
The P3N-PIPO Protein Encoded by Wheat Yellow Mosaic Virus Is a Pathogenicity Determinant and Promotes Its Pathogenicity through Interaction with NbRLK6 in Nicotiana benthamiana
 , , , , and
, , , , and 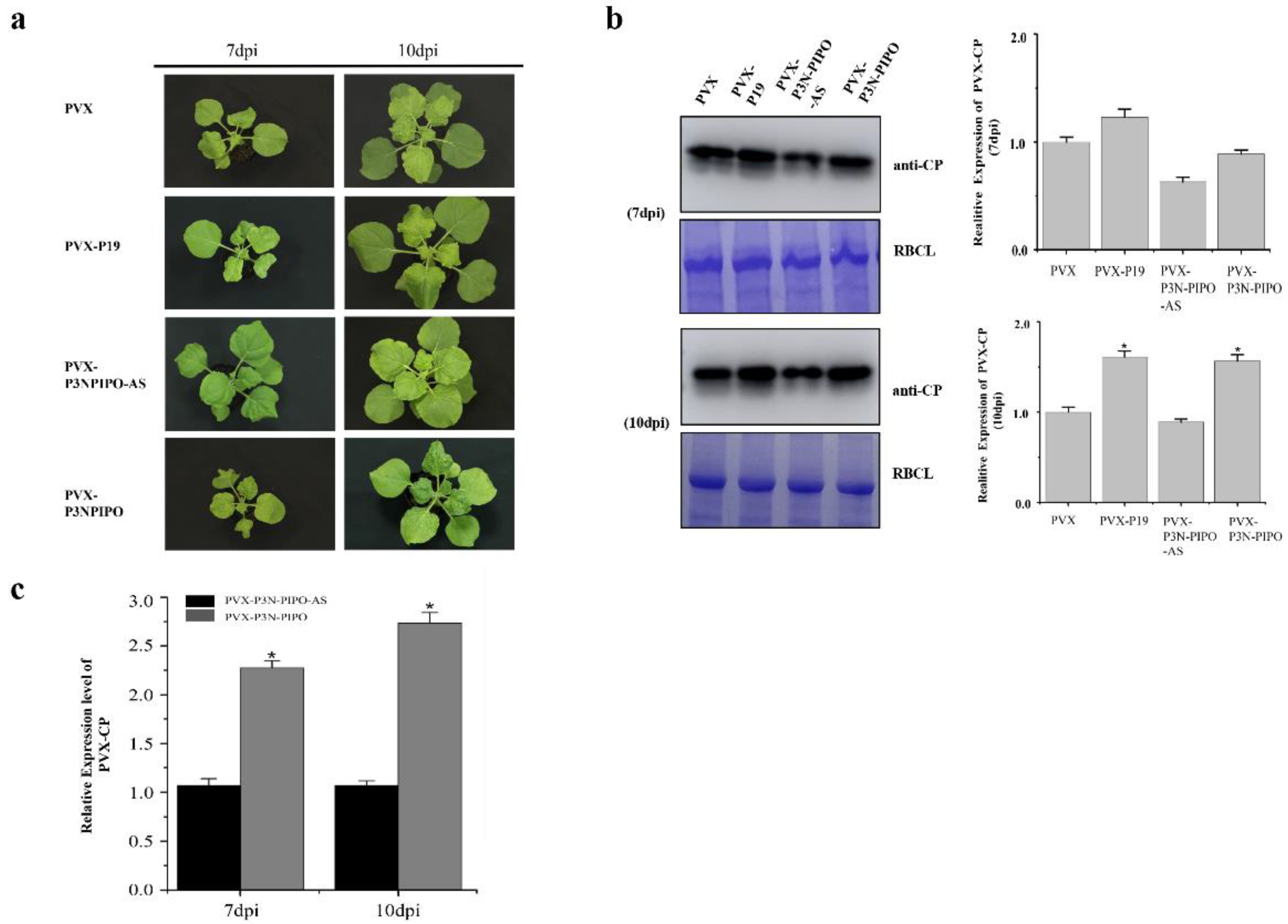{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Plant Growth and Agrobacterium Infection of Plants
2.3. Co-IP/MS
2.4. Bioinformatics Analysis
2.5. BiFC Assay
2.6. Co-IP Transient Assay
2.7. Virus-Induced Gene Silencing in N. benthamiana
2.8. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.9. Western Blot (WB) Analysis
3. Results
3.1. P3N-PIPO Enhanced the Pathogenicity of PVX in N. benthamiana
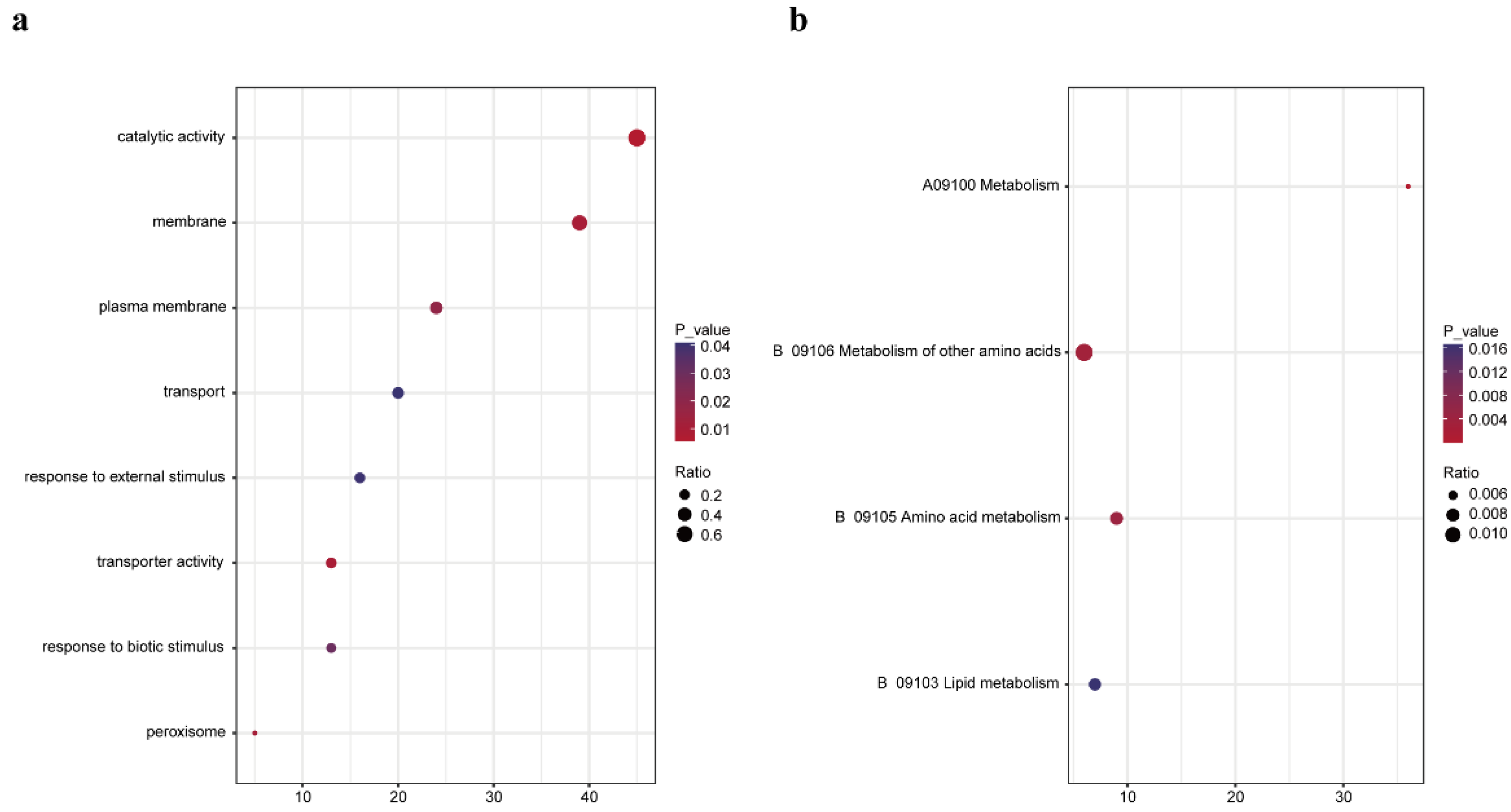
3.2. Identification and Analysis of Plant Proteins Interacting with WYMV P3N-PIPO
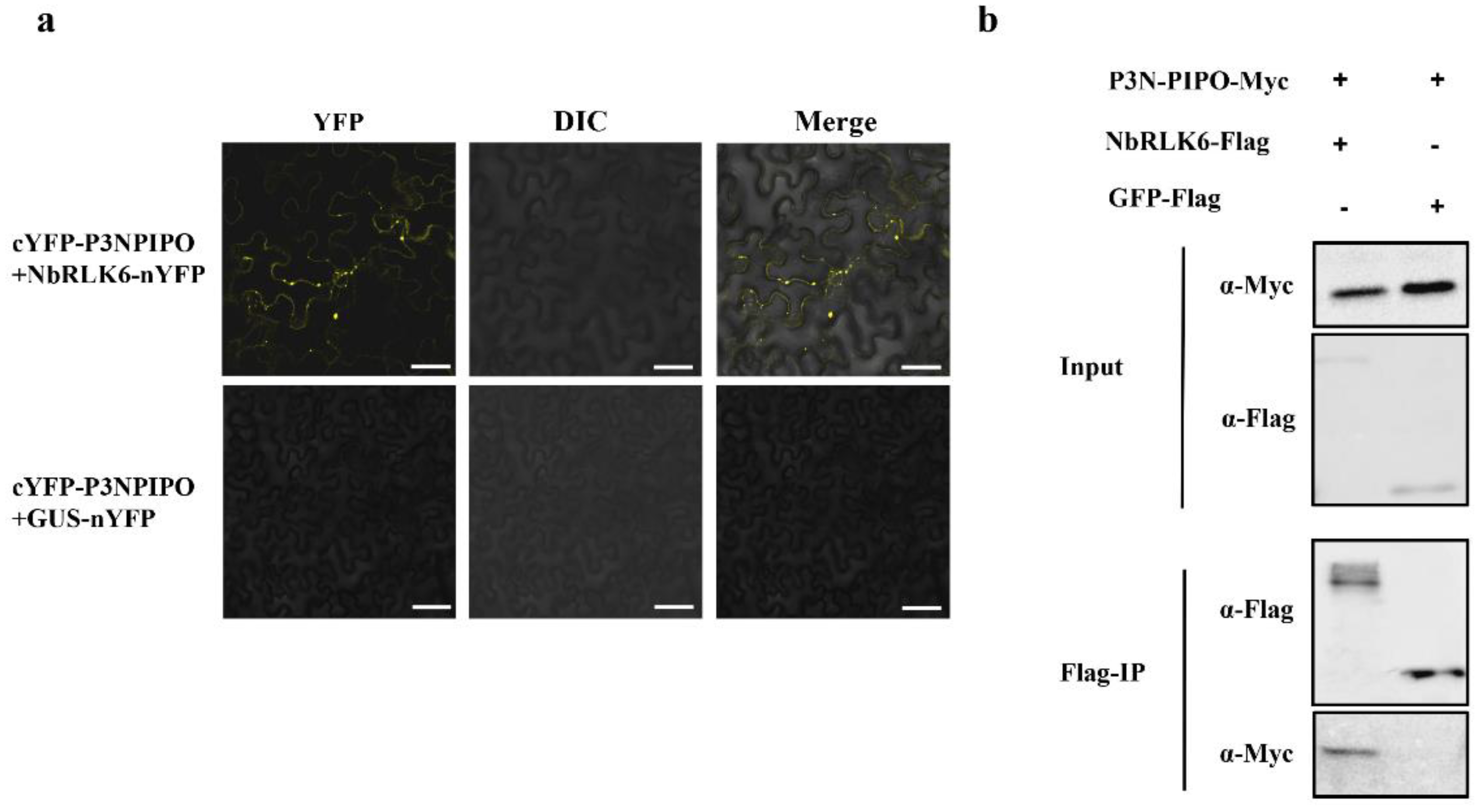
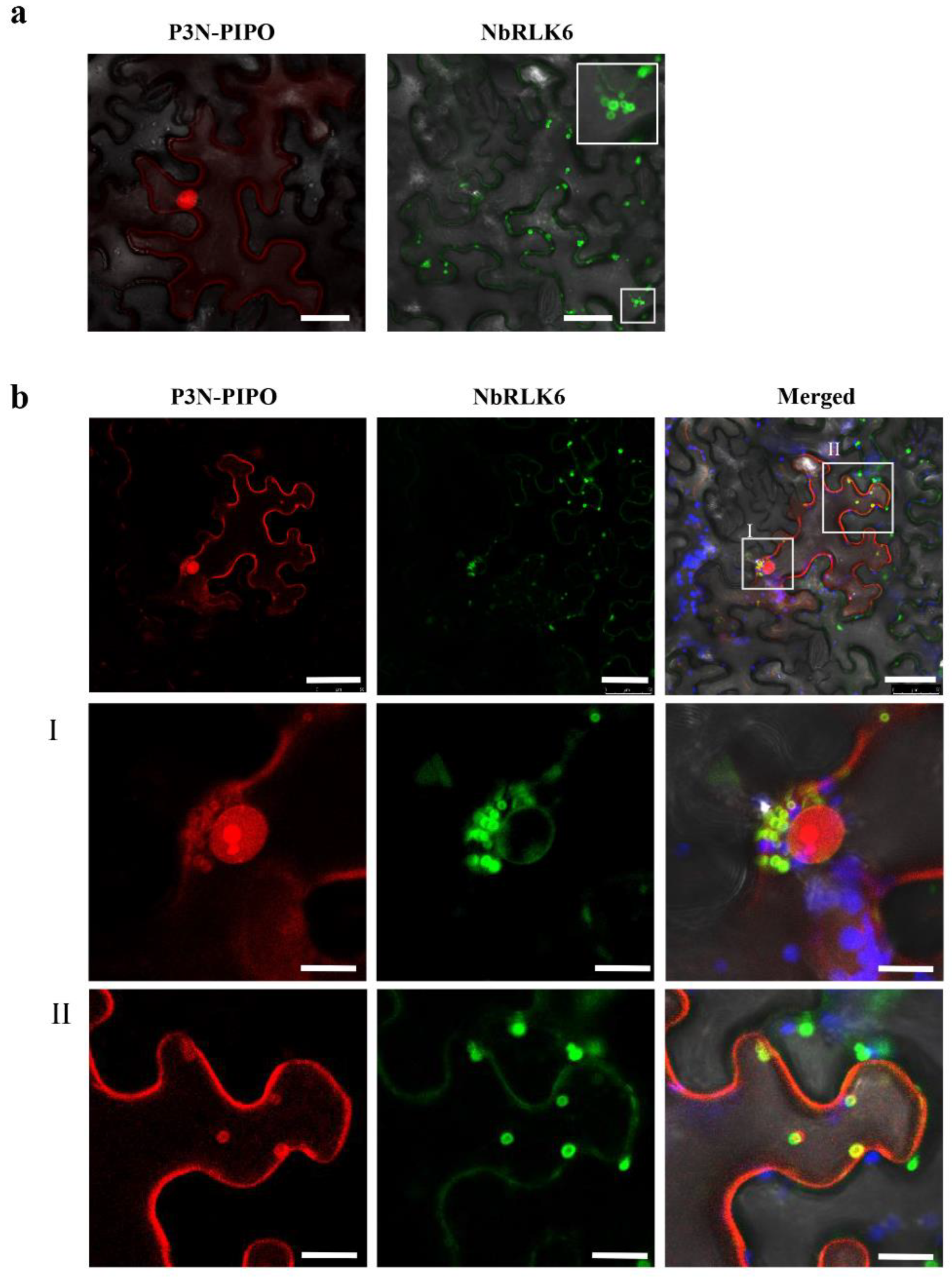
3.3. P3N-PIPO Interacts with NbRLK6
3.4. Silencing of NbRLK6 Expression in N. benthamiana Decreases PVX Accumulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Namba, S.; Kashiwazaki, S.; Lu, X.; Tamura, M.; Tsuchizaki, T. Complete Nucleotide Sequence of Wheat Yellow Mosaic Bymovirus Genomic RNAs. Arch. Virol. 1998, 143, 631–643. [Google Scholar] [CrossRef]
- Yamashita, Y.; Souma, C.; Ogura, R.; Suzuki, T. A Single QTL on Chromosome 6DS Derived from a Winter Wheat Cultivar ‘OW104’ Confers Resistance to Wheat Yellow Mosaic Virus. Breed. Sci. 2020, 70, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.Y.-W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An Overlapping Essential Gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef]
- Olspert, A.; Chung, B.Y.; Atkins, J.F.; Carr, J.P.; Firth, A.E. Transcriptional Slippage in the Positive-Sense RNA Virus Family Potyviridae. EMBO Rep. 2015, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Rodamilans, B.; Valli, A.; Mingot, A.; San León, D.; Baulcombe, D.; López-Moya, J.J.; García, J.A. RNA Polymerase Slippage as a Mechanism for the Production of Frameshift Gene Products in Plant Viruses of the Potyviridae Family. J. Virol. 2015, 89, 6965–6967. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Miao, R.; Ye, Z.; MacFarlane, S.; Lu, Y.; Li, J.; Yang, J.; Yan, F.; Dai, L.; Chen, J. Integrated Proteomics and Transcriptomics Analyses Reveal the Transcriptional Slippage of a Bymovirus P3N-PIPO Gene Expressed from a PVX Vector in Nicotiana Benthamiana. Viruses 2021, 13, 1247. [Google Scholar] [CrossRef] [PubMed]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the Trans-Frame Potyvirus Protein P3N-PIPO with Host Protein PCaP1 Facilitates Potyvirus Movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Zhang, C.; Hong, J.; Xiong, R.; Kasschau, K.D.; Zhou, X.; Carrington, J.C.; Wang, A. Formation of Complexes at Plasmodesmata for Potyvirus Intercellular Movement Is Mediated by the Viral Protein P3N-PIPO. PLoS Pathog. 2010, 6, e1000962. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.-H.; Hajimorad, M.R. Mutational Analysis of the Putative Pipo of Soybean Mosaic Virus Suggests Disruption of PIPO Protein Impedes Movement. Virology 2010, 400, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Cong, Q.-Q.; Li, X.-D.; Mou, A.-L.; Gao, R.; Liu, J.-L.; Tian, Y.-P. DEVELOPMENTALLY REGULATED PLASMA MEMBRANE PROTEIN of Nicotiana Benthamiana Contributes to Potyvirus Movement and Transports to Plasmodesmata via the Early Secretory Pathway and the Actomyosin System. Plant Physiol. 2015, 167, 394–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, M.; Wu, X.; Liu, J.; Fang, Y.; Luan, Y.; Cui, X.; Zhou, X.; Wang, A.; Cheng, X. P3N-PIPO Interacts with P3 via the Shared N-Terminal Domain to Recruit Viral Replication Vesicles for Cell-to-Cell Movement. J. Virol. 2020, 94, e01898-19. [Google Scholar] [CrossRef]
- Hayward, A.; Padmanabhan, M.; Dinesh-Kumar, S.P. Virus-Induced Gene Silencing in Nicotiana Benthamiana and Other Plant Species. Methods Mol. Biol. 2011, 678, 55–63. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, X.; Ge, C.; Ma, Z.; Jiang, M.; Li, T.; Koiwa, H.; Yang, S.W.; Zhang, X. KETCH1 Imports HYL1 to Nucleus for MiRNA Biogenesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 4011–4016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De Novo Sequencing Assisted Database Search for Sensitive and Accurate Peptide Identification. Mol. Cell. Proteom. 2012, 11, M111.010587. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; Kaschani, F.; Grosse-Holz, F.M.; Homma, F.; Kaiser, M.; van der Hoorn, R.A.L. A Homology-Guided, Genome-Based Proteome for Improved Proteomics in the Alloploid Nicotiana Benthamiana. BMC Genom. 2019, 20, 722. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, H.; Chen, C.; Huang, H.; Tan, X.; Wei, Z.; Li, J.; Yan, F.; Zhang, C.; Chen, J.; et al. A Class of Independently Evolved Transcriptional Repressors in Plant RNA Viruses Facilitates Viral Infection and Vector Feeding. Proc. Natl. Acad. Sci. USA 2021, 118, e2016673118. [Google Scholar] [CrossRef]
- Li, S.; Zhao, J.; Zhai, Y.; Yuan, Q.; Zhang, H.; Wu, X.; Lu, Y.; Peng, J.; Sun, Z.; Lin, L.; et al. The Hypersensitive Induced Reaction 3 (HIR3) Gene Contributes to Plant Basal Resistance via an EDS1 and Salicylic Acid-Dependent Pathway. Plant J. 2019, 98, 783–797. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.; Kavanagh, T.; Baulcombe, D. Potato Virus X as a Vector for Gene Expression in Plants. Plant J. 1992, 2, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Dickmeis, C.; Fischer, R.; Commandeur, U. Potato Virus X-Based Expression Vectors Are Stabilized for Long-Term Production of Proteins and Larger Inserts. Biotechnol. J. 2014, 9, 1369–1379. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef]
- Breiden, M.; Simon, R. Q&A: How Does Peptide Signaling Direct Plant Development? BMC Biol. 2016, 14, 58. [Google Scholar] [CrossRef]
- Ye, Y.; Ding, Y.; Jiang, Q.; Wang, F.; Sun, J.; Zhu, C. The Role of Receptor-like Protein Kinases (RLKs) in Abiotic Stress Response in Plants. Plant Cell Rep. 2017, 36, 235–242. [Google Scholar] [CrossRef]
- He, Y.; Zhou, J.; Shan, L.; Meng, X. Plant Cell Surface Receptor-Mediated Signaling—A Common Theme amid Diversity. J. Cell Sci. 2018, 131, jcs209353. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Lozano-Duran, R. Molecular Dialogues between Viruses and Receptor-like Kinases in Plants. Mol. Plant Pathol. 2019, 20, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tax, F.E. Receptor-Like Kinases: Key Regulators of Plant Development and Defense. J. Integr. Plant Biol. 2013, 55, 1184–1187. [Google Scholar] [CrossRef]
- Kørner, C.J.; Klauser, D.; Niehl, A.; Domínguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The Immunity Regulator BAK1 Contributes to Resistance Against Diverse RNA Viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Zorzatto, C.; Machado, J.P.B.; Lopes, K.V.G.; Nascimento, K.J.T.; Pereira, W.A.; Brustolini, O.J.B.; Reis, P.A.B.; Calil, I.P.; Deguchi, M.; Sachetto-Martins, G.; et al. NIK1-Mediated Translation Suppression Functions as a Plant Antiviral Immunity Mechanism. Nature 2015, 520, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.B.; Santos, A.A.; Fontes, E.P.B. Immune Receptors and Co-Receptors in Antiviral Innate Immunity in Plants. Front. Microbiol. 2016, 7, 2139. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-Stranded RNAs Induce a Pattern-Triggered Immune Signaling Pathway in Plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef] [Green Version]
- Calil, I.P.; Fontes, E.P.B. Plant Immunity against Viruses: Antiviral Immune Receptors in Focus. Ann. Bot. 2017, 119, mcw200. [Google Scholar] [CrossRef]
- Carluccio, A.V.; Prigigallo, M.I.; Rosas-Diaz, T.; Lozano-Duran, R.; Stavolone, L. S-acylation Mediates Mungbean Yellow Mosaic Virus AC4 Localization to the Plasma Membrane and in Turns Gene Silencing Suppression. PLOS Pathog. 2018, 14, e1007207. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zeng, R.; Chen, Z.; Liu, X.; Cao, Z.; Xie, Q.; Yang, C.; Lai, J. S-Acylation of a Geminivirus C4 Protein Is Essential for Regulating the CLAVATA Pathway in Symptom Determination. J. Exp. Bot. 2018, 69, 4459–4468. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Diaz, T.; Zhang, D.; Fan, P.; Wang, L.; Ding, X.; Jiang, Y.; Jimenez-Gongora, T.; Medina-Puche, L.; Zhao, X.; Feng, Z.; et al. A Virus-Targeted Plant Receptor-like Kinase Promotes Cell-to-Cell Spread of RNAi. Proc. Natl. Acad. Sci. USA 2018, 115, 1388–1393. [Google Scholar] [CrossRef]
- Fontes, E.P.B.; Santos, A.A.; Luz, D.F.; Waclawovsky, A.J.; Chory, J. The Geminivirus Nuclear Shuttle Protein Is a Virulence Factor That Suppresses Transmembrane Receptor Kinase Activity. Genes Dev. 2004, 18, 2545–2556. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.B.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P.B. Regulated Nuclear Trafficking of RpL10A Mediated by NIK1 Represents a Defense Strategy of Plant Cells against Virus. PLoS Pathog. 2008, 4, e1000247. [Google Scholar] [CrossRef]
- Rocha, C.S.; Santos, A.A.; Machado, J.P.B.; Fontes, E.P.B. The Ribosomal Protein L10/QM-like Protein Is a Component of the NIK-Mediated Antiviral Signaling. Virology 2008, 380, 165–169. [Google Scholar] [CrossRef]
- Zeng, R.; Liu, X.; Yang, C.; Lai, J. Geminivirus C4: Interplaying with Receptor-like Kinases. Trends Plant Sci. 2018, 23, 1044–1046. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, G.; Suzuki, H.; Miyashita, Y.; Choi, S.H.; Hisa, Y.; Rihei, S.; Shimada, R.; Jeon, E.J.; Abe, J.; Nakahara, K.S.; et al. P3N-PIPO, a Frameshift Product from the P3 Gene, Pleiotropically Determines the Virulence of Clover Yellow Vein Virus in Both Resistant and Susceptible Peas. J. Virol. 2016, 90, 7388–7404. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, R.; Ye, Z.; MacFarlane, S.; Li, Y.; Mao, Q.; Tian, Y.; Deng, Z.; Sun, Z.; Yang, J.; Li, J.; et al. The P3N-PIPO Protein Encoded by Wheat Yellow Mosaic Virus Is a Pathogenicity Determinant and Promotes Its Pathogenicity through Interaction with NbRLK6 in Nicotiana benthamiana. Viruses 2022, 14, 2171. https://doi.org/10.3390/v14102171
Miao R, Ye Z, MacFarlane S, Li Y, Mao Q, Tian Y, Deng Z, Sun Z, Yang J, Li J, et al. The P3N-PIPO Protein Encoded by Wheat Yellow Mosaic Virus Is a Pathogenicity Determinant and Promotes Its Pathogenicity through Interaction with NbRLK6 in Nicotiana benthamiana. Viruses. 2022; 14(10):2171. https://doi.org/10.3390/v14102171
Chicago/Turabian StyleMiao, Runpu, Zhuangxin Ye, Stuart MacFarlane, Yanjun Li, Qianzhuo Mao, Yanzhen Tian, Zhiping Deng, Zongtao Sun, Jian Yang, Junmin Li, and et al. 2022. "The P3N-PIPO Protein Encoded by Wheat Yellow Mosaic Virus Is a Pathogenicity Determinant and Promotes Its Pathogenicity through Interaction with NbRLK6 in Nicotiana benthamiana" Viruses 14, no. 10: 2171. https://doi.org/10.3390/v14102171