
Insights into Genetic Characteristics and Virological Features of Endemic Avian Influenza A (H9N2) Viruses in Egypt from 2017–2021

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Analysis
2.1.1. Viruses
2.1.2. Amplification of Full Genome and Sequencing
2.1.3. Sequence Analysis and Phylogenetic Tree Construction
2.2. Virological Characteristics of Egyptian H9N2 Viruses
2.2.1. Cells
2.2.2. Viruses
2.2.3. Growth Kinetics of H9N2 Viruses in Mammalian Cells
2.2.4. Replication Rate of H9N2 Viruses in SPF-ECEs
2.2.5. Animal Experiments
2.2.6. Pathogenicity in Chickens and Ducks
2.2.7. Pathogenicity in Mice
2.3. Vaccine Evaluation
2.3.1. Vaccine Preparation
2.3.2. Immunization of SPF Chickens
2.4. Statistical Analysis
3. Results
3.1. Genetic Analysis of Internal Proteins
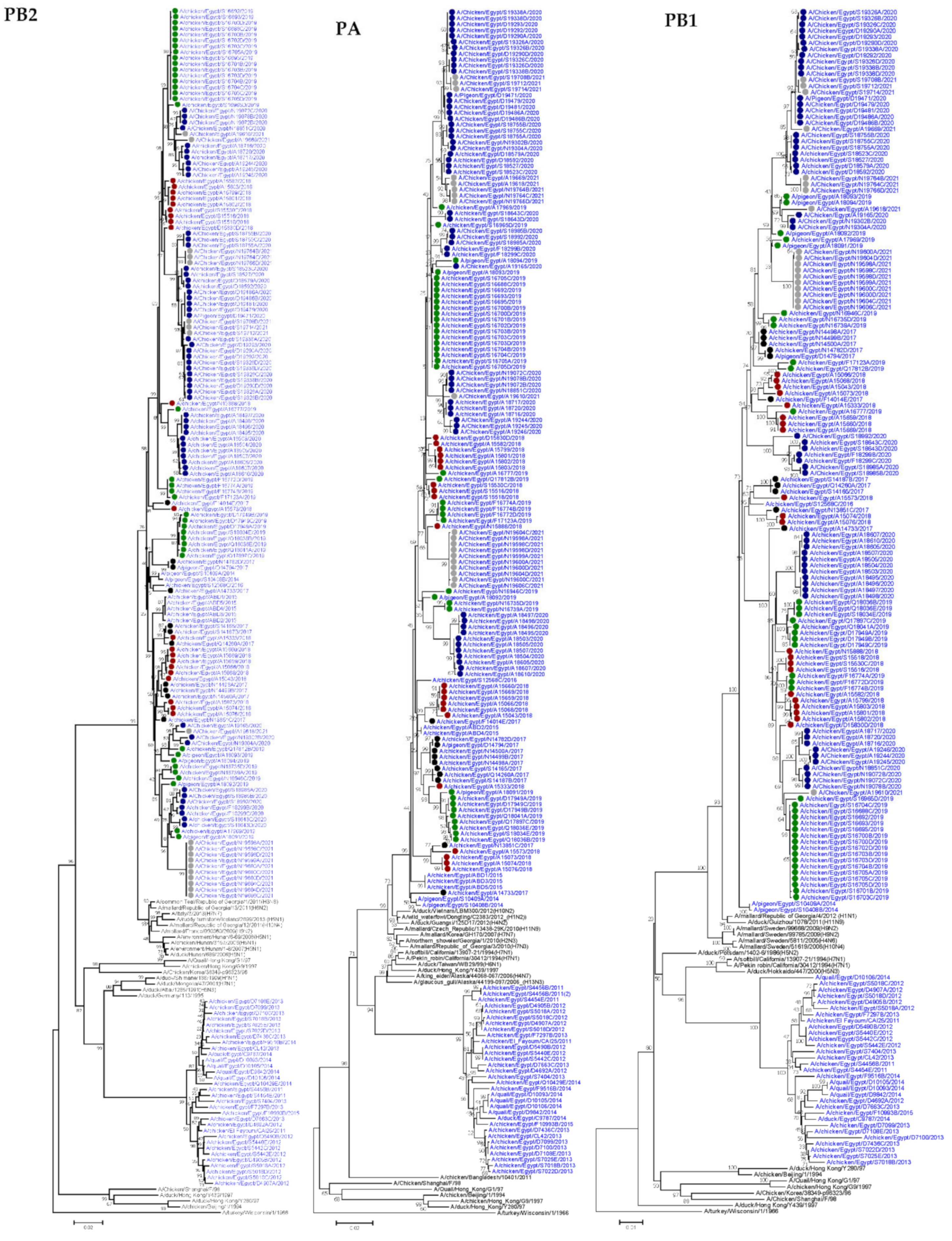
3.1.1. PB2
3.1.2. PB1
3.1.3. PA
3.1.4. HA
3.1.5. NP
3.1.6. NA
3.1.7. M
3.1.8. NS
3.2. Genesis of Egyptian H9N2 Viruses
3.3. Virological Characteristics and Pathogenicity of the Most Dominant Genotypes of Egyptian H9N2 Viruses
3.3.1. Growth Kinetics of Egyptian H9N2 Viruses in Mammalian Cells and SPF-ECEs
3.3.2. Viral Replication and Pathogenicity in Chickens and Ducks
3.3.3. Viral Replication and Pathogenicity in Mice
3.4. Immunogenicity of AI H9N2 Inactivated Vaccine in SPF Chickens
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Homme, P.J.; Easterday, B.C. Avian influenza virus infections. I. Characteristics of influenza A-turkey-Wisconsin-1966 virus. Avian Dis. 1970, 14, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.; Speidel, E.C.; Harris, P.A.; Alexander, D.J. Phylogenetic analysis of influenza A viruses of H9 haemagglutinin subtype. Avian Pathol. 2000, 29, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular characterization of H9N2 influenza viruses: Were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-H.; Song, C.-S. H9N2 avian influenza virus in Korea: Evolution and vaccination. Clin. Exp. Vaccine Res. 2013, 2, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Matrosovich, M.N.; Krauss, S.; Webster, R.G. H9N2 influenza A viruses from poultry in Asia have human virus-like receptor specificity. Virology 2001, 281, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, A.; Monne, I.; Salviato, A.; Valastro, V.; Schivo, A.; Amarin, N.M.; Gonzalez, C.; Ismail, M.M.; Al-Ankari, A.-R.; Al-Blowi, M.H. Phylogeography and evolutionary history of reassortant H9N2 viruses with potential human health implications. J. Virol. 2011, 85, 8413–8421. [Google Scholar] [CrossRef] [Green Version]
- Shahsavandi, S.; Salmanian, A.-H.; Ghorashi, S.A.; Masoudi, S.; Ebrahimi, M.M. Evolutionary characterization of hemagglutinin gene of H9N2 influenza viruses isolated from Asia. Res. Vet. Sci. 2012, 93, 234–239. [Google Scholar] [CrossRef]
- Monne, I.; Cattoli, G.; Mazzacan, E.; Amarin, N.M.; Maaitah, H.M.A.; Al-Natour, M.Q.; Capua, I. Genetic comparison of H9N2 AI viruses isolated in Jordan in 2003. Avian Dis. 2007, 51, 451–454. [Google Scholar] [CrossRef]
- Golender, N.; Panshin, A.; Banet-Noach, C.; Nagar, S.; Pokamunski, S.; Pirak, M.; Tendler, Y.; Davidson, I.; García, M.; Perk, S. Genetic characterization of avian influenza viruses isolated in Israel during 2000–2006. Virus Genes 2008, 37, 289–297. [Google Scholar] [CrossRef]
- Afifi, M.A.; El-Kady, M.F.; Zoelfakar, S.A.; Abddel-Moneim, A.S. Serological surveillance reveals widespread influenza A H7 and H9 subtypes among chicken flocks in Egypt. Trop. Anim. Health Prod. 2013, 45, 687–690. [Google Scholar] [CrossRef]
- El-Zoghby, E.F.; Arafa, A.-S.; Hassan, M.K.; Aly, M.M.; Selim, A.; Kilany, W.H.; Selim, U.; Nasef, S.; Aggor, M.G.; Abdelwhab, E. Isolation of H9N2 avian influenza virus from bobwhite quail (Colinus virginianus) in Egypt. Arch. Virol. 2012, 157, 1167–1172. [Google Scholar] [CrossRef]
- Kandeil, A.; El-Shesheny, R.; Maatouq, A.M.; Moatasim, Y.; Shehata, M.M.; Bagato, O.; Rubrum, A.; Shanmuganatham, K.; Webby, R.J.; Ali, M.A. Genetic and antigenic evolution of H9N2 avian influenza viruses circulating in Egypt between 2011 and 2013. Arch. Virol. 2014, 159, 2861–2876. [Google Scholar] [CrossRef]
- Monne, I.; Hussein, H.A.; Fusaro, A.; Valastro, V.; Hamoud, M.M.; Khalefa, R.A.; Dardir, S.N.; Radwan, M.I.; Capua, I.; Cattoli, G. H9N2 influenza A virus circulates in H5N1 endemically infected poultry population in Egypt. Influenza Other Respir. Viruses 2013, 7, 240–243. [Google Scholar] [CrossRef] [Green Version]
- Shakal, M.; Youssef, Y.; El Zeedy, S.; Ibrahim, S.; Al Baroudi, B. Surveillance on avian influenza H5N1 and H9N2 subtypes in Egypt 2012–2013. Poult. Fish Wildl. Sci. 2013, 2, 2. [Google Scholar]
- Ahmed, Z.; Hussein, H.; Rohaim, M. Efficacy of Composting Poultry Mortality and Farms Wastes with Mixed Respiratory Infection Viruses H9N2 and NDV in Egypt. Glob. Vet. 2013, 11, 177–185. [Google Scholar]
- Dabbour, A. Studies on Avian Influenza Subtype H9 in West Delta Governments in Egypt. Master’s Thesis, Alexandria University, Alexandria, Egypt, 2015. [Google Scholar]
- Arafa, A.-S.; Hagag, N.; Erfan, A.; Mady, W.; El-Husseiny, M.; Adel, A.; Nasef, S. Complete genome characterization of avian influenza virus subtype H9N2 from a commercial quail flock in Egypt. Virus Genes 2012, 45, 283–294. [Google Scholar] [CrossRef]
- Naguib, M.M.; Arafa, A.-S.A.; El-Kady, M.F.; Selim, A.A.; Gunalan, V.; Maurer-Stroh, S.; Goller, K.V.; Hassan, M.K.; Beer, M.; Abdelwhab, E. Evolutionary trajectories and diagnostic challenges of potentially zoonotic avian influenza viruses H5N1 and H9N2 co-circulating in Egypt. Infect. Genet. Evol. 2015, 34, 278–291. [Google Scholar] [CrossRef]
- Kandeil, A.; Hicks, J.T.; Young, S.G.; El Taweel, A.N.; Kayed, A.S.; Moatasim, Y.; Kutkat, O.; Bagato, O.; McKenzie, P.P.; Cai, Z.; et al. Active surveillance and genetic evolution of avian influenza viruses in Egypt, 2016–2018. Emerg. Microbes Infect. 2019, 8, 1370–1382. [Google Scholar] [CrossRef] [Green Version]
- Kayali, G.; Kandeil, A.; El-Shesheny, R.; Kayed, A.S.; Gomaa, M.M.; Maatouq, A.M.; Shehata, M.M.; Moatasim, Y.; Bagato, O.; Cai, Z. Active surveillance for avian influenza virus, Egypt, 2010–2012. Emerg. Infect. Dis. 2014, 20, 542–551. [Google Scholar] [CrossRef]
- Kandeil, A.; El-Shesheny, R.; Maatouq, A.; Moatasim, Y.; Cai, Z.; McKenzie, P.; Webby, R.; Kayali, G.; Ali, M.A. Novel reassortant H9N2 viruses in pigeons and evidence for antigenic diversity of H9N2 viruses isolated from quails in Egypt. J. Gen. Virol. 2017, 98, 548. [Google Scholar] [CrossRef]
- Kandeil, A.; Gomaa, M.R.; Shehata, M.M.; El Taweel, A.N.; Mahmoud, S.H.; Bagato, O.; Moatasim, Y.; Kutkat, O.; Kayed, A.S.; Dawson, P.; et al. Isolation and Characterization of a Distinct Influenza A Virus from Egyptian Bats. J. Virol. 2019, 93, e01059-18. [Google Scholar] [CrossRef] [Green Version]
- Kayed, A.S.; Kandeil, A.; Gomaa, M.R.; El-Shesheny, R.; Mahmoud, S.; Hegazi, N.; Fayez, M.; Sheta, B.; McKenzie, P.P.; Webby, R.J.; et al. Surveillance for avian influenza viruses in wild birds at live bird markets, Egypt, 2014–2016. Influenza Other Respir Viruses 2019, 13, 407–414. [Google Scholar] [CrossRef]
- Peacock, T.H.P.; James, J.; Sealy, J.E.; Iqbal, M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.; Perez, D. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; IRL Press at Oxford University Press: Oxford, UK, 1999; pp. 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Taylor, L.H. OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2014. Available online: https://www.fao.org/fileadmin/templates/rap/files/meetings/2014/140318-reference.pdf (accessed on 17 June 2021).
- World Health Organization (WHO). WHO Manual on Animal Influenza Diagnosis and Surveillance; WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Rolling, T.; Koerner, I.; Zimmermann, P.; Holz, K.; Haller, O.; Staeheli, P.; Kochs, G. Adaptive mutations resulting in enhanced polymerase activity contribute to high virulence of influenza A virus in mice. J. Virol. 2009, 83, 6673–6680. [Google Scholar] [CrossRef] [Green Version]
- Guilligay, D.; Tarendeau, F.; Resa-Infante, P.; Coloma, R.; Crepin, T.; Sehr, P.; Lewis, J.; Ruigrok, R.W.; Ortin, J.; Hart, D.J.; et al. The structural basis for cap binding by influenza virus polymerase subunit PB2. Nat. Struct. Mol. Biol. 2008, 15, 500–506. [Google Scholar] [CrossRef]
- Kuzuhara, T.; Kise, D.; Yoshida, H.; Horita, T.; Murazaki, Y.; Nishimura, A.; Echigo, N.; Utsunomiya, H.; Tsuge, H. Structural basis of the influenza A virus RNA polymerase PB2 RNA-binding domain containing the pathogenicity-determinant lysine 627 residue. J. Biol. Chem. 2009, 284, 6855–6860. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-W.; Chang, S.-C.; Mok, C.-K.; Lo, Y.-L.; Kung, Y.-N.; Huang, J.-H.; Shih, Y.-H.; Wang, J.-Y.; Chiang, C.; Chen, C.-J. Genomic signatures of human versus avian influenza A viruses. Emerg. Infect. Dis. 2006, 12, 1353. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.; Cooper, L.; Xu, X.; Thompson, W.; Krauss, S.; Guan, Y.; Zhou, N.; Klimov, A.; Cox, N.; Webster, R. Molecular changes associated with the transmission of avian influenza a H5N1 and H9N2 viruses to humans. J. Med. Virol. 2002, 66, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Hu, Y.; Chang, G.; Sun, W.; Yang, Y.; Kang, X.; Wu, X.; Zhu, Q. PB1-mediated virulence attenuation of H5N1 influenza virus in mice is associated with PB2. J. Gen. Virol. 2011, 92, 1435–1444. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Y.; Xu, Q.; Tan, Y.; Pu, J.; Yang, H.; Brown, E.G.; Liu, J. Mouse-adapted H9N2 influenza A virus PB2 protein M147L and E627K mutations are critical for high virulence. PLoS ONE 2012, 7, e40752. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Pearce, M.B.; Li, Y.; Wang, J.; Mason, R.J.; Tumpey, T.M.; Wentworth, D.E. Asparagine substitution at PB2 residue 701 enhances the replication, pathogenicity, and transmission of the 2009 pandemic H1N1 influenza A virus. PLoS ONE 2013, 8, e67616. [Google Scholar] [CrossRef] [Green Version]
- Subbarao, E.K.; London, W.; Murphy, B.R. A single amino acid in the PB2 gene of influenza A virus is a determinant of host range. J. Virol. 1993, 67, 1761–1764. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.; Ma, S.; Wang, J.; Gu, C.; Wang, Z.; Shi, J.; Deng, G.; Guan, Y.; Chen, H. Identification of Key Amino Acids in the PB2 and M1 Proteins of H7N9 Influenza Virus That Affect Its Transmission in Guinea Pigs. J. Virol. 2019, 94, e01180-19. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Hatta, M.; Kim, J.H.; Halfmann, P.; Imai, M.; Macken, C.A.; Le, M.Q.; Nguyen, T.; Neumann, G.; Kawaoka, Y. Novel residues in avian influenza virus PB2 protein affect virulence in mammalian hosts. Nat. Commun. 2014, 5, 5021. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Ma, W.; Sun, N.; Huang, L.; Li, Y.; Zeng, Z.; Wen, Y.; Zhang, Z.; Li, H.; Li, Q.; et al. PB2-588 V promotes the mammalian adaptation of H10N8, H7N9 and H9N2 avian influenza viruses. Sci. Rep. 2016, 6, 19474. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Chen, H.; Jiao, P.; Deng, G.; Tian, G.; Li, Y.; Hoffmann, E.; Webster, R.G.; Matsuoka, Y.; Yu, K. Molecular basis of replication of duck H5N1 influenza viruses in a mammalian mouse model. J. Virol. 2005, 79, 12058–12064. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Huang, J.; Chen, Y.; Chen, H.; Li, Q.; He, L.; Hao, X.; Liu, J.; Gu, M.; Hu, J.; et al. Virulence determinants in the PB2 gene of a mouse-adapted H9N2 virus. J. Virol. 2015, 89, 877–882. [Google Scholar] [CrossRef] [Green Version]
- Mok, C.K.P.; Lee, H.H.Y.; Lestra, M.; Nicholls, J.M.; Chan, M.C.W.; Sia, S.F.; Zhu, H.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Amino acid substitutions in polymerase basic protein 2 gene contribute to the pathogenicity of the novel A/H7N9 influenza virus in mammalian hosts. J. Virol. 2014, 88, 3568–3576. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Deng, M.; Lin, Y.; Chang, C.; Shieh, H.K.; Shiau, J.; Huang, C. Characterization of an H5N1 avian influenza virus from Taiwan. Vet. Microbiol. 2007, 124, 193–201. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Z.; Shi, J.; Deng, G.; Kong, H.; Tao, S.; Li, C.; Liu, L.; Guan, Y.; Chen, H. Glycine at Position 622 in PB1 Contributes to the Virulence of H5N1 Avian Influenza Virus in Mice. J. Virol. 2016, 90, 1872–1879. [Google Scholar] [CrossRef] [Green Version]
- Conenello, G.M.; Zamarin, D.; Perrone, L.A.; Tumpey, T.; Palese, P. A single mutation in the PB1-F2 of H5N1 (HK/97) and 1918 influenza A viruses contributes to increased virulence. PLoS Pathog. 2007, 3, 1414–1421. [Google Scholar] [CrossRef]
- Lycett, S.; Ward, M.; Lewis, F.; Poon, A.; Kosakovsky Pond, S.; Brown, A.L. Detection of mammalian virulence determinants in highly pathogenic avian influenza H5N1 viruses: Multivariate analysis of published data. J. Virol. 2009, 83, 9901–9910. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Xu, J.; Shi, J.; Li, Y.; Chen, H. Synergistic Effect of S224P and N383D Substitutions in the PA of H5N1 Avian Influenza Virus Contributes to Mammalian Adaptation. Sci. Rep. 2015, 5, 10510. [Google Scholar] [CrossRef] [Green Version]
- Suguitan, A.L., Jr.; Matsuoka, Y.; Lau, Y.F.; Santos, C.P.; Vogel, L.; Cheng, L.I.; Orandle, M.; Subbarao, K. The multibasic cleavage site of the hemagglutinin of highly pathogenic A/Vietnam/1203/2004 (H5N1) avian influenza virus acts as a virulence factor in a host-specific manner in mammals. J. Virol. 2012, 86, 2706–2714. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Zhang, B.; Shi, J.; Yin, X.; Wang, G.; Cui, P.; Liu, L.; Deng, G.; Jiang, Y.; Li, C.; et al. Amino Acid Mutations A286V and T437M in the Nucleoprotein Attenuate H7N9 Viruses in Mice. J. Virol. 2020, 94, e01530-19. [Google Scholar] [CrossRef] [Green Version]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef] [Green Version]
- Dankar, S.K.; Wang, S.; Ping, J.; Forbes, N.E.; Keleta, L.; Li, Y.; Brown, E.G. Influenza A virus NS1 gene mutations F103L and M106I increase replication and virulence. Virol. J. 2011, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiang, Y.; Jiao, P.; Wang, A.; Zhao, F.; Tian, G.; Wang, X.; Yu, K.; Bu, Z.; Chen, H. The NS1 gene contributes to the virulence of H5N1 avian influenza viruses. J. Virol. 2006, 80, 11115–11123. [Google Scholar] [CrossRef] [Green Version]
- Subbarao, K.; Shaw, M.W. Molecular aspects of avian influenza (H5N1) viruses isolated from humans. Rev. Med. Virol. 2000, 10, 337–348. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Reid, A.H.; Lourens, R.M.; Wang, R.; Jin, G.; Fanning, T.G. Characterization of the 1918 influenza virus polymerase genes. Nature 2005, 437, 889–893. [Google Scholar] [CrossRef]
- Conenello, G.M.; Tisoncik, J.R.; Rosenzweig, E.; Varga, Z.T.; Palese, P.; Katze, M.G. A single N66S mutation in the PB1-F2 protein of influenza A virus increases virulence by inhibiting the early interferon response in vivo. J. Virol. 2011, 85, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Wanitchang, A.; Jengarn, J.; Jongkaewwattana, A. The N terminus of PA polymerase of swine-origin influenza virus H1N1 determines its compatibility with PB2 and PB1 subunits through a strain-specific amino acid serine 186. Virus Res. 2011, 155, 325–533. [Google Scholar] [CrossRef]
- Sorrell, E.M.; Wan, H.; Araya, Y.; Song, H.; Perez, D.R. Minimal molecular constraints for respiratory droplet transmission of an avian–human H9N2 influenza A virus. Proc. Natl. Acad. Sci. USA 2009, 106, 7565–7570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aamir, U.; Wernery, U.; Ilyushina, N.; Webster, R. Characterization of avian H9N2 influenza viruses from United Arab Emirates 2000 to 2003. Virology 2007, 361, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosh, C.; Nagarajan, S.; Behera, P.; Rajukumar, K.; Purohit, K.; Kamal, R.; Murugkar, H.; Gounalan, S.; Pattnaik, B.; Vanamayya, P. Genetic analysis of H9N2 avian influenza viruses isolated from India. Arch. Virol. 2008, 153, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-L.; McKimm-Breschkin, J.L.; Choy, K.-T.; Wong, D.; Cheung, P.; Zhou, J.; Ng, I.H.; Zhu, H.; Webby, R.J.; Guan, Y. Resistance to neuraminidase inhibitors conferred by an R292K mutation in a human influenza virus H7N9 isolate can be masked by a mixed R/K viral population. MBio 2013, 4, e00396-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, J.D.; Gong, L.I.; Baltimore, D. Permissive secondary mutations enable the evolution of influenza oseltamivir resistance. Science 2010, 328, 1272–1275. [Google Scholar] [CrossRef] [Green Version]
- Soubies, S.M.; Volmer, C.; Croville, G.; Loupias, J.; Peralta, B.; Costes, P.; Lacroux, C.; Guérin, J.L.; Volmer, R. Species-specific contribution of the four C-terminal amino acids of influenza A virus NS1 protein to virulence. J. Virol. 2010, 84, 6733–6747. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Shaw, M.; Gregory, V.; Cameron, K.; Lim, W.; Klimov, A.; Subbarao, K.; Guan, Y.; Krauss, S.; Shortridge, K. Avian-to-human transmission of H9N2 subtype influenza A viruses: Relationship between H9N2 and H5N1 human isolates. Proc. Natl. Acad. Sci. USA 2000, 97, 9654–9658. [Google Scholar] [CrossRef] [Green Version]
- Umar, S.; Guerin, J.L.; Ducatez, M.F. Low Pathogenic Avian Influenza and Coinfecting Pathogens: A Review of Experimental Infections in Avian Models. Avian Dis. 2017, 61, 3–15. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Avian Influenza Weekly Update Number 836; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- Pu, J.; Wang, S.; Yin, Y.; Zhang, G.; Carter, R.A.; Wang, J.; Xu, G.; Sun, H.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc. Natl. Acad. Sci. USA 2015, 112, 548–553. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yuan, H.; Gao, R.; Zhang, J.; Wang, D.; Xiong, Y.; Fan, G.; Yang, F.; Li, X.; Zhou, J.; et al. Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: A descriptive study. Lancet 2014, 383, 714–721. [Google Scholar] [CrossRef]
- Hagag, N.M.; Erfan, A.M.; El-Husseiny, M.; Shalaby, A.G.; Saif, M.A.; Tawakol, M.M.; Nour, A.A.; Selim, A.A.; Arafa, A.S.; Hassan, M.K.; et al. Isolation of a Novel Reassortant Highly Pathogenic Avian Influenza (H5N2) Virus in Egypt. Viruses 2019, 11, 565. [Google Scholar] [CrossRef] [Green Version]
- Hassan, K.E.; King, J.; El-Kady, M.; Afifi, M.; Abozeid, H.H.; Pohlmann, A.; Beer, M.; Harder, T. Novel Reassortant Highly Pathogenic Avian Influenza A(H5N2) Virus in Broiler Chickens, Egypt. Emerg. Infect. Dis. 2020, 26, 129–133. [Google Scholar] [CrossRef]
- Nagy, A.; Mettenleiter, T.C.; Abdelwhab, E.M. A brief summary of the epidemiology and genetic relatedness of avian influenza H9N2 virus in birds and mammals in the Middle East and North Africa. Epidemiol. Infect. 2017, 145, 3320–3333. [Google Scholar] [CrossRef] [Green Version]
- Abdel Hamid, H.S.; Ellakany, H.F.; Hussien, H.A.; El-Bestawy, A.R.; Abdel Baky, K.M. Pathogenicity of an Avian Influenza H9N2 Virus isolated From Broiler Chickens in Egypt. Alex. J. Vet. Sci. 2016, 51, 90. [Google Scholar] [CrossRef]
- Awadin, W.; Said, H.; Abdin, S.; El-Sawak, A. Pathological and Molecular Studies on Avian Influenza Virus (H9N2) in Broilers. Asian J. Anim. Vet. Adv. 2018, 13, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Zhang, Y.; Wang, Z.; Wang, J.; Tong, Q.; Wang, M.; Sun, H.; Pu, J.; Liu, C.; Liu, J.; et al. Mouse-adapted H9N2 avian influenza virus causes systemic infection in mice. Virol. J. 2019, 16, 135. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Site | Avirulent | Virulent | Subtypes Tested | Total Egyptian H9N2 (2017–2021) = 173 | References |
|---|---|---|---|---|---|---|
| PB2 | 627 | E | K | H9N2 | E (172), K (1) | [37,39] |
| 147 | M | L | H9N2 | I (155), N/A (18) | [37] | |
| 250 | V | G | H9N2 | V (155), N/A (18) | [37] | |
| 292 | I | V | H7N9 | I (101), V (57), N/A (15) | [40] | |
| 504 | I | V | H1N1 | V (170), N/A (3) | [31] | |
| 588 | A | V | H5N1 H7N9 | A (169), V (4) | [41,42] | |
| 701 | D | N | H1N1, H5N1 | D (173) | [38,43] | |
| 404 | F | L | H9N2 | F (163), N/A (10) | [44] | |
| 591 | Q | K | H7N9 | Q (171), L (2) | [45] | |
| PB1 | 317 | M/V | I | (H5N1, H9N2, H7N2, H7N7), H5N1 | M (165), I (5), N/A (3) | [34,46] |
| 622 | D | G | H5N1 | G (171), S (1), N/A (1) | [47] | |
| PB1-F2 | 66 | N | S | H5N1 | N (120), K (33), N/A (13) 166 | [48] |
| PA | 127 | I | V | H5N1 | V (155), N/A (18) | [49] |
| 383 | N | D | H5N1 | D(173) | [50] | |
| 224 | S | P | H5N1 | S (156), N/A (17) | [50] | |
| 550 | I | L | H1N1 | L (173) | [31] | |
| HA | Cleavage site | Monobasic | Multibasic | H5N1 | KSSR*GLF (4), VSDR*GLF (1), RSSR*GLF (168) | [51] |
| NP | 286 | A | V | H7N9 | A (173) | [52] |
| 437 | T | M | H7N9 | T (173) | ||
| M2 | 64 | P | S/A/F | H5N1 | S (173) | [49] |
| 69 | L | P | H5N1 | P (173) | [49] | |
| NS1 | 42 | A/P | S | H5N1 | S (172), N/A (1) | [53] |
| 92 | D | E | H5N1 | D (170), E (3) | [46] | |
| 103 | F | L | H3N2 | F (173) | [54] | |
| 106 | M | I | H3N2 | M (173) | [54] | |
| 149 | V | A | H5N1 | A (173) | [55] | |
| 189 | D/G | N | H5N1 | D (173) | [56] | |
| NS2 | 31 | M | I | H5N1 | M (173) | [56] |
| 56 | H/L | Y | H5N1 | H (173) | [56] |
| Organs | DPI | S4456B | A17358 |
|---|---|---|---|
| Lung | 3 | 3.6 ± 1 (2/3) * | 1.96 ± 0.65 (2/3) |
| Brain | 8.7(1/3) | ND | |
| Kidney | ND | ND | |
| Liver | 2.55 ± 1 (2/3) | 1(1/3) | |
| Lung | 6 | 1.6 (1/3) | ND |
| Brain | ND | ND | |
| Kidney | ND | ND | |
| Liver | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Sayes, M.; Kandeil, A.; Moatasim, Y.; El Taweel, A.; Rubrum, A.; Kutkat, O.; Kamel, M.N.; Badra, R.; Barakat, A.B.; McKenzie, P.P.; et al. Insights into Genetic Characteristics and Virological Features of Endemic Avian Influenza A (H9N2) Viruses in Egypt from 2017–2021. Viruses 2022, 14, 1484. https://doi.org/10.3390/v14071484
El Sayes M, Kandeil A, Moatasim Y, El Taweel A, Rubrum A, Kutkat O, Kamel MN, Badra R, Barakat AB, McKenzie PP, et al. Insights into Genetic Characteristics and Virological Features of Endemic Avian Influenza A (H9N2) Viruses in Egypt from 2017–2021. Viruses. 2022; 14(7):1484. https://doi.org/10.3390/v14071484
Chicago/Turabian StyleEl Sayes, Mohamed, Ahmed Kandeil, Yassmin Moatasim, Ahmed El Taweel, Adam Rubrum, Omnia Kutkat, Mina Nabil Kamel, Rebecca Badra, Ahmed B. Barakat, Pamela P. McKenzie, and et al. 2022. "Insights into Genetic Characteristics and Virological Features of Endemic Avian Influenza A (H9N2) Viruses in Egypt from 2017–2021" Viruses 14, no. 7: 1484. https://doi.org/10.3390/v14071484