full-length cDNA clones of cucumber mosaic virus RNAs 1, 2 and 3: Generation of infectious RNA transcripts. Mol. Gen. Genet. 1990, 222, 249–256. [Google Scholar] [CrossRef]
- Pita, J.; Roossinck, M.J. Fixation of emerging interviral recombinants in Cucumber mosaic virus populations. J. Virol. 2014, 87, 1264–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soards, A.J.; Murphy, A.M.; Palukaitis, P.; Carr, J.P. Virulence and differential local and systemic spread of cucumber mosaic virus in tobacco are affected by the CMV 2b protein. Mol. Plant-Microbe Interact. 2002, 15, 647–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, S.J.; Zhang, C.; Long, V.; Roossinck, M.J.; Koh, S.H.; Jones, M.G.K.; Iqbal, S.; Li, H. Differential responses to virus challenge of laboratory and wild accessions of Australian species of Nicotiana, and comparative analysis of RDR1 gene sequences. PLoS ONE 2015, 10, e0121787. [Google Scholar] [CrossRef] [PubMed]
- Schwach, F.; Vaistij, F.E.; Jones, L.; Baulcombe, D.C. An RNA-dependent RNA polymerase prevents meristem invasion by potato virus X and is required for the activity but not the production of a systemic silencing signal. Plant. Physiol. 2015, 138, 1842–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palukaitis, P. Virus purification and RNA isolation. In Cucumber Mosaic Virus; Palukaitis, P., García-Arenal, F., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2019; pp. 201–225. [Google Scholar]
- Hu, J.S. Comparison of dot blot, ELISA, and RT-PCR Assays for detection of two cucumber mosaic virus isolates infecting banana in Hawaii. Plant Dis. 1995, 79, 903. [Google Scholar] [CrossRef]
- R Core Team. Available online: https://intro2r.com/citing-r.html (accessed on 13 June 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Beale, M.H.; Birkett, M.A.; Bruce, T.J.A.; Chamberlain, K.; Field, L.M.; Huttly, A.K.; Martin, J.L.; Parker, R.; Phillips, A.L.; Pickett, J.A.; et al. Aphid alarm pheromone produced by transgenic plants affects aphid and parasitoid behaviour. Proc. Natl. Acad. Sci. USA 2006, 103, 10509–10513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesley, M.M. The “wiry” tomato: A recessive mutant form resembling a plant affected with mosaic disease. J. Hered. 1928, 19, 337–344. [Google Scholar]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cillo, F.; Mascia, T.; Pasciuto, M.M.; Gallitelli, D. Differential effects of mild and severe Cucumber mosaic virus strains in the perturbation of MicroRNA-regulated gene expression in tomato map to the 3’ sequence of RNA 2. Mol. Plant-Microbe Interact. 2009, 22, 1239–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewsey, M.G.; Murphy, A.M.; Maclean, D.; Dalchau, N.; Westwood, J.H.; Macaulay, K.; Bennett, M.H.; Moulin, M.; Hanke, D.E.; Powell, G.; et al. Disruption of two defensive signalling pathways by a viral RNA silencing suppressor. Mol. Plant-Microbe Interact. 2010, 23, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Murphy, A.M.; Lewsey, M.G.; Westwood, J.H.; Zhang, H.M.; González, I.; Canto, T.; Carr, J.P. Domains of the cucumber mosaic virus 2b silencing suppressor protein affecting inhibition of salicylic acid-induced resistance and priming of salicylic acid accumulation during infection. J. Gen. Virol. 2014, 95, 1408–1413. [Google Scholar] [CrossRef] [Green Version]
- López-Gresa, M.P.; Lisón, P.; Yenush, L.; Conejero, V.; Rodrigo, I.; Bellés, J.M. Salicylic acid is involved in the basal resistance of tomato plants to citrus exocortis viroid and tomato spotted wilt virus. PLoS ONE 2016, 11, e0166938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, I.; Martínez, L.; Rakinita, D.V.; Lewsey, M.G.; Atienzo, F.A.; Llave, C.; Kalinina, N.O.; Carr, J.P.; Palukaitis, P.; Canto, T. Cucumber mosaic virus 2b protein subcellular targets and interactions: Their significance to RNA silencing suppressor activity. Mol. Plant-Microbe Interact. 2010, 23, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, I.; Rakitina, D.; Semashko, M.; Praveen, S.; Taliansky, M.; Palukaitis, P.; Carr, J.P.; Kalinina, N.; Canto, T. RNA binding is more critical to the suppression of silencing function of Cucumber mosaic virus 2b protein than nuclear localization. RNA 2012, 18, 771–782. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yang, J.; Zhang, X.; Chen, A.; Gu, Z.; Du, Z. The weak small RNA-binding activity of the 2b proteins of subgroup II cucumber mosaic virus strains is insufficient for RNA silencing suppression. Front. Microbiol. 2021, 12, 760937. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Qi, T.; Li, W.X.; Tian, H.; Gao, H.; Wang, J.; Ge, J.; Yao, R.; Ren, C.; Wang, X.B.; et al. Viral effector protein manipulates host hormone signalling to attract insect vectors. Cell Res. 2017, 27, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, W.H.; Kim, H.; Nakada, Y.; Masuta, C. A plant virus satellite RNA directly accelerates wing formation in its insect vector for spread. Nat. Commun. 2021, 12, 7087. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, B.O.; Dewar, A.M. The effect of barley yellow dwarf virus on field populations of the cereal aphids, Sitobion avenae and Metopolophium dirhodum. Ann. Appl. Biol. 1983, 103, 1–11. [Google Scholar] [CrossRef]
- Shi, X.; Gao, Y.; Yan, S.; Tang, X.; Zhou, X.; Zhang, D.; Liu, Y. Aphid performance changes with plant defence mediated by cucumber mosaic virus titer. Virol. J. 2016, 13, 70. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Deng, J.; Zhang, Z.; Yan, S.; Zheng, L.; Sun, S.; Gao, Y.; Zhou, X.; Zhang, D.; Liu, Y. Initial ingestion of CMV-infected plants reduces subsequent aphid performance. Arthropod-Plant Interact. 2021, 15, 153–160. [Google Scholar] [CrossRef]
- Ji, L.H.; Ding, S.W. The suppressor of transgene RNA silencing encoded by Cucumber mosaic virus interferes with salicylic acid-mediated virus resistance. Mol. Plant-Microbe Interact. 2001, 14, 715–724. [Google Scholar] [CrossRef] [Green Version]
- Whitham, S.A.; Quan, S.; Chang, H.S.; Cooper, B.; Estes, B.; Zhu, T.; Wang, X.; Hou, Y.M. Diverse RNA viruses elicit the expression of common sets of genes in susceptible Arabidopsis thaliana plants. Plant J. 2003, 33, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Krečič-Stres, H.; Vučak, C.; Ravnikar, M.; Kovač, M. Systemic Potato virus YNTN infection and levels of salicylic and gentisic acids in different potato genotypes. Plant Pathol. 2005, 54, 441–447. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef]
- Naylor, M.; Murphy, A.M.; Berry, J.O.; Carr, J.P. Salicylic acid can induce resistance to plant virus movement. Mol. Plant-Microbe Interact. 1998, 11, 860–868. [Google Scholar] [CrossRef] [Green Version]
- Sabir Tariq, R.M.; Akhtar, K.P.; Hameed, A.; Ullah, N.; Saleem, N.Y.; Ul Haq, I. Determination of the role of salicylic acid and Benzothiadiazole on physico-chemical alterations caused by Cucumber mosaic virus in tomato. Eur. J. Plant Pathol. 2018, 150, 911–922. [Google Scholar] [CrossRef]
- Murphy, A.M.; Zhou, T.; Carr, J.P. An update on salicylic acid biosynthesis, its induction and its potential exploitation by plant viruses. Curr. Opin. Virol. 2020, 42, 8–17. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arinaitwe, W.; Guyon, A.; Tungadi, T.D.; Cunniffe, N.J.; Rhee, S.-J.; Khalaf, A.; Mhlanga, N.M.; Pate, A.E.; Murphy, A.M.; Carr, J.P. The Effects of Cucumber Mosaic Virus and Its 2a and 2b Proteins on Interactions of Tomato Plants with the Aphid Vectors Myzus persicae and Macrosiphum euphorbiae. Viruses 2022, 14, 1703. https://doi.org/10.3390/v14081703
Arinaitwe W, Guyon A, Tungadi TD, Cunniffe NJ, Rhee S-J, Khalaf A, Mhlanga NM, Pate AE, Murphy AM, Carr JP. The Effects of Cucumber Mosaic Virus and Its 2a and 2b Proteins on Interactions of Tomato Plants with the Aphid Vectors Myzus persicae and Macrosiphum euphorbiae. Viruses. 2022; 14(8):1703. https://doi.org/10.3390/v14081703
Chicago/Turabian StyleArinaitwe, Warren, Alex Guyon, Trisna D. Tungadi, Nik J. Cunniffe, Sun-Ju Rhee, Amjad Khalaf, Netsai M. Mhlanga, Adrienne E. Pate, Alex M. Murphy, and John P. Carr. 2022. "The Effects of Cucumber Mosaic Virus and Its 2a and 2b Proteins on Interactions of Tomato Plants with the Aphid Vectors Myzus persicae and Macrosiphum euphorbiae" Viruses 14, no. 8: 1703. https://doi.org/10.3390/v14081703