
Ivermectin Inhibits the Replication of Usutu Virus In Vitro

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viruses
2.3. Viral Replication Kinetics
2.4. Viral Titer Determination
2.5. Immunofluorescence Analysis
2.6. Substances
2.7. Cytotoxicity Assay
2.8. Screening for Antiviral Activity on USUV by FDA-Approved Compounds
2.9. Cell Line- and Lineage-Dependent Characterization of Antiviral Molecules
2.10. Statistical Analysis
3. Results
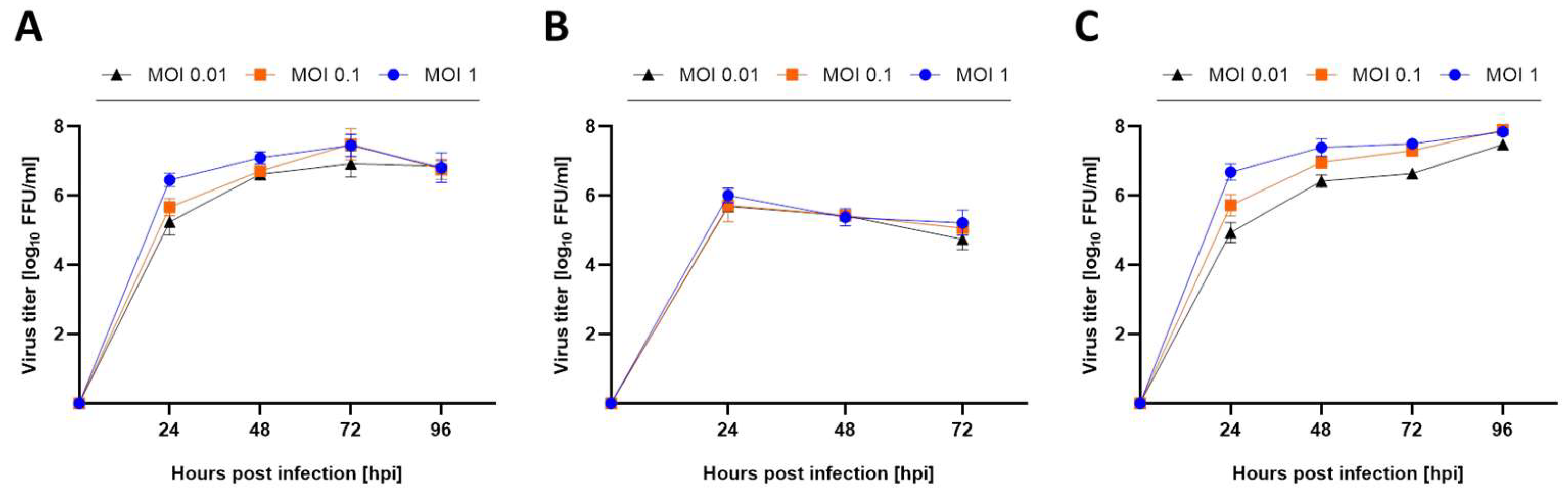
3.1. Mammalian and Avian Cell Lines Are Susceptible to USUV Infection
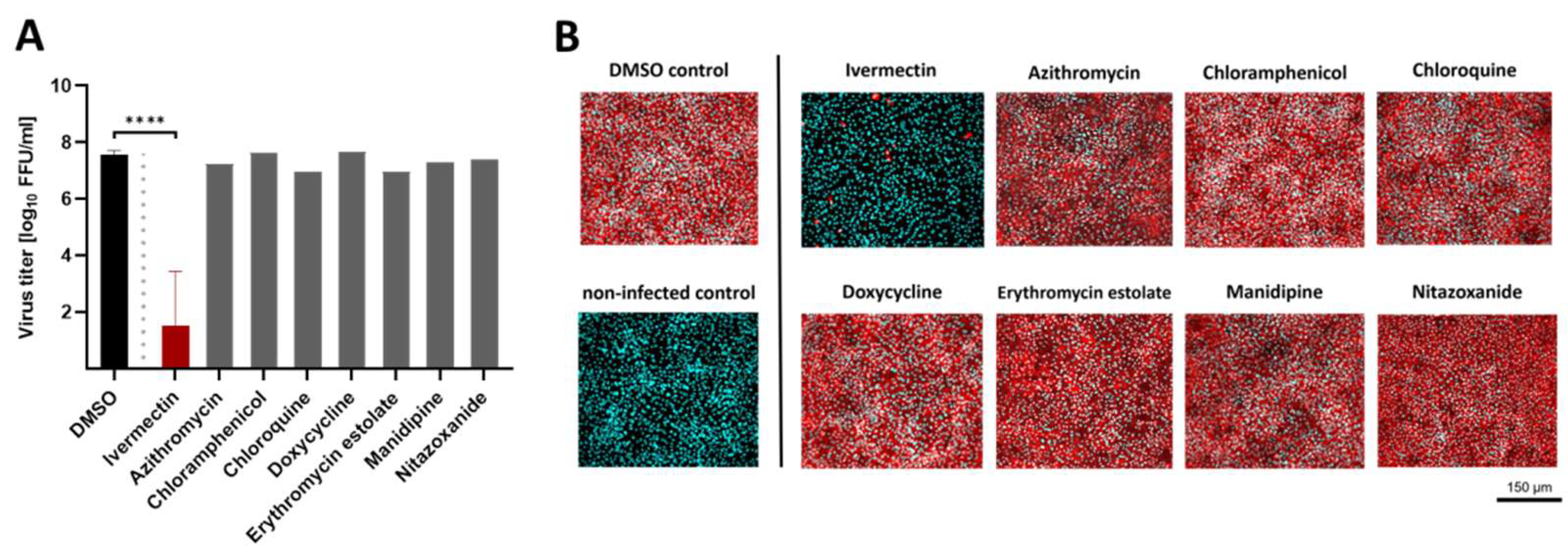
3.2. The FDA-Approved Compound Ivermectin Was Effective against USUV In Vitro
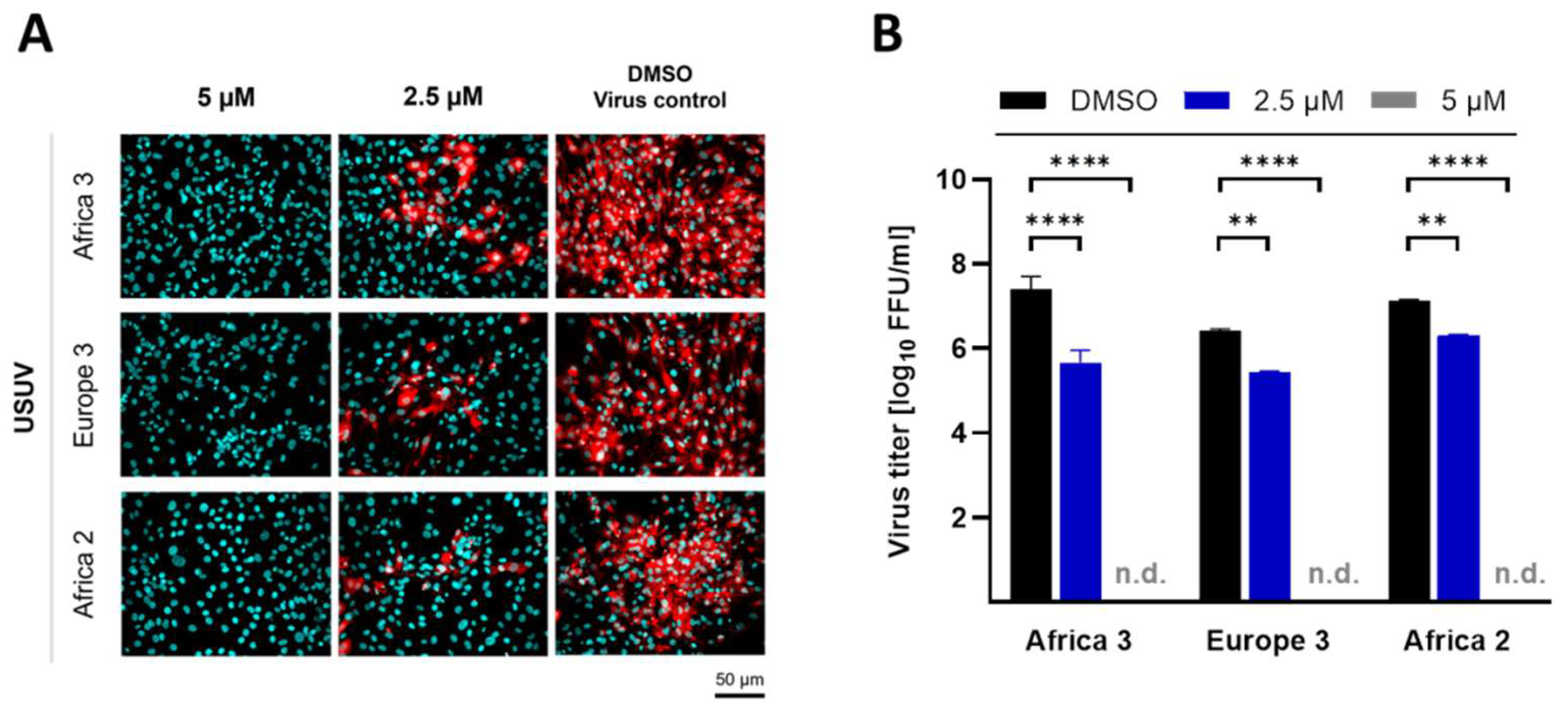
3.3. Ivermectin Inhibits USUV Infectivity in a Dose-Dependent Manner
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, H.W.; Bakonyi, T.; Weissenböck, H.; Hatt, J.-M.; Eulenberger, U.; Robert, N.; Hoop, R.; Nowotny, N. Emergence and establishment of Usutu virus infection in wild and captive avian species in and around Zurich, Switzerland-genomic and pathologic comparison to other central European outbreaks. Vet. Microbiol. 2011, 148, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijks, J.M.; Kik, M.L.; Slaterus, R.; Foppen, R.; Stroo, A.; IJzer, J.; Stahl, J.; Gröne, A.; Koopmans, M.; van der Jeugd, H.P.; et al. Widespread Usutu virus outbreak in birds in The Netherlands, 2016. Eurosurveillance 2016, 21, 30391. [Google Scholar] [CrossRef]
- Sieg, M.; Schmidt, V.; Ziegler, U.; Keller, M.; Höper, D.; Heenemann, K.; Rückner, A.; Nieper, H.; Muluneh, A.; Groschup, M.H.; et al. Outbreak and Cocirculation of Three Different Usutu Virus Strains in Eastern Germany. Vector-Borne Zoonotic Dis. 2017, 17, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveillance 2017, 22, 30452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu Virus in Germany and Its Effect on Breeding Bird Populations. Emerg. Infect. Dis. 2017, 23, 1994–2001. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- Williams, M.C.; Simpson, D.I.; Haddow, A.J.; Knight, E.M. The isolation of west nile virus from man and of usutu virus from the bird-biting mosquito mansonia aurites (theobald) in the entebbe area of Uganda. Ann. Trop. Med. Parasitol. 1964, 58, 367–374. [Google Scholar] [CrossRef]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef]
- Mannasse, B.; Mendelson, E.; Orshan, L.; Mor, O.; Shalom, U.; Yeger, T.; Lustig, Y. Usutu Virus RNA in Mosquitoes, Israel, 2014–2015. Emerg. Infect. Dis. 2017, 23, 1699–1702. [Google Scholar] [CrossRef]
- Chvala, S.; Kolodziejek, J.; Nowotny, N.; Weissenböck, H. Pathology and viral distribution in fatal Usutu virus infections of birds from the 2001 and 2002 outbreaks in Austria. J. Comp. Pathol. 2004, 131, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu Virus: An Arbovirus on the Rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [Green Version]
- Giglia, G.; Agliani, G.; Munnink, B.B.O.; Sikkema, R.S.; Mandara, M.T.; Lepri, E.; Kik, M.; Ijzer, J.; Rijks, J.M.; Fast, C.; et al. Pathology and Pathogenesis of Eurasian Blackbirds (Turdus merula) Naturally Infected with Usutu Virus. Viruses 2021, 13, 1481. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, V.; Cramer, K.; Böttcher, D.; Heenemann, K.; Rückner, A.; Harzer, M.; Ziegler, U.; Vahlenkamp, T.; Sieg, M. Usutu virus infection in aviary birds during the cold season. Avian Pathol. 2021, 50, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, E.; Sarlet, M.; Franssen, M.; Cadar, D.; Schmidt-Chanasit, J.; Rivas, J.F.; Linden, A.; Desmecht, D.; Garigliany, M. Usutu Virus Epizootic in Belgium in 2017 and 2018: Evidence of Virus Endemization and Ongoing Introduction Events. Vector-Borne Zoonotic Dis. 2020, 20, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Bergmann, F.; Fischer, D.; Müller, K.; Holicki, C.M.; Sadeghi, B.; Sieg, M.; Keller, M.; Schwehn, R.; Reuschel, M.; et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms 2022, 10, 807. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zu, S.; Deng, Y.-Q.; Li, D.; Parvatiyar, K.; Quanquin, N.; Shang, J.; Sun, N.; Su, J.; Liu, Z.; et al. Azithromycin Protects against Zika virus Infection by Upregulating virus-induced Type I and III Interferon Responses. Antimicrob. Agents Chemother. 2019, 63, e00394-19. [Google Scholar] [CrossRef]
- Wang, X.; Xia, S.; Zou, P.; Lu, L. Erythromycin Estolate Inhibits Zika Virus Infection by Blocking Viral Entry as a Viral Inactivator. Viruses 2019, 11, 1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.-Y.; Xu, Y.-f.; Zhang, T.-H.; Yang, J.-J.; Yuan, Y.; Hao, P.; Shi, Y.; Zhong, J.; Zhong, W. Pediatric Drug Nitazoxanide: A Potential Choice for Control of Zika. Open Forum Infect. Dis. 2017, 4, ofx009. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Wei, J.; Deng, X.; Li, S.; Qiu, Y.; Shao, D.; Li, B.; Zhang, K.; Xue, F.; Wang, X.; et al. Nitazoxanide inhibits the replication of Japanese encephalitis virus in cultured cells and in a mouse model. Virol. J. 2014, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delvecchio, R.; Higa, L.M.; Pezzuto, P.; Valadão, A.L.; Garcez, P.P.; Monteiro, F.L.; Loiola, E.C.; Dias, A.A.; Silva, F.J.M.; Aliota, M.T.; et al. Chloroquine, an Endocytosis Blocking Agent, Inhibits Zika Virus Infection in Different Cell Models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothan, H.A.; Mohamed, Z.; Paydar, M.; Rahman, N.A.; Yusof, R. Inhibitory effect of doxycycline against dengue virus replication in vitro. Arch. Virol. 2014, 159, 711–718. [Google Scholar] [CrossRef]
- Chong Teoh, T.; J Al-Harbi, S.; Abdulrahman, A.Y.; Rothan, H.A. Doxycycline Interferes with Zika Virus Serine Protease and Inhibits Virus Replication in Human Skin Fibroblasts. Molecules 2021, 26, 4321. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, Y.; Guo, J.; Wang, P.; Zhang, L.; Xiao, G.; Wang, W. Screening of FDA-Approved Drugs for Inhibitors of Japanese Encephalitis Virus Infection. J. Virol. 2017, 91, e01055-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Shi, P.-Y.; Li, H.; Zhou, J. Broad Spectrum Antiviral Agent Niclosamide and Its Therapeutic Potential. ACS Infect. Dis. 2020, 6, 909–915. [Google Scholar] [CrossRef]
- Kinobe, R.T.; Owens, L. A systematic review of experimental evidence for antiviral effects of ivermectin and an in silico analysis of ivermectin’s possible mode of action against SARS-CoV-2. Fundam. Clin. Pharmacol. 2021, 35, 260–276. [Google Scholar] [CrossRef]
- Burg, R.W.; Miller, B.M.; Baker, E.E.; Birnbaum, J.; Currie, S.A.; Hartman, R.; Kong, Y.L.; Monaghan, R.L.; Olson, G.; Putter, I.; et al. Avermectins, new family of potent anthelmintic agents: Producing organism and fermentation. Antimicrob. Agents Chemother. 1979, 15, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Lindley, D. Merck’s new drug free to WHO for river blindness programme. Nature 1987, 329, 752. [Google Scholar] [CrossRef] [Green Version]
- Omura, S.; Crump, A. The life and times of ivermectin—A success story. Nat. Rev. Microbiol. 2004, 2, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Brien, J.D.; Lazear, H.M.; Diamond, M.S. Propagation, quantification, detection, and storage of West Nile virus. Curr. Protoc. Microbiol. 2013, 31, 15D.3.1–15D.3.18. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, H.; Ishiyama, M.; Ohseto, F.; Sasamoto, K.; Hamamoto, T.; Suzuki, K.; Watanabe, M. A water-soluble tetrazolium salt useful for colorimetric cell viability assay. Anal. Commun. 1999, 36, 47–50. [Google Scholar] [CrossRef]
- Kuchinsky, S.C.; Hawks, S.A.; Mossel, E.C.; Coutermarsh-Ott, S.; Duggal, N.K. Differential pathogenesis of Usutu virus isolates in mice. PLoS Negl. Trop. Dis. 2020, 14, e0008765. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Hoeper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D.E.; et al. Evidence for an independent third Usutu virus introduction into Germany. Vet. Microbiol. 2016, 192, 60–66. [Google Scholar] [CrossRef]
- Tricou, V.; Minh, N.N.; Van, T.P.; Lee, S.J.; Farrar, J.; Wills, B.; Tran, H.T.; Simmons, C.P. A randomized controlled trial of chloroquine for the treatment of dengue in Vietnamese adults. PLoS Negl. Trop. Dis. 2010, 4, e785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.-D.; Meng, W.; Wang, X.-J.; Wang, H.-C.R. Broad-spectrum antiviral agents. Front. Microbiol. 2015, 6, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, L.L.; Padilla, L.; Castaño, J.C. Inhibitors compounds of the flavivirus replication process. Virol. J. 2017, 14, 95. [Google Scholar] [CrossRef]
- Mazzon, M.; Ortega-Prieto, A.M.; Imrie, D.; Luft, C.; Hess, L.; Czieso, S.; Grove, J.; Skelton, J.K.; Farleigh, L.; Bugert, J.J.; et al. Identification of Broad-Spectrum Antiviral Compounds by Targeting Viral Entry. Viruses 2019, 11, 176. [Google Scholar] [CrossRef]
- Farias, K.J.S.; Machado, P.R.L.; da Fonseca, B.A.L. Chloroquine inhibits dengue virus type 2 replication in Vero cells but not in C6/36 cells. Sci. World J. 2013, 2013, 282734. [Google Scholar] [CrossRef] [Green Version]
- Desmyter, J.; Melnick, J.L.; Rawls, W.E. Defectiveness of interferon production and of rubella virus interference in a line of African green monkey kidney cells (Vero). J. Virol. 1968, 2, 955–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The genome landscape of the african green monkey kidney-derived vero cell line. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2014, 21, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, M.Y.F.; Fraser, J.E.; Chan, W.K.K.; Moreland, N.J.; Rathore, A.P.; Wang, C.; Vasudevan, S.G.; Jans, D.A. Nuclear localization of dengue virus (DENV) 1-4 non-structural protein 5; protection against all 4 DENV serotypes by the inhibitor Ivermectin. Antivir. Res. 2013, 99, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Holcomb, K.M.; Nguyen, C.; Foy, B.D.; Ahn, M.; Cramer, K.; Lonstrup, E.T.; Mete, A.; Tell, L.A.; Barker, C.M. Effects of ivermectin treatment of backyard chickens on mosquito dynamics and West Nile virus transmission. PLoS Negl. Trop. Dis. 2022, 16, e0010260. [Google Scholar] [CrossRef]
- Cirak, V.Y.; Aksit, D.; Cihan, H.; Gokbulut, C. Plasma dispositions and concentrations of ivermectin in eggs following treatment of laying hens. N. Z. Vet. J. 2018, 66, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Lierz, M. Evaluation of the dosage of ivermectin in falcons. Vet. Rec. 2001, 148, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, E.; Pezzullo, M.; de Burghgraeve, T.; Kaptein, S.; Pastorino, B.; Dallmeier, K.; de Lamballerie, X.; Neyts, J.; Hanson, A.M.; Frick, D.N.; et al. Ivermectin is a potent inhibitor of flavivirus replication specifically targeting NS3 helicase activity: New prospects for an old drug. J. Antimicrob. Chemother. 2012, 67, 1884–1894. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.N.Y.; Atkinson, S.C.; Wang, C.; Lee, A.; Bogoyevitch, M.A.; Borg, N.A.; Jans, D.A. The broad spectrum antiviral ivermectin targets the host nuclear transport importin α/β1 heterodimer. Antivir. Res. 2020, 177, 104760. [Google Scholar] [CrossRef] [PubMed]
- Blakley, B.R.; Rousseaux, C.G. Effect of ivermectin on the immune response in mice. Am. J. Vet. Res. 1991, 52, 593–595. [Google Scholar] [PubMed]
- Bray, M.; Rayner, C.; Noël, F.; Jans, D.; Wagstaff, K. Ivermectin and COVID-19: A report in Antiviral Research, widespread interest, an FDA warning, two letters to the editor and the authors’ responses. Antivir. Res. 2020, 178, 104805. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.D.; Maia, A. A COVID-19 prophylaxis? Lower incidence associated with prophylactic administration of ivermectin. Int. J. Antimicrob. Agents 2021, 57, 106248. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.W. Pharmacokinetics and metabolism of avermectins in livestock. Vet. Parasitol. 1993, 48, 45–57. [Google Scholar] [CrossRef]
- Wang, X.; Lv, C.; Ji, X.; Wang, B.; Qiu, L.; Yang, Z. Ivermectin treatment inhibits the replication of Porcine circovirus 2 (PCV2) in vitro and mitigates the impact of viral infection in piglets. Virus Res. 2019, 263, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Liu, W.; Wang, B.; Dang, R.; Qiu, L.; Ren, J.; Yan, C.; Yang, Z.; Wang, X. Ivermectin inhibits DNA polymerase UL42 of pseudorabies virus entrance into the nucleus and proliferation of the virus in vitro and vivo. Antivir. Res. 2018, 159, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Ketkar, H.; Yang, L.; Wormser, G.P.; Wang, P. Lack of efficacy of ivermectin for prevention of a lethal Zika virus infection in a murine system. Diagn. Microbiol. Infect. Dis. 2019, 95, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Karim, M.M.; Ross, A.G.; Hossain, M.S.; Clemens, J.D.; Sumiya, M.K.; Phru, C.S.; Rahman, M.; Zaman, K.; Somani, J.; et al. A five-day course of ivermectin for the treatment of COVID-19 may reduce the duration of illness. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2021, 103, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Buonfrate, D.; Chesini, F.; Martini, D.; Roncaglioni, M.C.; Ojeda Fernandez, M.L.; Alvisi, M.F.; de Simone, I.; Rulli, E.; Nobili, A.; Casalini, G.; et al. High-dose ivermectin for early treatment of COVID-19 (COVER study): A randomised, double-blind, multicentre, phase II, dose-finding, proof-of-concept clinical trial. Int. J. Antimicrob. Agents 2022, 59, 106516. [Google Scholar] [CrossRef]
- Eerike, M.; Raj, G.M.; Priyadarshini, R.; Ravi, G.; Bisoi, D.; Konda, V.G.R. Ivermectin in COVID-19 Management: What is the current evidence? Infect. Disord. Drug Targets 2022, 22, e190122200367. [Google Scholar] [CrossRef] [PubMed]
- Ozer, M.; Goksu, S.Y.; Conception, R.; Ulker, E.; Balderas, R.M.; Mahdi, M.; Manning, Z.; To, K.; Effendi, M.; Anandakrishnan, R.; et al. Effectiveness and safety of Ivermectin in COVID-19 patients: A prospective study at a safety-net hospital. J. Med. Virol. 2022, 94, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M. Ivermectin as a possible treatment for West Nile virus in raptors. HawkChalk 2018, 80–82. [Google Scholar]
- Straub, J.; Pohl, A.; Enders, F.; Will, H.; Ziegler, U.; Fischer, D. (Eds.) Beschreibung Nicht-Letaler, Klinischer Fälle von Usutu- und West-Nil-Virus-Infektionen bei Greifvögeln und Eulen, 1st ed.; Jahrestagung der DVG-Fachgruppe: Zier-, Zoo- und Wildvögel, Reptilien, Amphibien und Fische (ZZWRAF), Online-Fortbildung, 4–6 March 2021; Verlag der DVG Service GmbH: Gießen, Germany, 2021. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Inhibition Efficacy IC50 (µM) | Cell Viability CC50 (µM) | Selectivity Index SI (CC50/IC50) |
|---|---|---|---|
| Vero CCL-81 | 0.55 ± 0.03 | 7.24 ± 0.67 | 13.16 |
| A549 | 1.94 ± 0.21 | 15.18 ± 1.33 | 7.82 |
| TME-R | 1.38 ± 0.16 | 8.26 ± 1.11 | 5.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wald, M.E.; Claus, C.; Konrath, A.; Nieper, H.; Muluneh, A.; Schmidt, V.; Vahlenkamp, T.W.; Sieg, M. Ivermectin Inhibits the Replication of Usutu Virus In Vitro. Viruses 2022, 14, 1641. https://doi.org/10.3390/v14081641
Wald ME, Claus C, Konrath A, Nieper H, Muluneh A, Schmidt V, Vahlenkamp TW, Sieg M. Ivermectin Inhibits the Replication of Usutu Virus In Vitro. Viruses. 2022; 14(8):1641. https://doi.org/10.3390/v14081641
Chicago/Turabian StyleWald, Maria Elisabeth, Claudia Claus, Andrea Konrath, Hermann Nieper, Aemero Muluneh, Volker Schmidt, Thomas Wilhelm Vahlenkamp, and Michael Sieg. 2022. "Ivermectin Inhibits the Replication of Usutu Virus In Vitro" Viruses 14, no. 8: 1641. https://doi.org/10.3390/v14081641