
The Role of Latency-Associated Transcripts in the Latent Infection of Pseudorabies Virus


{kind=link}
Abstract
:1. Introduction
2. The Characteristics of PRV Latent Infection
3. Transcriptional Characteristics of LAT Gene
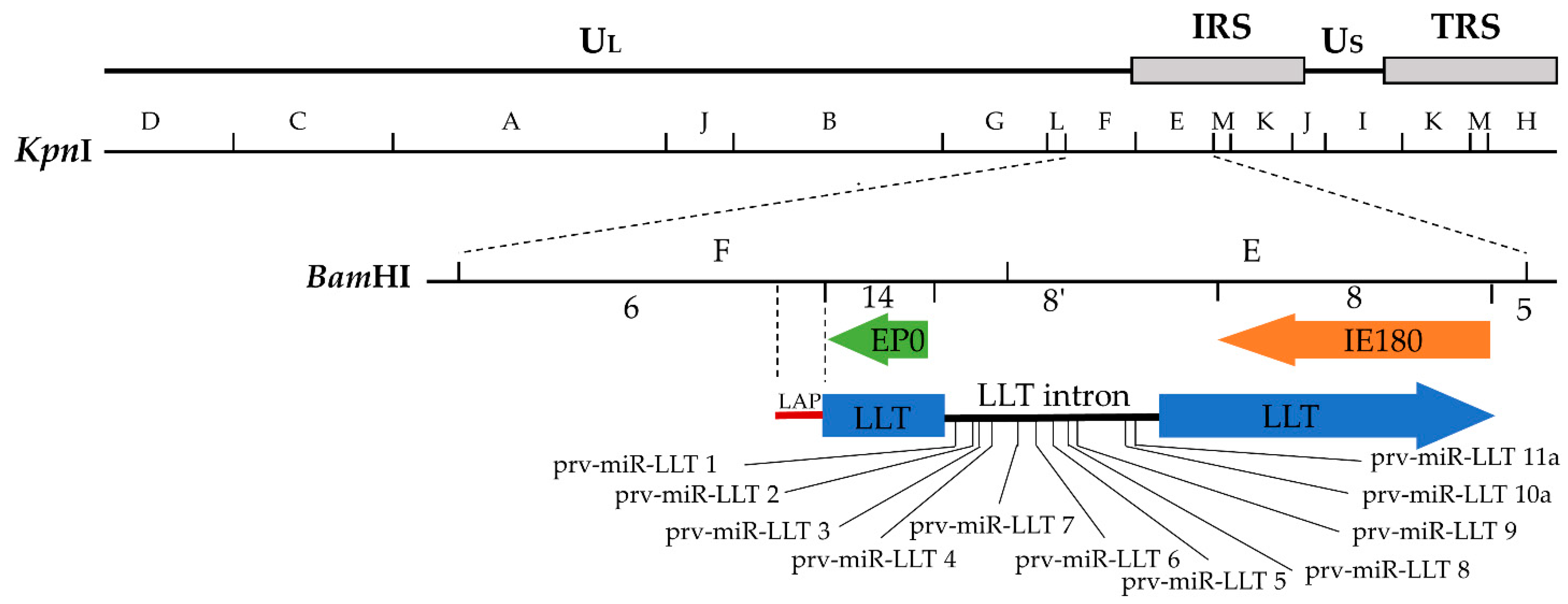
3.1. Transcriptional Region and Sizes of the LAT Gene
3.2. The Structure and Function of LAT Promoter
4. The Role of LAT Gene in Latent Infection
4.1. MicroRNAs Transcribed by the LAT Gene and Host Cell
4.2. Other Non-Coding RNAs Transcribed by LAT Gene
4.3. LAT Encoding Protein in Latent Infection
5. Effect of Pseudorabies Vaccine on Virus Latent Infection
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hengartner, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular Biology of Pseudorabies Virus: Impact on Neurovirology and Veterinary Medicine. Microbiol. Mol. Biol. 2005, 69, 462–500. [Google Scholar] [CrossRef] [Green Version]
- Klupp, B.G.; Hengartner, C.J.; Mettenleiter, T.C.; Enquist, L.W. Complete, Annotated Sequence of the Pseudorabies Virus Genome. J. Virol. 2004, 78, 17. [Google Scholar] [CrossRef] [Green Version]
- Hugoson, G.; Rockborn, G. On the Occurrence of Pseudorabies in Sweden II. An Outbreak in Dogs Caused by Feeding Abattoir Offal. Zentralbl Vet. B 2010, 19, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Fico, R.; Banks, M.; Tamba, M.; Calzetta, G. Isolation and Characterisation of an Aujeszky’s Disease Virus Naturally Infecting a Wild Boar (Sus Scrofa). Vet. Microbiol. 1997, 55, 141–146. [Google Scholar] [CrossRef]
- Cheng, Z.; Kong, Z.; Liu, P.; Fu, Z.; Zhang, J.; Liu, M.; Shang, Y. Natural Infection of a Variant Pseudorabies Virus Leads to Bovine Death in China. Transbound. Emerg. Dis. 2020, 67, 518–522. [Google Scholar] [CrossRef]
- Kaneko, C.; Kaneko, Y.; Sudaryatma, P.E.; Mekata, H.; Kirino, Y.; Yamaguchi, R.; Okabayashi, T. Pseudorabies Virus Infection in Hunting Dogs in Oita, Japan: Report from a Prefecture Free from Aujeszky’s Disease in Domestic Pigs. J. Vet. Med. Sci. 2021, 83, 680–684. [Google Scholar] [CrossRef]
- Marcaccini, A.; López Peña, M.; Quiroga, M.I.; Bermúdez, R.; Nieto, J.M.; Alemañ, N. Pseudorabies Virus Infection in Mink: A Host-Specific Pathogenesis. Vet. Immunol. Immunopathol. 2008, 124, 264–273. [Google Scholar] [CrossRef]
- Laval, K.; Vernejoul, J.B.; Van Cleemput, J.; Koyuncu, O.O.; Enquist, L.W. Virulent Pseudorabies Virus Infection Induces a Specific and Lethal Systemic Inflammatory Response in Mice. J. Virol. 2018, 92, e01614-18. [Google Scholar] [CrossRef] [Green Version]
- Di Marco Lo Presti, V.; Moreno, A.; Castelli, A.; Ippolito, D.; Aliberti, A.; Amato, B.; Vitale, M.; Fiasconaro, M.; Pruiti Ciarello, F. Retrieving Historical Cases of Aujeszky’s Disease in Sicily (Italy): Report of a Natural Outbreak Affecting Sheep, Goats, Dogs, Cats and Foxes and Considerations on Critical Issues and Perspectives in Light of the Recent EU Regulation 429/2016. Pathogens 2021, 10, 1301. [Google Scholar] [CrossRef]
- Laval, K.; Enquist, L.W. The Neuropathic Itch Caused by Pseudorabies Virus. Pathogens 2020, 9, 254. [Google Scholar] [CrossRef] [Green Version]
- Sehl, J.; Teifke, J.P. Comparative Pathology of Pseudorabies in Different Naturally and Experimentally Infected Species—A Review. Pathogens 2020, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Gutekunst, D.C.; Pirtle, E.C.; Miller, L.D.; Stewart, W.C. Isolation of Pseudorabies Virus from Trigeminal Ganglia of a Latently Infected Sow. Am. J. Vet. Res. 1980, 41, 1315–1316. [Google Scholar] [PubMed]
- Brown, T.M.; Osorio, F.A.; Rock, D.L. Detection of Latent Pseudorabies Virus in Swine Using in Situ Hybridization. Vet. Microbiol. 1990, 24, 273–280. [Google Scholar] [CrossRef]
- Romero, C.H.; Meade, P.N.; Homer, B.L.; Shultz, J.E.; Lollis, G. Potential Sites of Virus Latency Associated with Indigenous Pseudorabies Viruses in Feral Swine. J. Wildl. Dis. 2003, 39, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Brittle, E.E.; Reynolds, A.E.; Enquist, L.W. Two Modes of Pseudorabies Virus Neuroinvasion and Lethality in Mice. J. Virol. 2004, 78, 12951–12963. [Google Scholar] [CrossRef] [Green Version]
- Seiichi, T.; Takashi, I.; Masashi, S.; Kazuaki, M. Acetylcholine Reactivates Latent Pseudorabies Virus in Mice. J. Virol. Methods 1998, 70, 103–106. [Google Scholar] [CrossRef]
- Seiichi, T.; Kazuaki, M. Activation of Latent Pseudorabies Virus Infection in Mice Treated with Acetylcholine. Exp. Anim. 2002, 51, 407–709. [Google Scholar] [CrossRef] [Green Version]
- Osorio, F.A.; Rock, D.L. A Murine Model of Pseudorabies Virus Latency. Microb. Pathog. 1992, 12, 39–46. [Google Scholar] [CrossRef]
- Seiichi, T.; Kazuaki, M. Analysis of the Mechanism of Reactivation of Latently Infecting Pseudorabies Virus by Acetylcholine. J. Vet. Med. Sci. 2014, 76, 719–722. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liang, W.; Liu, Q.; Zhao, T.; Zhu, H.; Hua, L.; Peng, Z.; Tang, X.; Stratton, C.; Zhou, D.; et al. Epidemiological and Genetic Characteristics of Swine Pseudorabies Virus in Mainland China between 2012 and 2017. PeerJ 2018, 6, e5785. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Jin, Y.; Hou, C.; Li, X.; Zhao, L.; Wang, Z.; Chen, H. Seroprevalence Investigation and Genetic Analysis of Pseudorabies Virus within Pig Populations in Henan Province of China during 2018–2019. Infect. Genet. Evol. 2021, 92, 104835. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Ren, H.; Gu, J.; Wang, J.; Jiang, L.; Gao, S. The Epidemiological Analysis of Pseudorabies Virus and Pathogenicity of the Variant Strain in Shandong Province. Front. Vet. Sci. 2022, 9, 806824. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Tan, L.; Wang, C.; He, S.; Fang, L.; Wang, Z.; Zhong, Y.; Zhang, K.; Liu, D.; Yang, Q.; et al. Serological Investigation and Genetic Characteristics of Pseudorabies Virus in Hunan Province of China from 2016 to 2020. Front. Vet. Sci. 2021, 8, 762326. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Luo, Y.; Wang, C.H.; Yuan, J.; Li, N.; Song, K.; Qiu, H.J. Control of Swine Pseudorabies in China: Opportunities and Limitations. Vet. Microbiol. 2016, 183, 119–124. [Google Scholar] [CrossRef]
- Tong, W.; Liu, F.; Zheng, H.; Liang, C.; Zhou, Y.; Jiang, Y.; Shan, T.; Gao, F.; Li, G.; Tong, G. Emergence of a Pseudorabies Virus Variant with Increased Virulence to Piglets. Vet. Microbiol. 2015, 181, 236–240. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Peng, J.; Tian, Z.; Zhao, H.; Li, N.; Liu, Y.; Chen, J.; Leng, C.; Sun, Y.; Chang, D.; et al. Pseudorabies Virus Variant in Bartha-K61-Vaccinated Pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef]
- Yu, Z.; Tong, W.; Zheng, H.; Li, L.; Li, G.; Gao, F.; Wang, T.; Liang, C.; Ye, C.; Wu, J.; et al. Variations in Glycoprotein B Contribute to Immunogenic Difference between PRV Variant JS-2012 and Bartha-K61. Vet. Microbiol. 2017, 208, 97–105. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A Novel Human Acute Encephalitis Caused by Pseudorabies Virus Variant Strain. Clin. Infect. Dis. 2021, 73, e3690–e3700. [Google Scholar] [CrossRef]
- Ai, J.; Weng, S.; Cheng, Q.; Cui, P.; Li, Y.; Wu, H.; Zhu, Y.; Xu, B.; Zhang, W. Human Endophthalmitis Caused By Pseudorabies Virus Infection, China, 2017. Emerg. Infect. Dis. 2018, 24, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Yuan, H.; Liu, L.; Li, H.; Wang, S.; Zhao, W.; Wu, Y.; Wang, P.; Hu, Y.; Han, J.; et al. Pseudorabies Virus Encephalitis in Humans: A Case Series Study. J. Neurovirol. 2020, 26, 556–564. [Google Scholar] [CrossRef]
- Wang, Y.; Nian, H.; Li, Z.; Wang, W.; Wang, X.; Cui, Y. Human Encephalitis Complicated with Bilateral Acute Retinal Necrosis Associated with Pseudorabies Virus Infection: A Case Report. Int. J. Infect. Dis. 2019, 89, 51–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Guan, H.; Li, C.; Li, Y.; Wang, S.; Zhao, X.; Zhao, Y.; Liu, Y. Characteristics of Human Encephalitis Caused by Pseudorabies Virus: A Case Series Study. Int. J. Infect. Dis. 2019, 87, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Han, H.; Wang, H.; Cui, Y.; Liu, H.; Ding, S. A Case of Human Viral Encephalitis Caused by Pseudorabies Virus Infection in China. Front. Neurol. 2019, 10, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Liu, X.; Yuan, D.; Li, R.; Lu, J.; Li, X.; Tian, K.; Dai, E. Dynamic Cerebrospinal Fluid Analyses of Severe Pseudorabies Encephalitis. Transbound. Emerg. Dis. 2019, 66, 2562–2565. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wang, J.; Peng, X.Y. Bilateral Necrotizing Retinitis Following Encephalitis Caused by the Pseudorabies Virus Confirmed by Next-Generation Sequencing. Ocul. Immunol. Inflamm. 2021, 29, 922–925. [Google Scholar] [CrossRef]
- Yan, W.; Hu, Z.; Zhang, Y.; Wu, X.; Zhang, H. Case Report: Metagenomic Next-Generation Sequencing for Diagnosis of Human Encephalitis and Endophthalmitis Caused by Pseudorabies Virus. Front. Med. 2022, 8, 753988. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tao, X.; Fei, M.; Chen, J.; Guo, W.; Li, P.; Wang, J. Human Encephalitis Caused by Pseudorabies Virus Infection: A Case Report. J. Neurovirol. 2020, 26, 442–448. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Tong, F.; Tian, M.; Li, M.; Wang, L.; Zou, Y.; Duan, J.; Bu, H.; He, J. Human Encephalitis Complicated With Ocular Symptoms Associated With Pseudorabies Virus Infection: A Case Report. Front. Neurol. 2022, 13, 878007. [Google Scholar]
- Zhou, Y.; Nie, C.; Wen, H.; Long, Y.; Zhou, M.; Xie, Z.; Hong, D. Human Viral Encephalitis Associated with Suid Herpesvirus 1. Neurol. Sci. 2022, 43, 2681–2692. [Google Scholar] [CrossRef]
- Motovski, A.; Kunev, Z.; Stoichev, P. Epizootic Process on a Farm Chronically Infected with Aujeszky’s Disease. Vet.-Meditsinski Nauki 1977, 14, 16–21. [Google Scholar]
- Zhang, Y.; Zeng, L.-S.; Wang, J.; Cai, W.-Q.; Cui, W.; Song, T.-J.; Peng, X.-C.; Ma, Z.; Xiang, Y.; Cui, S.-Z.; et al. Multifunctional Non-Coding RNAs Mediate Latent Infection and Recurrence of Herpes Simplex Viruses. Infect. Drug Resist. 2021, 14, 5335–5349. [Google Scholar] [CrossRef] [PubMed]
- Smith, G. Herpesvirus Transport to the Nervous System and Back Again. Annu. Rev. Microbiol. 2012, 66, 153–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyuncu, O.O.; Hogue, L.B.; Enquist, L.W. Virus Infections in the Nervous System. Cell Host Microbe 2013, 13, 379–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, C.M. Repression of Viral Transcription during Herpes Simplex Virus Latency. Microbiology 2000, 81, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.J.; Wong, M.; Huang, C.; Chang, T.J. Suppression of Promoter Activity of the LAT Gene by IE180 of Pseudorabies Virus. Virus Genes 2002, 13, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Rock, D.L.; Hagemoser, W.A.; Osorio, F.A.; McAllister, H.A. Transcription from the Pseudorabies Virus Genome during Latent Infection. Brief Report. Arch. Virol. 1988, 98, 99–106. [Google Scholar] [CrossRef]
- Rziha, H.J.; Mettenleiter, T.C.; Ohlinger, V.; Wittmann, G. Herpesvirus (Pseudorabies Virus) Latency in Swine: Occurrence and Physical State of Viral DNA in Neural Tissues. Virology 1986, 155, 600–613. [Google Scholar] [CrossRef]
- Tanaka, S.; Mannen, K. Effect of Mild Stress in Mice Latently Infected Pseudorabies Virus. Exp. Anim. 2003, 52, 383–386. [Google Scholar] [CrossRef] [Green Version]
- Szpara, M.L.; Kobiler, O.; Enquist, L.W. A Common Neuronal Response to Alphaherpesvirus Infection. J. Neuroimmune Pharmacol. 2010, 5, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Shu, M.; Du, T.; Zhou, G.; Roizman, B. Role of Activating Transcription Factor 3 in the Synthesis of Latency-Associated Transcript and Maintenance of Herpes Simplex Virus 1 in Latent State in Ganglia. Proc. Natl. Acad. Sci. USA 2015, 112, E5420–E5426. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Tang, J. Evasion of I Interferon-Mediated Innate Immunity by Pseudorabies Virus. Front. Microbiol. 2021, 12, 801257. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Scherba, G. Expression of the Pseudorabies Virus Latency-Associated Transcript Gene during Productive Infection of Cultured Cells. J. Virol. 1999, 73, 9781–9788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priola, S.A.; Gustafson, D.P.; Wagner, E.K.; Stevens, J.G. A Major Portion of the Latent Pseudorabies Virus Genome Is Transcribed in Trigeminal Ganglia of Pigs. J. Virol. 1990, 64, 4755–4760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priola, S.A.; Stevens, J.G. The 5′ and 3′ Limits of Transcription in the Pseudorabies Virus Latency Associated Transcription Unit. Virology 1991, 182, 852–856. [Google Scholar] [CrossRef]
- Cheung, A.K. Cloning of the Latency Gene and the Early Protein 0 Gene of Pseudorabies Virus. J. Virol. 1991, 65, 5260–5271. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.K. Detection of Pseudorabies Virus Transcripts in Trigeminal Ganglia of Latently Infected Swine. J. Virol. 1989, 63, 2908–2913. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.K.; Smith, T.A. Analysis of the Latency-Associated Transcript/UL1-3.5 Gene Cluster Promoter Complex of Pseudorabies Virus. Arch. Virol. 1999, 144, 381–391. [Google Scholar] [CrossRef]
- Jin, L.; Schnitzlein, W.M.; Scherba, G. Identification of the Pseudorabies Virus Promoter Required for Latency-Associated Transcript Gene Expression in the Natural Host. J. Virol. 2000, 74, 6333–6338. [Google Scholar] [CrossRef] [Green Version]
- Ou, C.J.; Chen, Y.; Huang, C. Cloning and Characterization of the Pseudorabies Virus Latency-Associated Transcript Promoter. Taiwan Vet. J. 2002, 28, 252–259. [Google Scholar]
- Taharaguchi, S.; Yoshino, S.; Amagai, K.; Ono, E. The Latency-Associated Transcript Promoter of Pseudorabies Virus Directs Neuron-Specific Expression in Trigeminal Ganglia of Transgenic Mice. J. Gen. Virol. 2003, 84, 2015–2022. [Google Scholar] [CrossRef]
- Taharaguchi, S.; Kobayashi, T.; Yoshino, S.; Ono, E. Analysis of Regulatory Functions for the Region Located Upstream from the Latency-Associated Transcript (LAT) Promoter of Pseudorabies Virus in Cultured Cells. Vet. Microbiol. 2002, 85, 197–208. [Google Scholar] [CrossRef]
- Papageorgiou, K.; Grivas, I.; Chiotelli, M.; Theodoridis, A.; Panteris, E.; Papadopoulos, D.; Petridou, E.; Papaioannou, N.; Nauwynck, H.; Kritas, S.K. Age-Dependent Invasion of Pseudorabies Virus into Porcine Central Nervous System via Maxillary Nerve. Pathogens 2022, 11, 157. [Google Scholar] [CrossRef] [PubMed]
- Tham, K.M.; Motha, M.X.J.; Horner, G.W.; Ralston, J.C. Polymerase Chain Reaction Amplification of Latent Aujeszky’s Disease Virus in Dexamethasone Treated Pigs. Arch. Virol. 1994, 136, 197–205. [Google Scholar] [CrossRef]
- Tycowski, K.T.; Guo, Y.E.; Lee, N.; Moss, W.N.; Vallery, T.K.; Xie, M.; Steitz, J.A. Viral Noncoding RNAs: More Surprises. Genes Dev. 2015, 29, 567–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timoneda, O.; Núñez-Hernández, F.; Balcells, I.; Muñoz, M.; Castelló, A.; Vera, G.; Pérez, L.J.; Egea, R.; Mir, G.; Córdoba, S.; et al. The Role of Viral and Host MicroRNAs in the Aujeszky’s Disease Virus during the Infection Process. PLoS ONE 2014, 9, e86965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, W.; Liu, Z.F. Long Non-Coding RNAs: Novel Players in Regulation of Immune Response Upon Herpesvirus Infection. Front. Immunol. 2018, 9, 761. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.T.; Wang, L.; Wang, H.; Tang, F.R.; Cai, W.Q.; Sethi, G.; Xin, H.W.; Ma, Z. Insights into Biological Role of LncRNAs in Epithelial-Mesenchymal Transition. Cells 2019, 8, 1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.F.D.; Jones, C. Small Non-Coding RNAs Encoded within the Herpes Simplex Virus Type 1 Latency Associated Transcript (LAT) Cooperate with the Retinoic Acid Inducible Gene I (RIG-I) to Induce Beta-Interferon Promoter Activity and Promote Cell Survival. Virus Res. 2013, 175, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Silva, M.S.E.; Jaber, T.; Vitvitskaia, O.; Li, S.; Henderson, G.; Jones, C. Two Small RNAs Encoded within the First 1.5 Kilobases of the Herpes Simplex Virus Type 1 Latency-Associated Transcript Can Inhibit Productive Infection and Cooperate To Inhibit Apoptosis. J. Virol. 2009, 83, 9131–9139. [Google Scholar] [CrossRef] [Green Version]
- Cliffe, A.R.; Garber, D.A.; Knipe, D.M. Transcription of the Herpes Simplex Virus Latency-Associated Transcript Promotes the Formation of Facultative Heterochromatin on Lytic Promoters. J. Virol. 2009, 83, 8182–8190. [Google Scholar] [CrossRef] [Green Version]
- Grey, F. Role of MicroRNAs in Herpesvirus Latency and Persistence. J. Gen. Virol. 2015, 96, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Jurak, L.; Griffiths, A.; Coen, D.M. Mammalian Alphaherpesvirus MiRNAs. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2011, 1809, 641–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umbach, J.L.; Kramer, M.F.; Jurak, L.; Karnowski, H.W.; Coen, D.M.; Cullen, B.R. MicroRNAs Expressed by Herpes Simplex Virus 1 during Latent Infection Regulate Viral MRNAs. Nature 2008, 454, 780–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Brown, D.; Osorio, N.; Hsiang, C.; BenMohamed, L.; Wechsler, S.L. Increased Neurovirulence and Reactivation of the Herpes Simplex Virus Type 1 Latency-Associated Transcript (LAT)-Negative Mutant DLAT2903 with a Disrupted LAT MiR-H2. J. Neurovirol. 2016, 22, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Kincaid, R.P.; Sullivan, C.S. Virus-Encoded MicroRNAs: An Overview and a Look to the Future. PLoS Pathog. 2012, 8, e1003018. [Google Scholar] [CrossRef] [Green Version]
- Anselmo, A.; Flori, L.; Jaffrezic, F.; Rutigliano, T.; Cecere, M.; Cortes-Perez, N.; Lefevre, F.; Rogel-Gaillard, C.; Giuffra, E. Co-Expression of Host and Viral MicroRNAs in Porcine Dendritic Cells Infected by the Pseudorabies Virus. PLoS ONE 2011, 6, e17374. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Q.; Chen, Q.J.; He, H.B.; Chen, D.S.; Chen, L.L.; Chen, H.C.; Liu, Z.F. Pseudorabies Virus Infected Porcine Epithelial Cell Line Generates a Diverse Set of Host MicroRNAs and a Special Cluster of Viral MicroRNAs. PLoS ONE 2012, 7, e30988. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Patel, A.; Krause, P.R. Novel Less-Abundant Viral MicroRNAs Encoded by Herpes Simplex Virus 2 Latency-Associated Transcript and Their Roles in Regulating ICP34.5 and ICP0 MRNAs. J. Virol. 2009, 83, 1433–1442. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, W.E.; Brough, D.E.; Kovesdi, I.; DeLuca, N.A. Efficient Activation of Viral Genomes by Levels of Herpes Simplex Virus ICP0 Insufficient To Affect Cellular Gene Expression or Cell Survival. J. Virol. 2001, 75, 3391–3403. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Bertke, A.S.; Patel, A.; Wang, K.; Cohen, J.I.; Krause, P.R. An Acutely and Latently Expressed Herpes Simplex Virus 2 Viral MicroRNA Inhibits Expression of ICP34.5, a Viral Neurovirulence Factor. Proc. Natl. Acad. Sci. USA 2008, 105, 10931–10936. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Zheng, H.; Tong, W.; Li, G.X.; Tian, Q.; Liang, C.; Li, L.W.; Zheng, X.C.; Tong, G.Z. Identification and Analysis of Novel Viral and Host Dysregulated MicroRNAs in Variant Pseudorabies Virus-Infected PK15 Cells. PLoS ONE 2016, 11, e0151546. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, G.; Zhang, Y.; Yang, X.; Liu, H.; Chang, H.; Wang, X.; Zhao, J.; Wang, C.; Chen, L. MicroRNA Analysis in Mouse Neuro-2a Cells after Pseudorabies Virus Infection. J. Neurovirol. 2017, 23, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Saba, R.; Sorensen, D.L.; Booth, S.A. MicroRNA-146a: A Dominant, Negative Regulator of the Innate Immune Response. Front. Immunol. 2014, 5, 578. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-ΚB-Dependent Induction of MicroRNA MiR-146, an Inhibitor Targeted to Signaling Proteins of Innate Immune Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.; Gao, Y.; Liu, X.; Bai, J.; Jiang, P.; Wang, X. Long Non-Coding RNA LNC_000641 Regulates Pseudorabies Virus Replication. Vet. Res. 2021, 52, 1–13. [Google Scholar] [CrossRef]
- Peng, W.; Vitvitskaia, O.; Carpenter, D.; Wechsler, S.L.; Jones, C. Identification of Two Small RNAs within the First 1.5-Kb of the Herpes Simplex Virus Type 1–Encoded Latency-Associated Transcript. J. Neurovirol. 2008, 14, 41–52. [Google Scholar] [CrossRef]
- Ahmed, M.; Lock, M.; Miller, C.G.; Fraser, N.W. Regions of the Herpes Simplex Virus Type 1 Latency-Associated Transcript That Protect Cells from Apoptosis In Vitro and Protect Neuronal Cells In Vivo. J. Virol. 2002, 76, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Nicoll, M.P.; Proenc, J.T.; Efstathiou, S. The Molecular Basis of Herpes Simplex Virus Latency. FEMS Microbiol. Rev. 2012, 36, 684–705. [Google Scholar] [CrossRef]
- Allen, S.J.; Rhode-Kurnow, A.; Mott, K.R.; Jiang, X.; Carpenter, D.; Rodriguez-Barbosa, J.I.; Jones, C.; Wechsler, S.L.; Ware, C.F.; Ghiasi, H. Interactions between Herpesvirus Entry Mediator (TNFRSF14) and Latency-Associated Transcript during Herpes Simplex Virus 1 Latency. J. Virol. 2014, 88, 1961–1971. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, D.; Henderson, G.; Hsiang, C.; Osorio, N.; BenMohamed, L.; Jones, C.; Wechsler, S.L. Introducing Point Mutations into the ATGs of the Putative Open Reading Frames of the HSV-1 Gene Encoding the Latency Associated Transcript (LAT) Reduces Its Anti-Apoptosis Activity. Microb. Pathog. 2008, 44, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.K.; Lilley, C.E.; Latchman, D.S.; Coffin, R.S. A Protein Encoded by the Herpes Simplex Virus (HSV) Type 1 2-Kilobase Latency-Associated Transcript Is Phosphorylated, Localized to the Nucleus, and Overcomes the Repression of Expression from Exogenous Promoters When Inserted into the Quiescent HSV Genome. J. Virol. 2002, 76, 4056–4067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.K.; Gough, G.; Latchman, D.S.; Coffin, R.S. Herpes Simplex Virus Latency-Associated Transcript Encodes a Protein Which Greatly Enhances Virus Growth, Can Compensate for Deficiencies in Immediate-Early Gene Expression, and Is Likely To Function during Reactivation from Virus Latency. J. Virol. 1999, 73, 6618–6625. [Google Scholar] [CrossRef] [Green Version]
- Mock, R.E.; Crandell, R.A.; Mesfin, G.M. Induced Latency in Pseudorabies Vaccinated Pigs. Can. J. Comp. Med. Rev. Can. Med. Comp. 1981, 45, 56–59. [Google Scholar]
- Mengeling, W.L. Virus Reactivation in Pigs Latently Infected with a Thymidine Kinase Negative Vaccine Strain of Pseudorabies Virus. Arch. Virol. 1991, 120, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Volz, D.M.; Lager, K.M.; Mengeling, W.L. Latency of a Thymidine Kinase-Negative Pseudorabies Vaccine Virus Detected by the Polymerase Chain Reaction. Arch. Virol. 1992, 122, 341–348. [Google Scholar] [CrossRef] [PubMed]
- van Oirschot, J.T.; Gielkens, A.L. In Vivo and in Vitro Reactivation of Latent Pseudorabies Virus in Pigs Born to Vaccinated Sows. Am. J. Vet. Res. 1984, 45, 567–571. [Google Scholar]
- McCaw, M.B.; Osorio, F.A.; Wheeler, J.; Xu, J.; Erickson, G.A. Effect of Maternally Acquired Aujeszky’s Disease (Pseudorabies) Virus-Specific Antibody in Pigs on Establishment of Latency and Seroconversion to Differential Glycoproteins after Low Dose Challenge. Vet. Microbiol. 1997, 55, 91–98. [Google Scholar] [CrossRef]
- Wittmann, G.; Ohlinger, V.; Rziha, J.H. Occurrence and Reactivation of Latent Aujeszky’s Disease Virus Following Challenge in Previously Vaccinated Pigs. Arch. Virol. 1983, 75, 29–41. [Google Scholar] [CrossRef]
- Schoenbaum, M.A.; Beran, G.W.; Murphy, D.P. Pseudorabies Virus Latency and Reactivation in Vaccinated Swine. Am. J. Vet. Res. 1990, 51, 334–338. [Google Scholar]
- Lu, J.J.; Yuan, W.Z.; Zhu, Y.P.; Hou, S.H.; Wang, X.J. Latent Pseudorabies Virus Infection in Medulla Oblongata from Quarantined Pigs. Transbound. Emerg. Dis. 2021, 68, 543–551. [Google Scholar] [CrossRef]
- Schang, L.M.; Kutish, G.F.; Osorio, F.A. Correlation between Precolonization of Trigeminal Ganglia by Attenuated Strains of Pseudorabies Virus and Resistance to Wild-Type Virus Latency. J. Virol. 1994, 68, 8470–8476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiry, E.; Meurens, F.; Muylkens, B.; McVoy, M.; Gogev, S.; Thiry, J.; Vanderplasschen, A.; Epstein, A.; Keil, G.; Schynts, F. Recombination in Alphaherpesviruses. Rev. Med. Virol. 2005, 15, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Thiry, E.; Muylkens, B.; Meurens, F.; Gogev, S.; Thiry, J.; Vanderplasschen, A.; Schynts, F. Recombination in the Alphaherpesvirus Bovine Herpesvirus 1. Vet. Microbiol. 2006, 113, 171–177. [Google Scholar] [CrossRef]
- Meurens, F.; Schynts, F.; Keil, G.M.; Muylkens, B.; Vanderplasschen, A.; Gallego, P.; Thiry, E. Superinfection Prevents Recombination of the Alphaherpesvirus Bovine Herpesvirus 1. J. Virol. 2004, 78, 3872–3879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, R.K.; Sussman, M.D.; Vilnis, A.; Thacker, B.J. Recent Developments in Latency and Recombination of Aujeszky’s Disease (Pseudorabies) Virus. Vet. Microbiol. 1997, 55, 13–27. [Google Scholar] [CrossRef]
- Mahjoub, N.; Dhorne-Pollet, S.; Fuchs, W.; Ahanda, E.; Lange, E.; Klupp, B.; Arya, A.; Loveland, J.E.; Lefevre, F.; Mettenleiter, T.C.; et al. A 2.5-Kilobase Deletion Containing a Cluster of Nine MicroRNAs in the Latency-Associated-Transcript Locus of the Pseudorabies Virus Affects the Host Response of Porcine Trigeminal Ganglia during Established Latency. J. Virol. 2015, 89, 428–442. [Google Scholar] [CrossRef] [Green Version]
- Mott, K.R. The Bovine Herpesvirus-1 LR ORF2 Is Critical for This Gene’s Ability to Restore the High Wild-Type Reactivation Phenotype to a Herpes Simplex Virus-1 LAT Null Mutant. J. Gen. Virol. 2003, 84, 2975–2985. [Google Scholar] [CrossRef]
- Harrison, K.S.; Zhu, L.; Thunuguntla, P.; Jones, C. Herpes Simplex Virus 1 Regulates β-Catenin Expression in TG Neurons during the Latency-Reactivation Cycle. PLoS ONE 2020, 15, e0230870. [Google Scholar] [CrossRef]
- Thiery, R.; Boutin, P.; Arnauld, C.; Jestin, A. Pseudorabies Virus Latency: A Quantitative Approach by Polymerase Chain Reaction. Acta Vet. Hung. 1994, 42, 277–287. [Google Scholar]
- White, A.K.; Ciacci-Zanella, J.; Galeota, J.; Ele, S.; Osorio Fernando, A. Comparison of the Abilities of Serologic Tests to Detect Pseudorabies-Infected Pigs during the Latent Phase of Infection. Am. J. Vet. Res. 1996, 57, 608–611. [Google Scholar]
- Tu, F.; Zhang, Y.; Xu, S.; Yang, X.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Yang, H. Detection of Pseudorabies Virus with a Real-time Recombinase-aided Amplification Assay. Transbound. Emerg. Dis. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.A.; Eo, S.K.; Aleyas, A.G.; Park, S.O.; Lee, J.H.; Chae, J.S.; Cho, J.G.; Song, H.J. Molecular Survey of Latent Pseudorabies Virus Infection in Nervous Tissues of Slaughtered Pigs by Nested and Real-Time PCR. J. Microbiol. 2005, 43, 430–436. [Google Scholar] [PubMed]
- Cheng, T.Y.; Henao-Diaz, A.; Poonsuk, K.; Buckley, A.; van Geelen, A.; Lager, K.; Harmon, K.; Gauger, P.; Wang, C.; Ambagala, A.; et al. Pseudorabies (Aujeszky’s Disease) Virus DNA Detection in Swine Nasal Swab and Oral Fluid Specimens Using a GB-Based Real-Time Quantitative PCR. Prev. Vet. Med. 2021, 189, 105308. [Google Scholar] [CrossRef]
- Flatschart, R.B.; Maurício, R. Acute and Latent Infection in Mice with a Virulent Strain of Aujeszky’s Disease Virus. Braz. J. Microbiol. 2000, 31, 308–311. [Google Scholar] [CrossRef]
- Ren, C.Z.; Hu, W.Y.; Zhang, J.W.; Wei, Y.Y.; Yu, M.L.; Hu, T.J. Establishment of Inflammatory Model Induced by Pseudorabies Virus Infection in Mice. J. Vet. Sci. 2021, 22, e20. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, J.; Wu, Z.; Liu, J.; Ji, Q.; Ju, C. The Role of Latency-Associated Transcripts in the Latent Infection of Pseudorabies Virus. Viruses 2022, 14, 1379. https://doi.org/10.3390/v14071379
Deng J, Wu Z, Liu J, Ji Q, Ju C. The Role of Latency-Associated Transcripts in the Latent Infection of Pseudorabies Virus. Viruses. 2022; 14(7):1379. https://doi.org/10.3390/v14071379
Chicago/Turabian StyleDeng, Jiahuan, Zhuoyun Wu, Jiaqi Liu, Qiuyun Ji, and Chunmei Ju. 2022. "The Role of Latency-Associated Transcripts in the Latent Infection of Pseudorabies Virus" Viruses 14, no. 7: 1379. https://doi.org/10.3390/v14071379