
The Role of Lebanon in the COVID-19 Butterfly Effect: The B.1.398 Example

 , , , , , , , , and
, , , , , , , , and 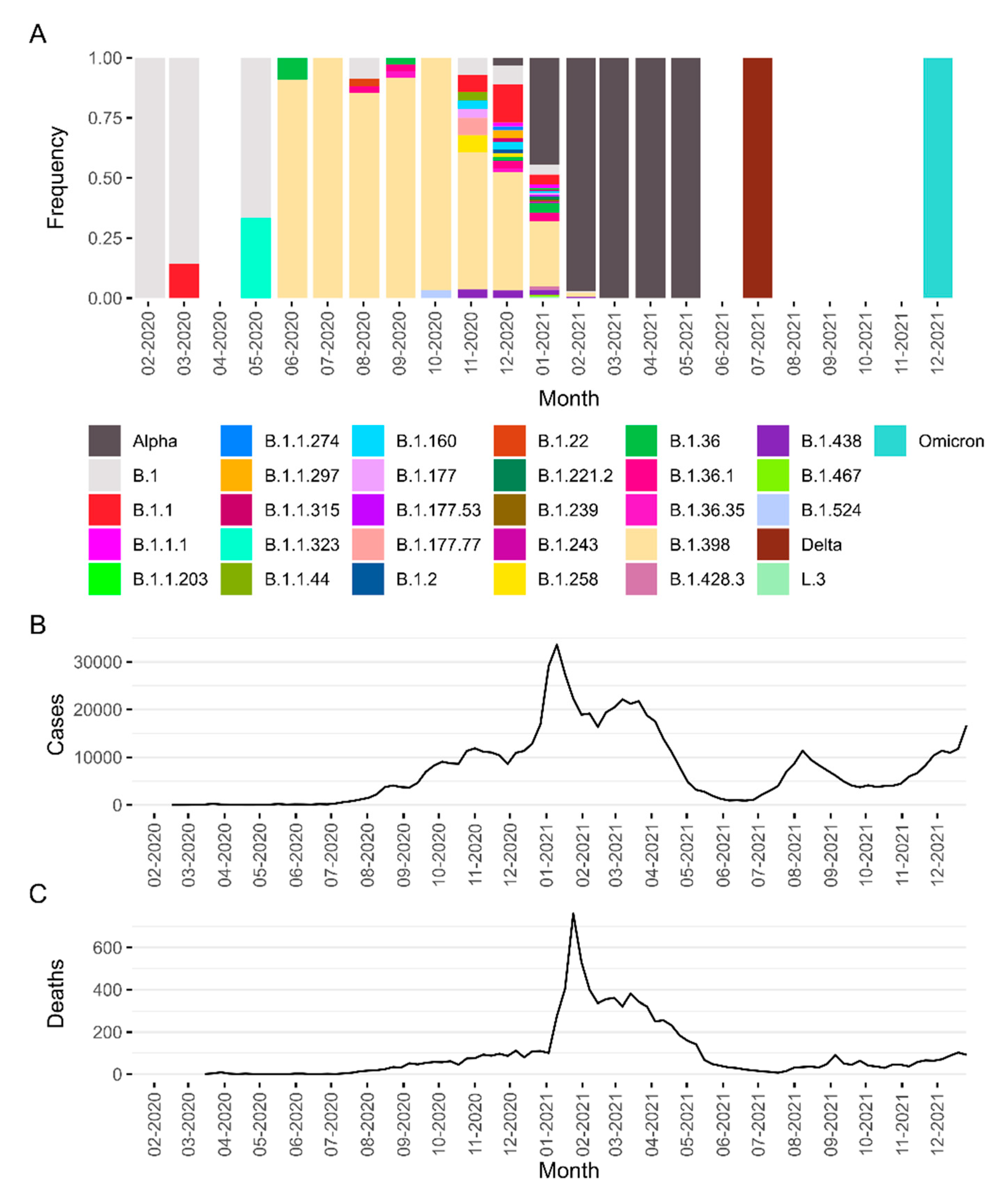{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Collecting Information and Handling Samples
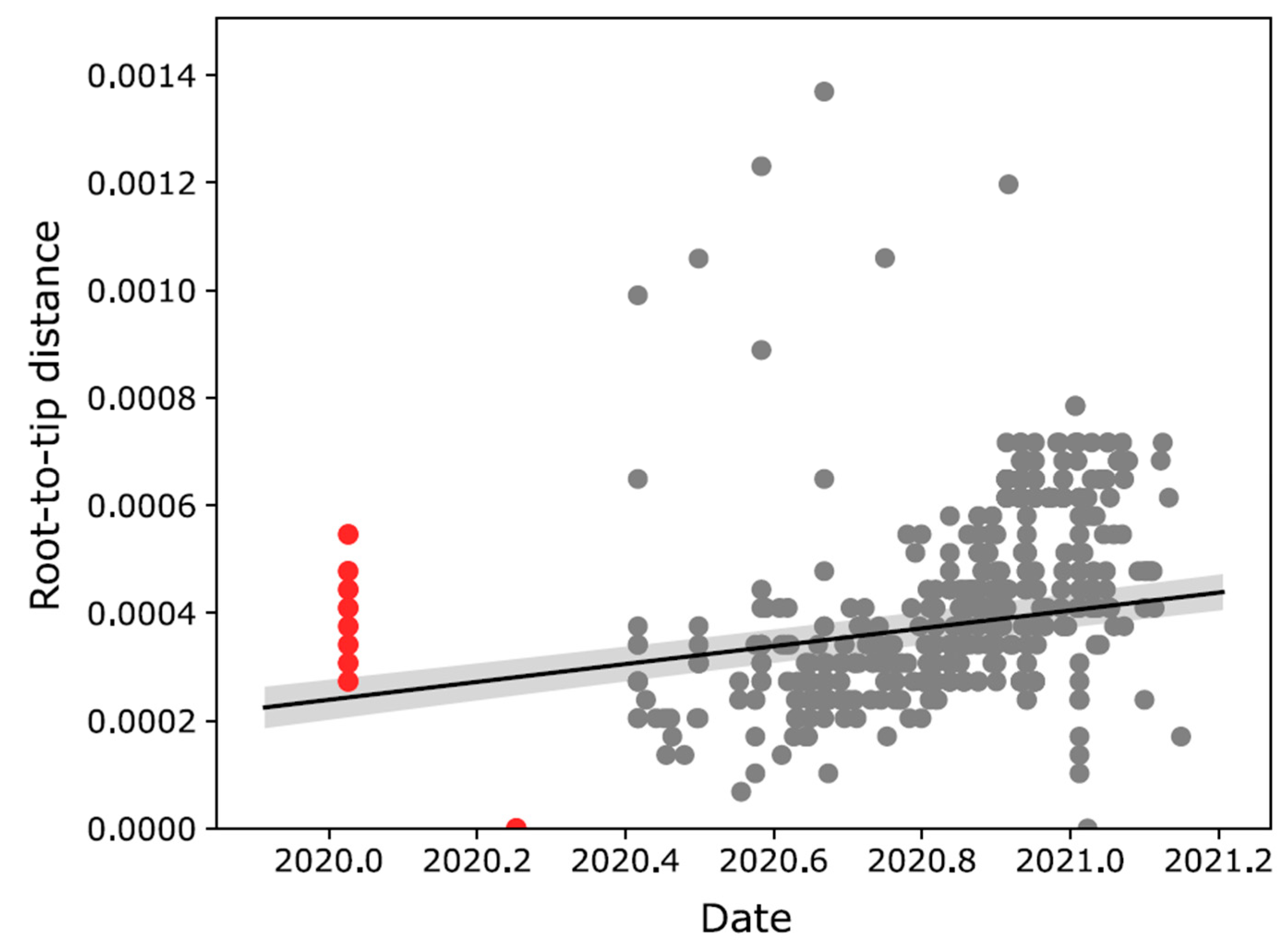
2.3. Evolutionary and Phylogeographic Analyses
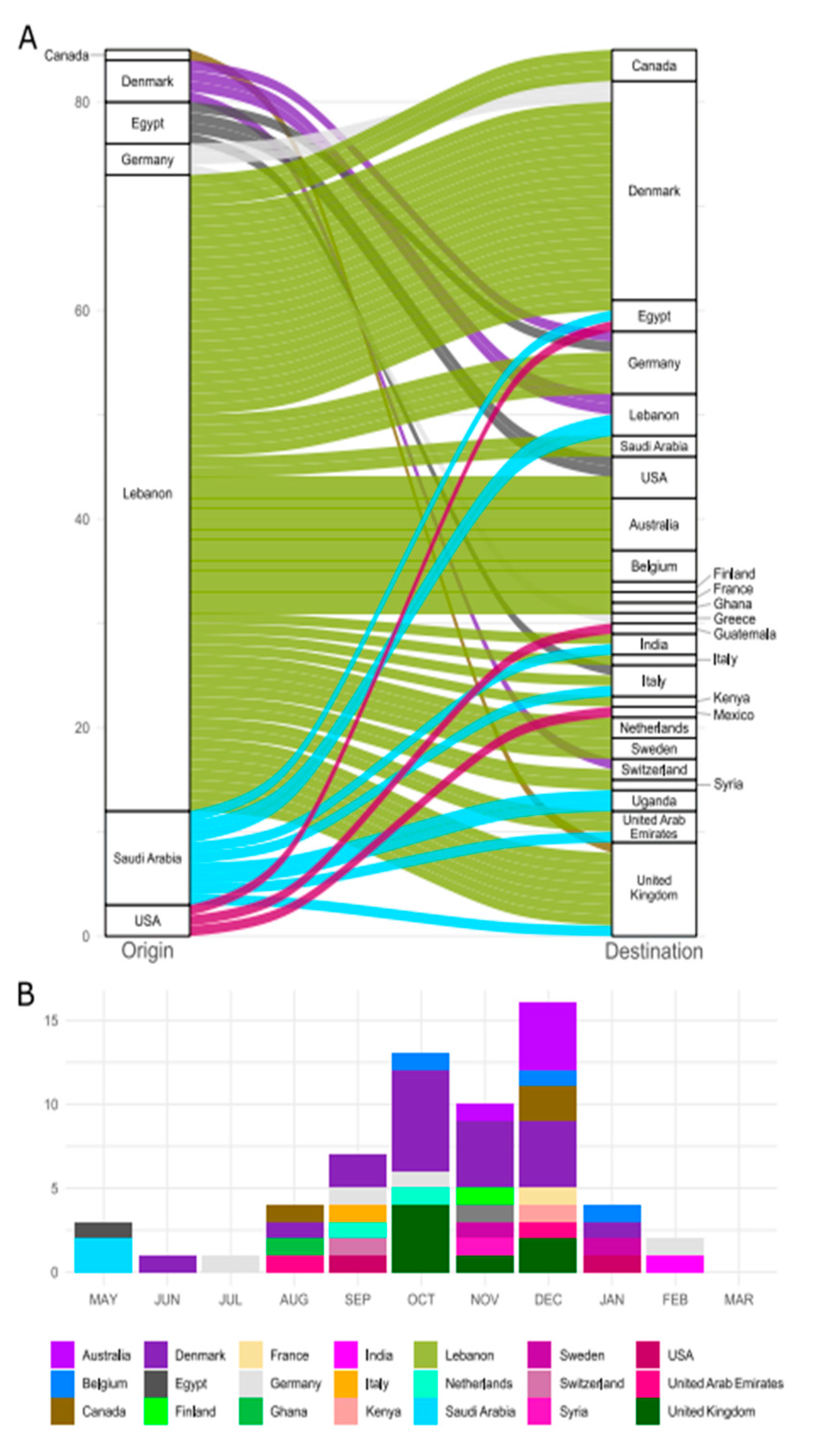
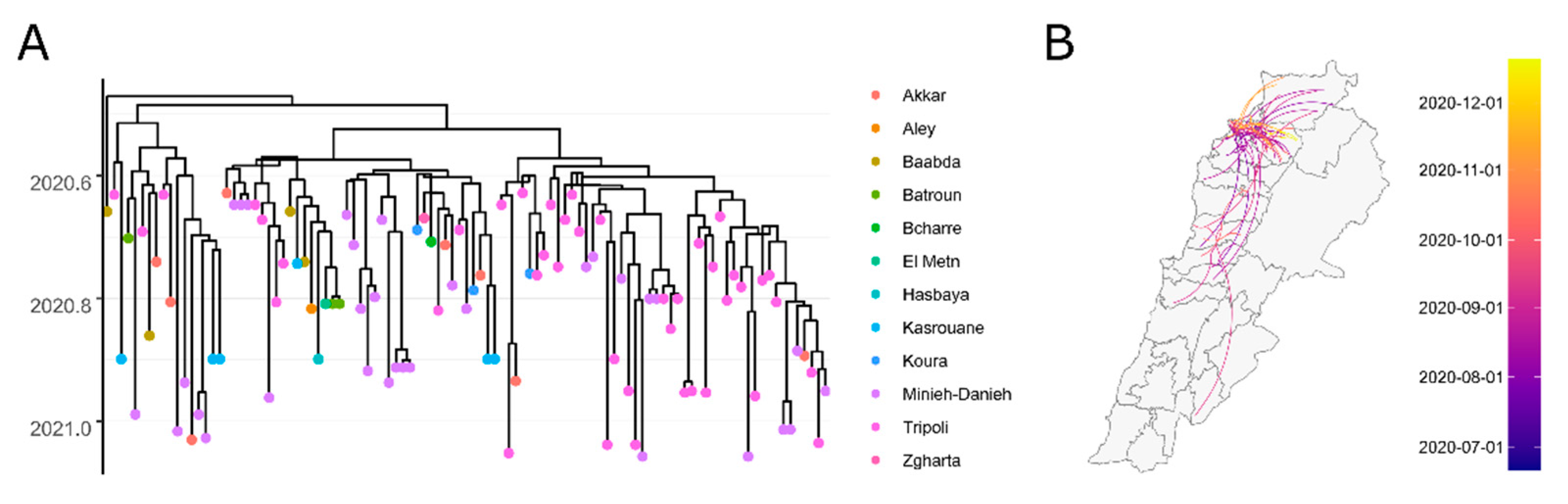
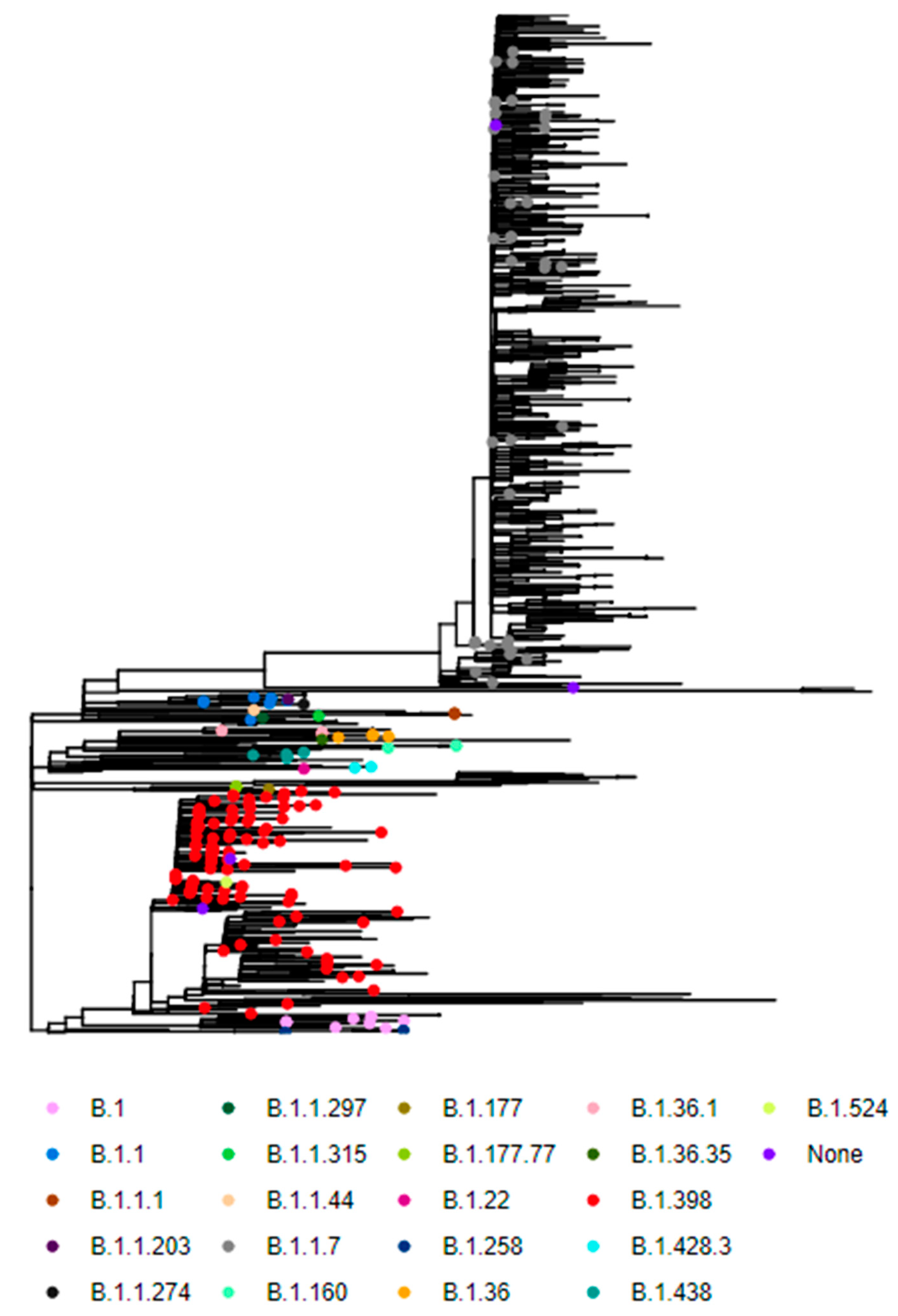
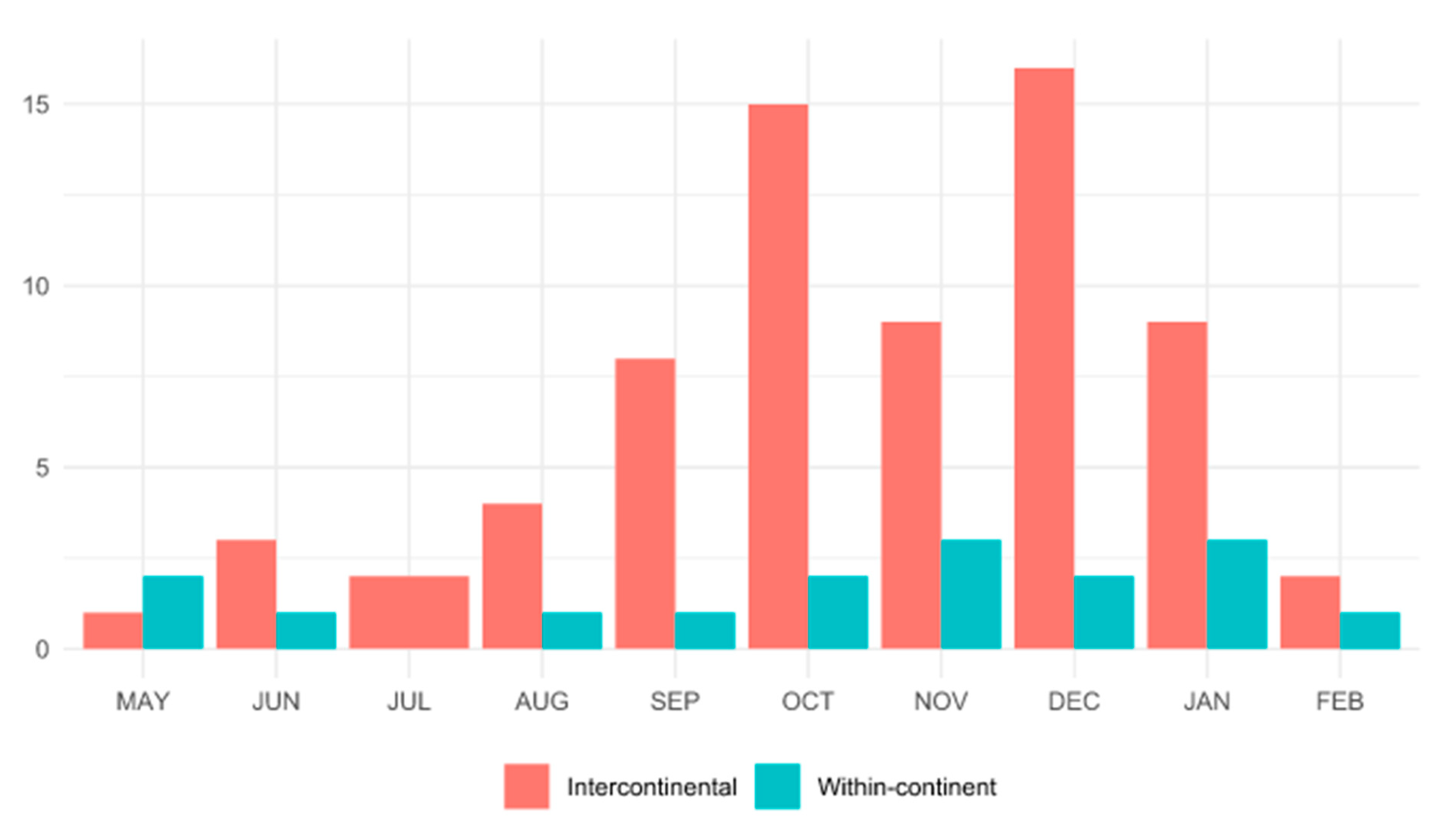
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Our World in Data. Available online: https://ourworldindata.org/explorers/coronavirus-data-explorer (accessed on 30 March 2022).
- Pango Lineages: Latest Epidemiological Lineages of SARS-CoV-2. Available online: https://cov-lineages.org/ (accessed on 30 March 2022).
- WHO: Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 30 March 2022).
- Brussow, H. COVID-19: Emergence and mutational diversification of SARS-CoV-2. Microb. Biotechnol. 2021, 14, 756–768. [Google Scholar] [CrossRef]
- Dubey, A.; Choudhary, S.; Kumar, P.; Tomar, S. Emerging SARS-CoV-2 Variants: Genetic Variability and Clinical Implications. Curr. Microbiol. 2021, 79, 20. [Google Scholar] [CrossRef] [PubMed]
- World Economic Situation and Prospects. Country Classification. Available online: https://www.un.org/en/development/desa/policy/wesp/wesp_current/2014wesp_country_classification.pdf (accessed on 16 May 2022).
- Elbe, S.; Buckland-Merrett, G. Data, disease and diplomacy: GISAID’s innovative contribution to global health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GISAID. Available online: https://www.gisaid.org/submission-tracker-global/ (accessed on 16 May 2022).
- Chen, Z.; Azman, A.S.; Chen, X.; Zou, J.; Tian, Y.; Sun, R.; Xu, X.; Wu, Y.; Lu, W.; Ge, S.; et al. Global landscape of SARS-CoV-2 genomic surveillance and data sharing. Nat. Genet. 2022, 54, 499–507. [Google Scholar] [CrossRef]
- Brito, A.F.; Semenova, E.; Dudas, G.; Hassler, G.W.; Kalinich, C.C.; Kraemer, M.U.G.; Ho, J.; Tegally, H.; Githinji, G.; Agoti, C.N.; et al. Global disparities in SARS-CoV-2 genomic surveillance. medRxiv 2021. [Google Scholar] [CrossRef]
- Koweyes, J.; Salloum, T.; Haidar, S.; Merhi, G.; Tokajian, S. COVID-19 Pandemic in Lebanon: One Year Later, What Have We Learnt? mSystems 2021, 6, e00351-21. [Google Scholar] [CrossRef]
- WHO Health Emergency Dashboard. Available online: https://covid19.who.int/region/emro/country/lb (accessed on 17 April 2022).
- Republic of Lebanon Ministry of Public Health (MOPH). Epidemiological Surveillance Program. Official Updates Coronavirus-COVID-19 In Lebanon. Available online: https://www.moph.gov.lb/en/ (accessed on 17 April 2022).
- GISAID. Available online: https://www.gisaid.org/ (accessed on 4 February 2022).
- Bal, A.; Simon, B.; Destras, G.; Chalvignac, R.; Semanas, Q.; Oblette, A.; Queromes, G.; Fanget, R.; Regue, H.; Morfin, F.; et al. Detection and prevalence of SARS-CoV-2 co-infections during the Omicron variant circulation, France, December 2021—February 2022. medRxiv 2022. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Holmes, E.C.; O’Toole, A.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Sagulenko, P.; Puller, V.; Neher, R.A. TreeTime: Maximum-likelihood phylodynamic analysis. Virus Evol. 2018, 4, vex042. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Dellicour, S.; Rose, R.; Faria, N.R.; Lemey, P.; Pybus, O.G. SERAPHIM: Studying environmental rasters and phylogenetically informed movements. Bioinformatics 2016, 32, 3204–3206. [Google Scholar] [CrossRef]
- Fayad, N.; Abi Habib, W.; Kandeil, A.; El-Shesheny, R.; Kamel, M.N.; Mourad, Y.; Mokhbat, J.; Kayali, G.; Goldstein, J.; Abdallah, J. SARS-CoV-2 Variants in Lebanon: Evolution and Current Situation. Biology 2021, 10, 531. [Google Scholar] [CrossRef]
- Feghali, R.; Merhi, G.; Kwasiborski, A.; Hourdel, V.; Ghosn, N.; Tokajian, S. Genomic characterization and phylogenetic analysis of the first SARS-CoV-2 variants introduced in Lebanon. PeerJ 2021, 9, e11015. [Google Scholar] [CrossRef] [PubMed]
- Merhi, G.T.A.; Martins, L.; Koweyes, J.; Le-Viet, T.; Abou Naja, H.; Al Buaini, M.; Prosolek, S.; Alikhan, N.F.; Lott, M.; Tohmeh, T.; et al. Replacement of the Alpha variant of SARS-CoV-2 by the Delta variant in Lebanon between April and June 2021. MedRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Lineage List. Available online: https://cov-lineages.org/lineage_list.html (accessed on 17 April 2022).
- Jackson, C.B.; Zhang, L.; Farzan, M.; Choe, H. Functional importance of the D614G mutation in the SARS-CoV-2 spike protein. Biochem. Biophys Res. Commun. 2021, 538, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Isabel, S.; Grana-Miraglia, L.; Gutierrez, J.M.; Bundalovic-Torma, C.; Groves, H.E.; Isabel, M.R.; Eshaghi, A.; Patel, S.N.; Gubbay, J.B.; Poutanen, T.; et al. Evolutionary and structural analyses of SARS-CoV-2 D614G spike protein mutation now documented worldwide. Sci. Rep. 2020, 10, 14031. [Google Scholar] [CrossRef] [PubMed]
- Plante, J.A.; Liu, Y.; Liu, J.; Xia, H.; Johnson, B.A.; Lokugamage, K.G.; Zhang, X.; Muruato, A.E.; Zou, J.; Fontes-Garfias, C.R.; et al. Spike mutation D614G alters SARS-CoV-2 fitness. Nature 2021, 592, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Medcities. Available online: https://medcities.org/fr/member/tripoli/ (accessed on 10 April 2022).
- Hubbard, B.; Saad, H. Lebanon’s Currency Plunges, and Protesters Surge Into Streets. The New York Times. 2020. Available online: https://www.nytimes.com/2020/06/11/world/middleeast/lebanon-protests.html (accessed on 10 April 2022).
- Cheaito, M.A.; Al-Hajj, S. A Brief Report on the Beirut Port Explosion. Mediterr. J. Emerg. Med. Acute Care 2020, 1. [Google Scholar] [CrossRef]
- Cheeseman, A. After Beirut Explosion, Lebanese Volunteers Flock to Help Clean up. NBC News. 2020. Available online: https://www.nbcnews.com/news/world/after-beirut-explosion-lebanese-volunteers-flock-help-clean-n1236403 (accessed on 17 April 2022).
- Aboud, C. Lebanon: A New General Quarantine Begins to Counter the Rise in Corona Virus Infections (Available in Arabic Version). France24 2020. Available online: https://www.france24.com/ar/ (accessed on 17 April 2022).
- Gomez, C.E.; Perdiguero, B.; Esteban, M. Emerging SARS-CoV-2 Variants and Impact in Global Vaccination Programs against SARS-CoV-2/COVID-19. Vaccines 2021, 9, 243. [Google Scholar] [CrossRef]
- Mumtaz, G.R.; El-Jardali, F.; Jabbour, M.; Harb, A.; Abu-Raddad, L.J.; Makhoul, M. Modeling the Impact of COVID-19 Vaccination in Lebanon: A Call to Speed-Up Vaccine Roll Out. Vaccines 2021, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Tabar, P.; Denison, A. IMISCOE Research Series. Diaspora Policies, Consular Services and Social Protection for Lebanese Citizens Abroad. Migr. Soc. Prot. Eur. Beyond 2020, 3, 199–215. [Google Scholar]
- World Population Review. Available online: https://worldpopulationreview.com/countries/lebanon-population (accessed on 6 April 2022).
- International Labor Organization. Available online: https://www.ilo.org/global/programmes-and-projects/prospects/countries/lebanon/lang--en/index.htm (accessed on 17 April 2022).
- Unated Nations ESCWA. Available online: https://www.unescwa.org/publications/multidimensional-poverty-lebanon-2019-2021 (accessed on 6 April 2022).
- Hubbard, B. As Lebanon’s Crisis Deepens, Lines for Fuel Grow, and Food and Medicine Are Scarce. New York Times. 2021. Available online: https://www.nytimes.com/2021/07/05/world/middleeast/lebanon-economic-crisis.html (accessed on 6 April 2022).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nour, D.; Rafei, R.; Lamarca, A.P.; de Almeida, L.G.P.; Osman, M.; Ismail, M.B.; Mallat, H.; Berry, A.; Burfin, G.; Semanas, Q.; et al. The Role of Lebanon in the COVID-19 Butterfly Effect: The B.1.398 Example. Viruses 2022, 14, 1640. https://doi.org/10.3390/v14081640
Nour D, Rafei R, Lamarca AP, de Almeida LGP, Osman M, Ismail MB, Mallat H, Berry A, Burfin G, Semanas Q, et al. The Role of Lebanon in the COVID-19 Butterfly Effect: The B.1.398 Example. Viruses. 2022; 14(8):1640. https://doi.org/10.3390/v14081640
Chicago/Turabian StyleNour, Dalal, Rayane Rafei, Alessandra P. Lamarca, Luiz G. P. de Almeida, Marwan Osman, Mohamad Bachar Ismail, Hassan Mallat, Atika Berry, Gwendolyne Burfin, Quentin Semanas, and et al. 2022. "The Role of Lebanon in the COVID-19 Butterfly Effect: The B.1.398 Example" Viruses 14, no. 8: 1640. https://doi.org/10.3390/v14081640