
Antigenicity Alternations of Variant PEDV S Protein Disclosed by Linear B Cell Epitope Mapping

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
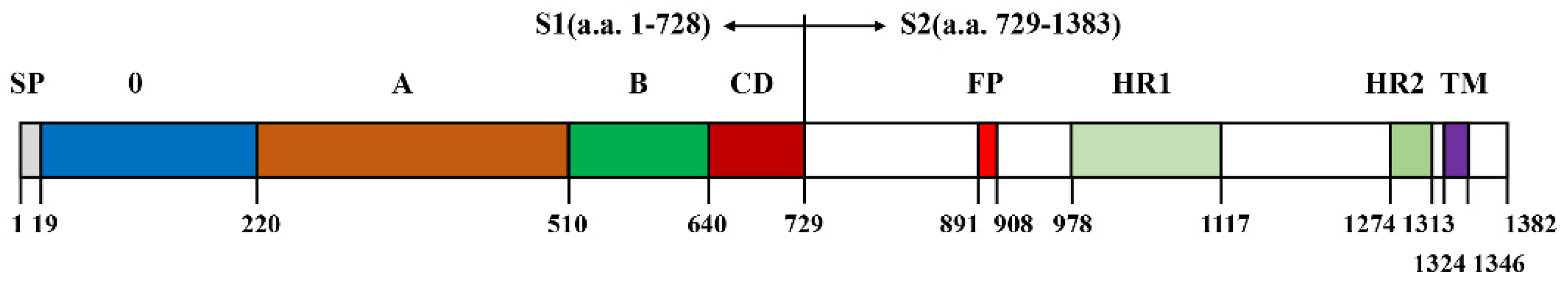
2.1. B Cell Epitope Prediction
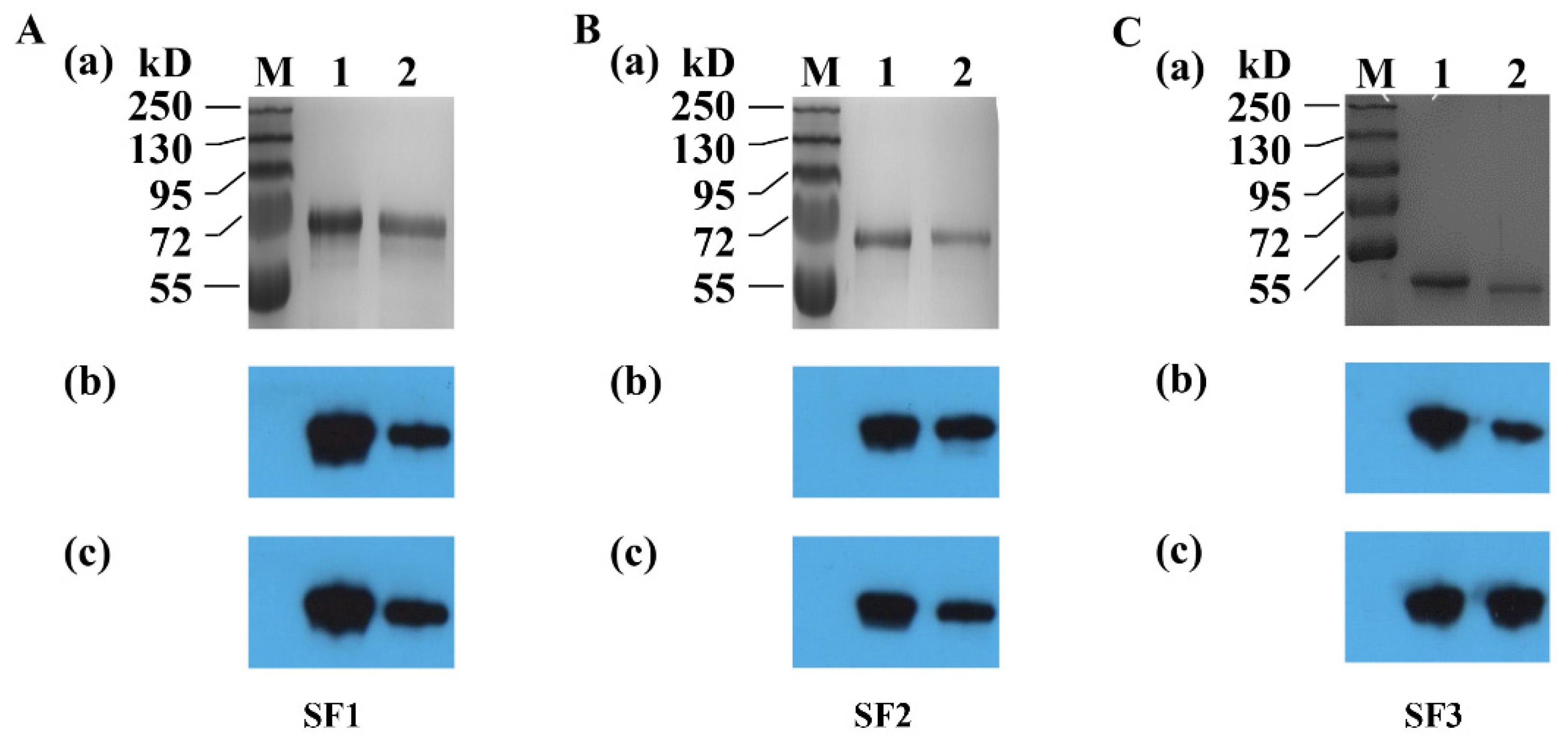
2.2. Preparation of Recombinant PEDV S Protein
2.3. Immunization of Animals
2.4. Biosynthesis of 16-Mer Peptides
2.5. SDS-PAGE and WB
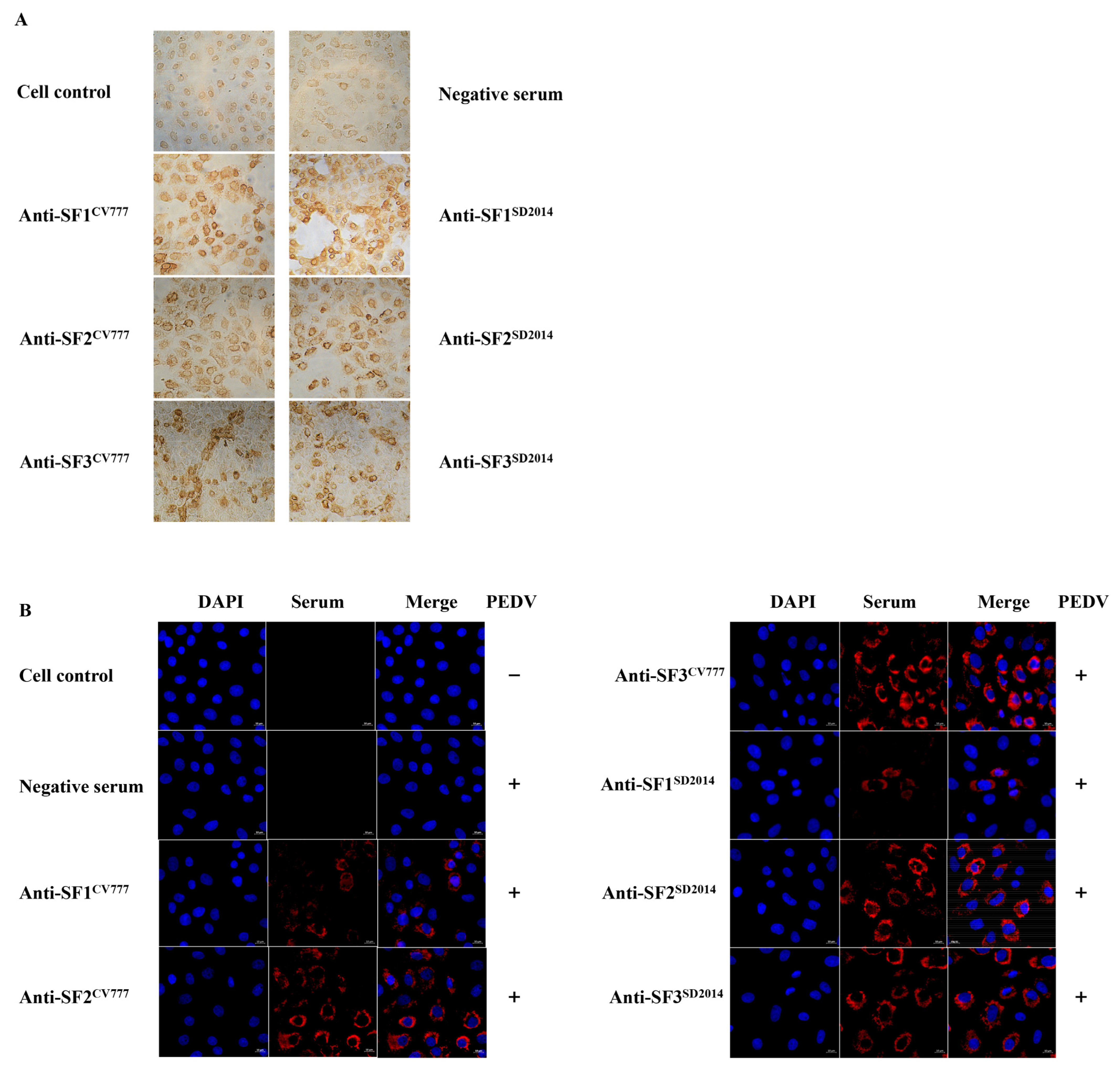
2.6. Immunofluorescence Assay (IFA)
2.7. Immunohistochemistry Assay (IHC)
3. Results
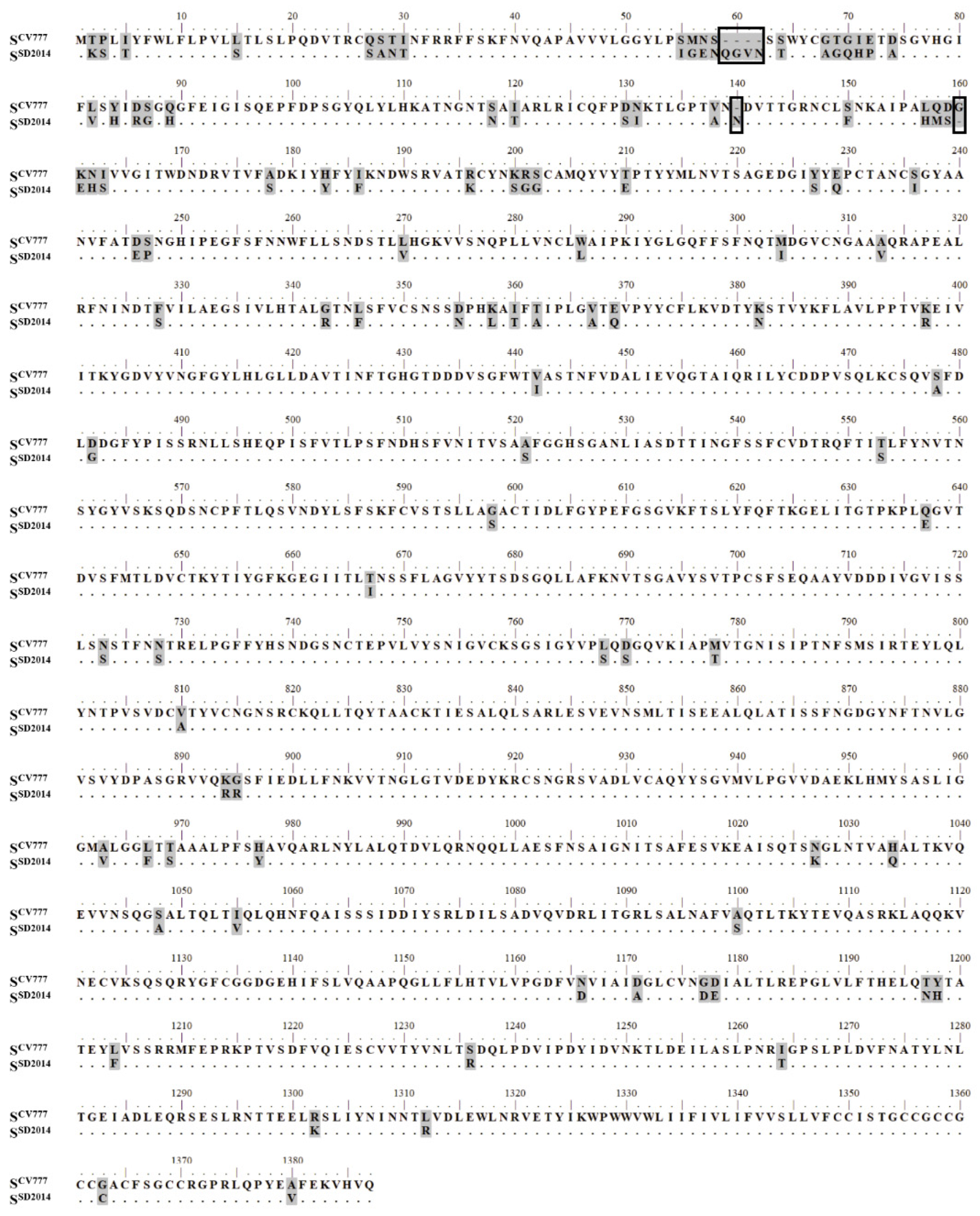
3.1. Prediction of BCEs on SCV777 and SSD2014
3.2. Reactivity of Acquired Anti-S Sera with PEDV DR13
3.3. Reactivity of Anti-S Sera with SCV777 and SSD2014
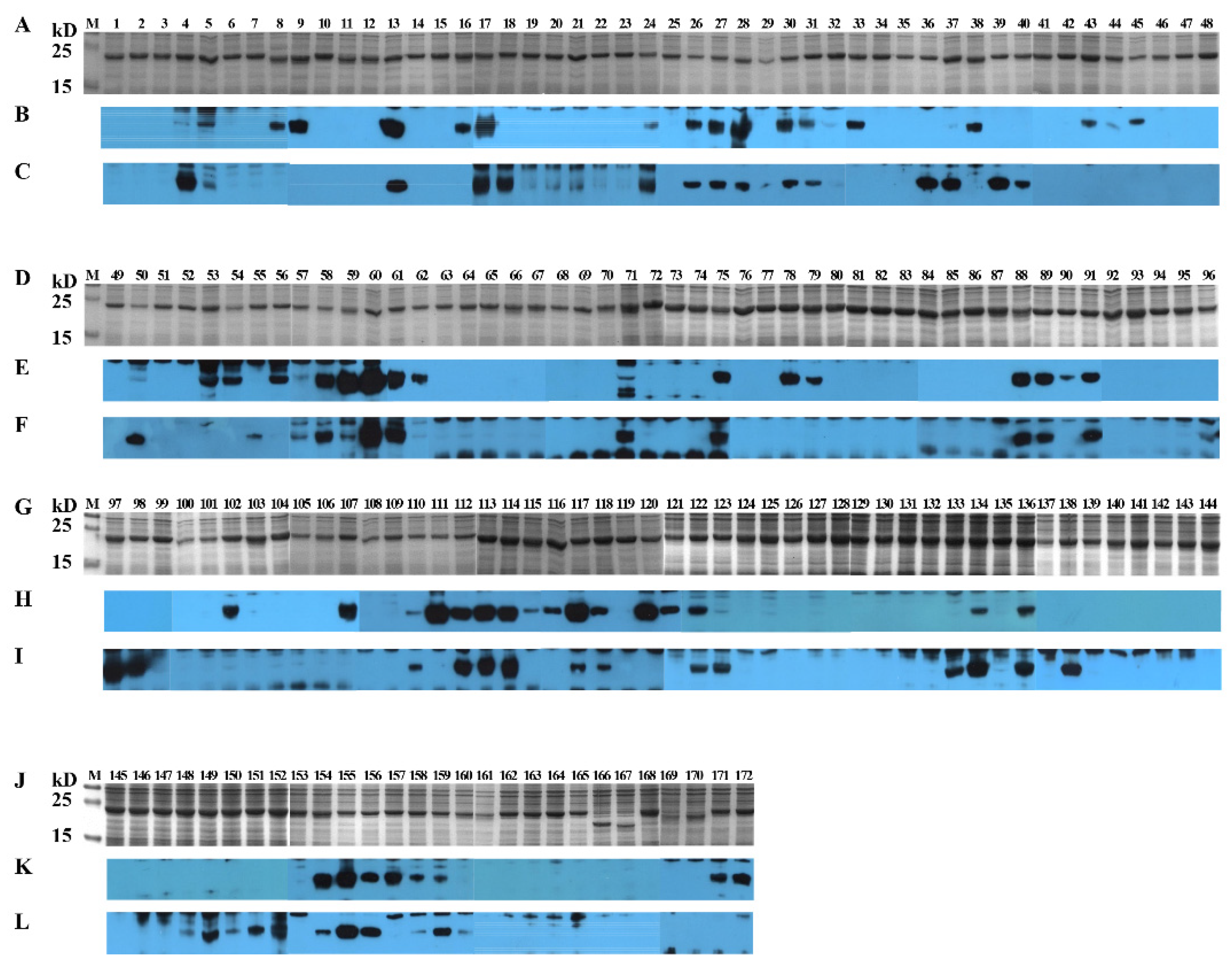
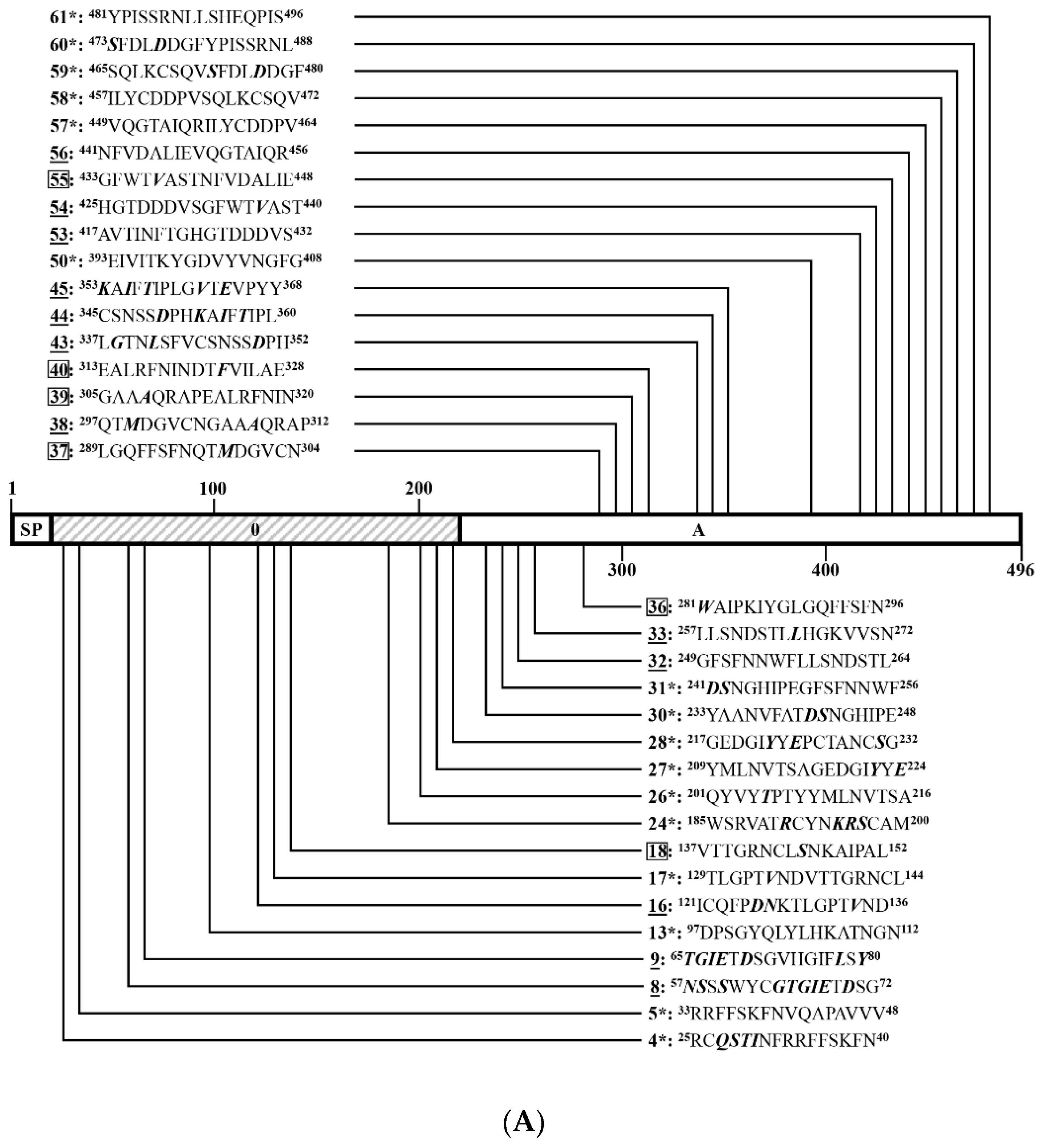
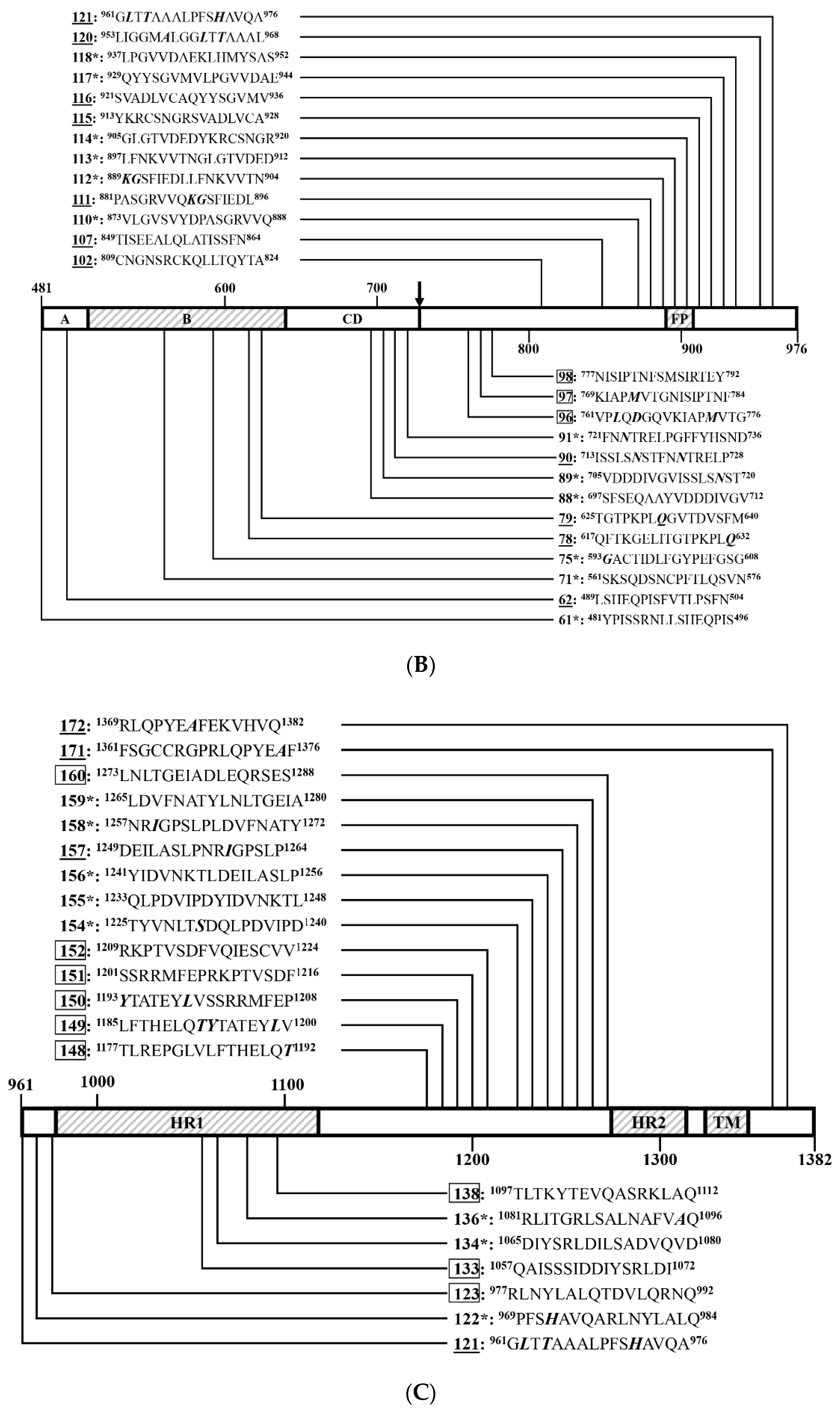
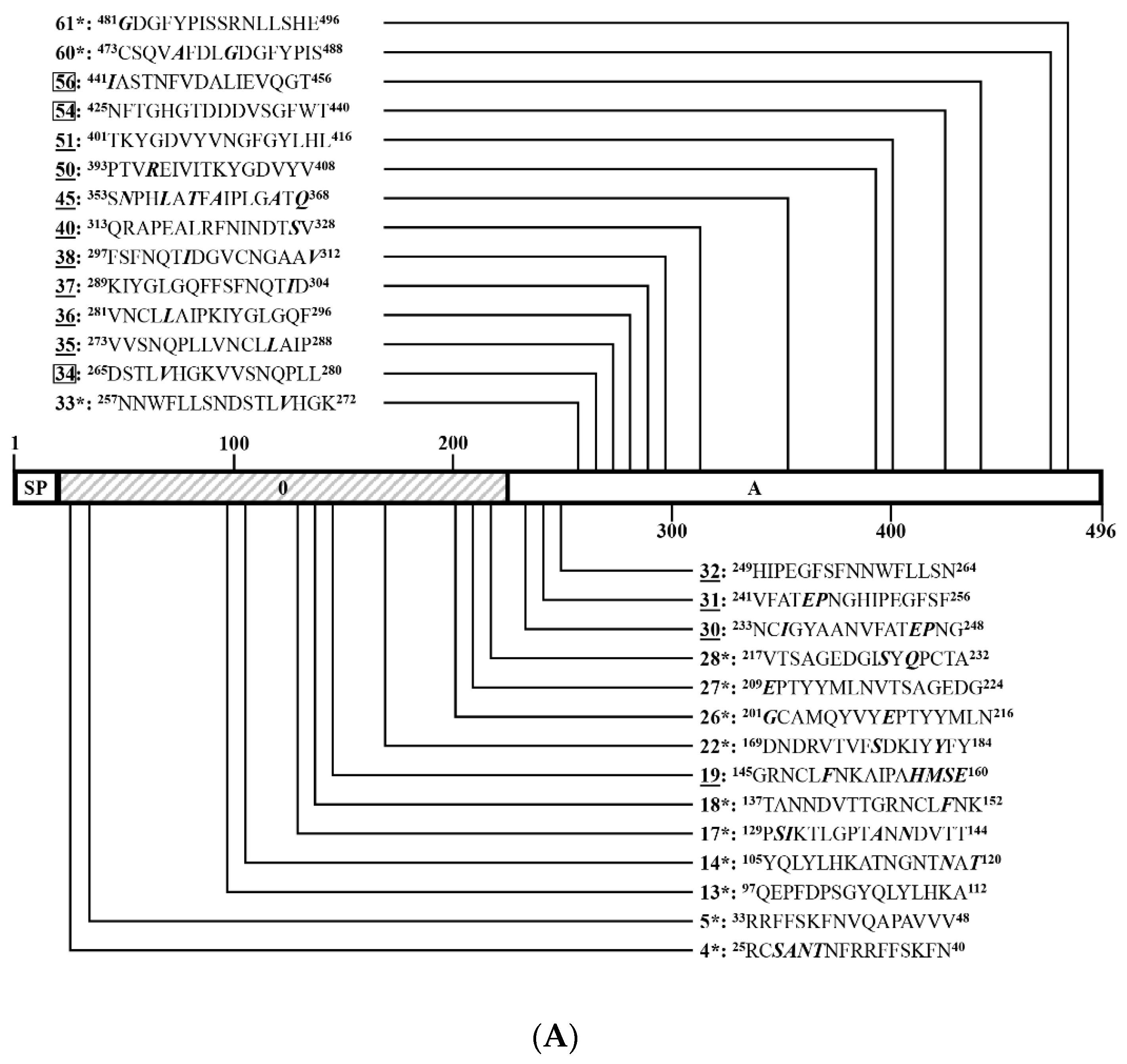
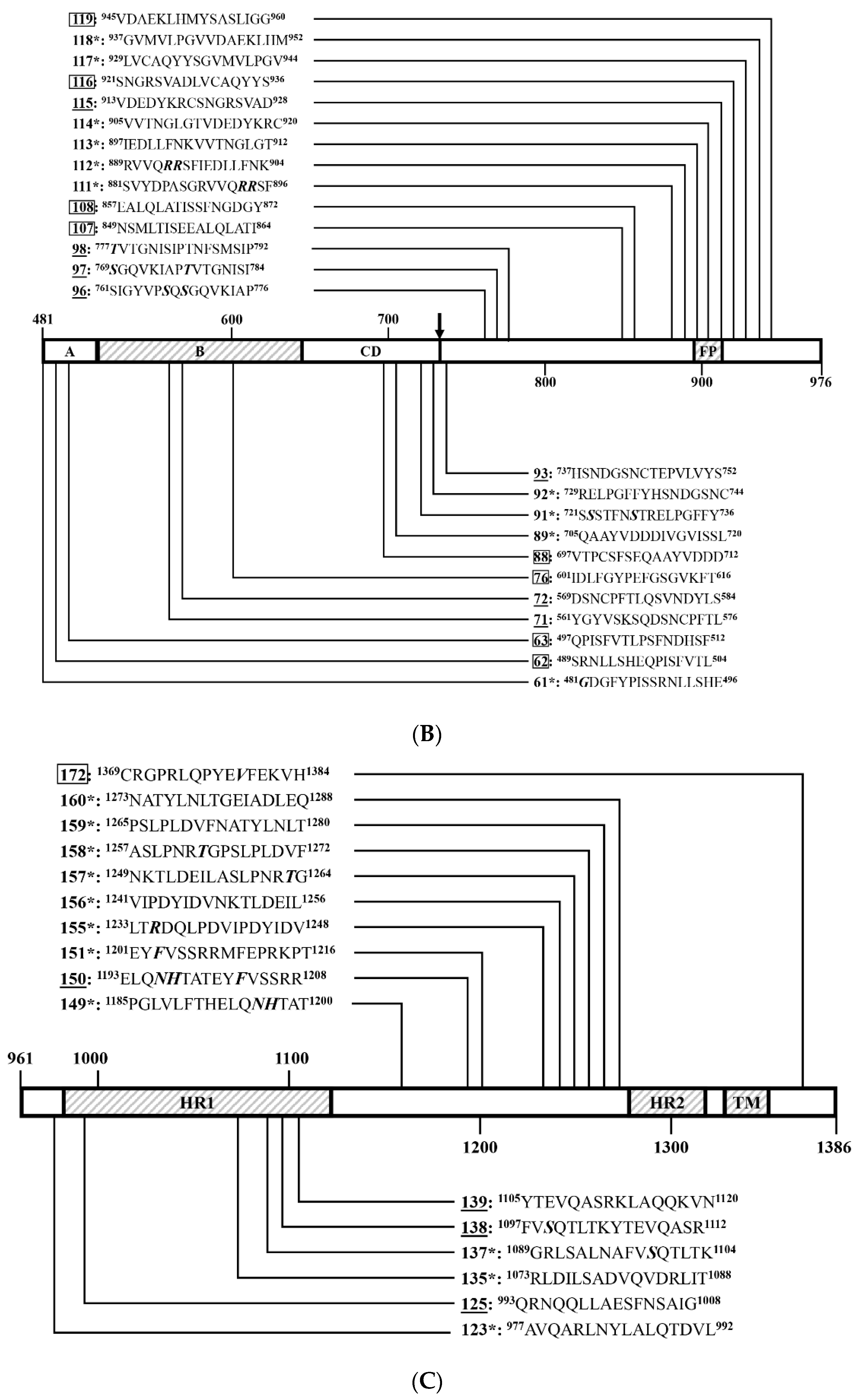
3.4. Identification of Positive 16-Mers on SCV777
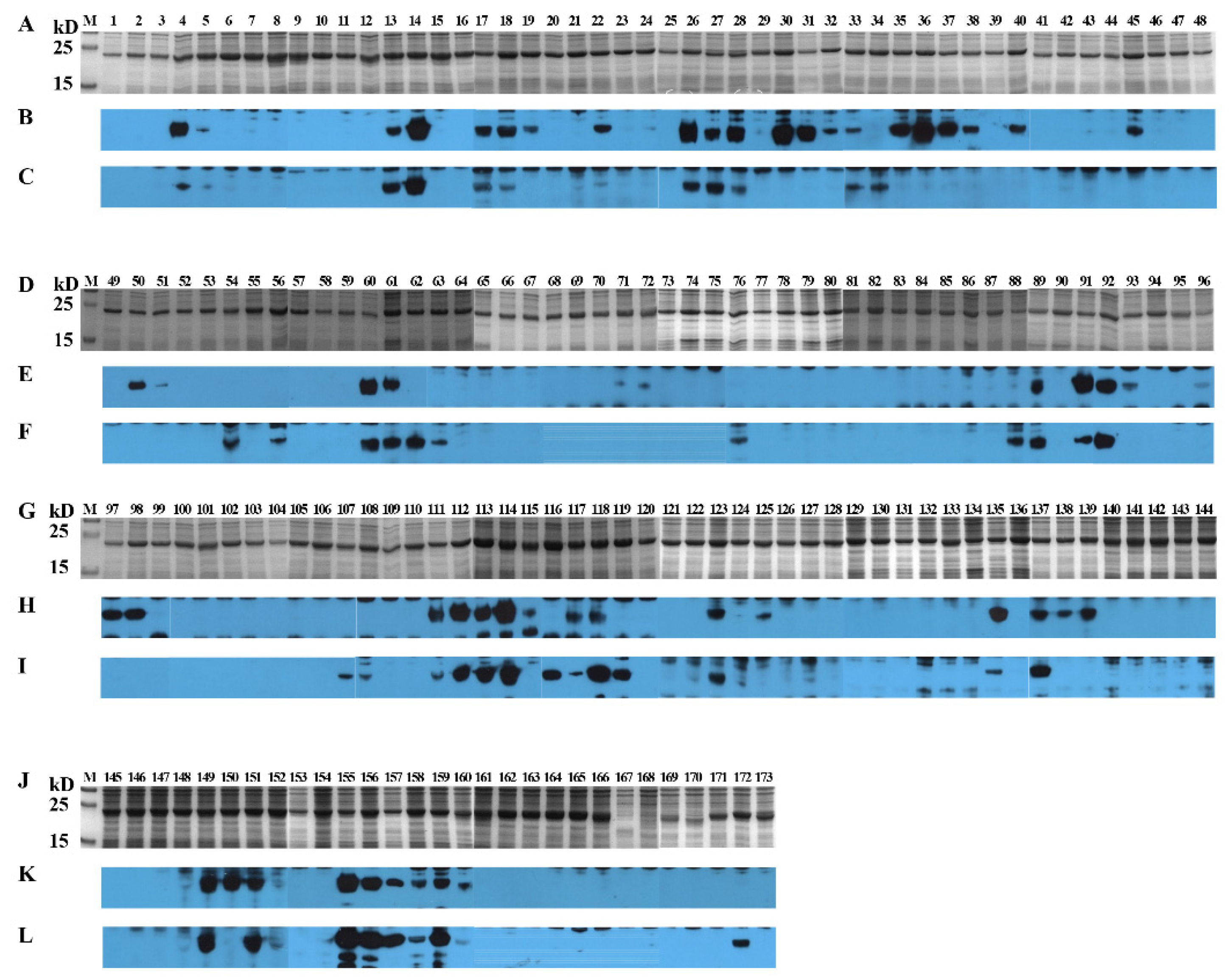
3.5. Identification of Positive 16-Mers on SSD2014
3.6. An Overview of Positive 16-Mers on SCV777 and SSD2014
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antas, M.; Wozniakowski, G. Current status of porcine epidemic diarrhoea (PED) in European pigs. J. Vet. Res. 2019, 63, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef]
- Lin, C.M.; Saif, L.J.; Marthaler, D.; Wang, Q. Evolution, antigenicity and pathogenicity of global porcine epidemic diarrhea virus strains. Virus Res. 2016, 226, 20–39. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of porcine epidemic diarrhea virus in the United States: Clinical signs, lesions, and viral genomic sequences. J. Vet. Diag. Invest 2013, 25, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Kubota, T.; Ujike, M.; Yahara, Y.; Taguchi, F. Phylogenetic and antigenic characterization of newly isolated porcine epidemic diarrhea viruses in Japan. Virus Res. 2016, 222, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y.; Liu, Y.; Chen, Y.; Jiao, W.; Feng, H.; Wei, Q.; Wang, J.; Zhang, Y.; Zhang, G. Isolation and identification of a recombinant porcine epidemic diarrhea virus with a novel insertion in S1 domain. Front. Microbiol. 2021, 12, 667084. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Bae, J.L.; Kang, T.J.; Kim, J.; Chung, G.H.; Lim, C.W.; Laude, H.; Yang, M.S.; Jang, Y.S. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cells 2002, 14, 295–299. [Google Scholar]
- Wang, D.; Fang, L.; Xiao, S. Porcine epidemic diarrhea in China. Virus Res. 2016, 226, 7–13. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Lucio de Esesarte, E.; Guo, H.; van den Elzen, P.; Aarts, E.; van den Born, E.; Rottier, P.J.M.; Bosch, B.J. Cell attachment domains of the porcine epidemic diarrhea virus spike protein are key targets of neutralizing antibodies. J. Virol. 2017, 91, e00273-17. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Cheng, I.C.; Chang, Y.C.; Tsai, P.S.; Lai, S.Y.; Huang, Y.L.; Jeng, C.R.; Pang, V.F.; Chang, H.W. Identification of neutralizing monoclonal antibodies targeting novel conformational epitopes of the porcine epidemic diarrhoea virus spike protein. Sci. Rep. 2019, 9, 2529. [Google Scholar] [CrossRef]
- Okda, F.A.; Lawson, S.; Singrey, A.; Nelson, J.; Hain, K.S.; Joshi, L.R.; Christopher-Hennings, J.; Nelson, E.A.; Diel, D.G. The S2 glycoprotein subunit of porcine epidemic diarrhea virus contains immunodominant neutralizing epitopes. Virology 2017, 509, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Li, F.; Shi, H.; Chen, J.; Cui, X.; Chen, H.; Liu, S.; Tong, Y.; Wang, Y.; Tong, G. Identification of two novel B cell epitopes on porcine epidemic diarrhea virus spike protein. Vet. Microbiol. 2008, 131, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Cruz, D.J.; Kim, C.J.; Shin, H.J. Phage-displayed peptides having antigenic similarities with porcine epidemic diarrhea virus (PEDV) neutralizing epitopes. Virology 2006, 354, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, D.J.M.; Kim, C.J.; Shin, H.J. The GPRLQPY motif located at the carboxy-terminal of the spike protein induces antibodies that neutralize Porcine epidemic diarrhea virus. Virus Res. 2008, 132, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, Z.; Zeng, X.; Zhang, G.; Niu, J.; Sun, B.; Ma, J. Sequence analysis of the spike gene of Porcine epidemic diarrhea virus isolated from South China during 2011–2015. J. Vet. Sci. 2017, 18, 237–243. [Google Scholar] [CrossRef]
- Zhang, F.; Luo, S.; Gu, J.; Li, Z.; Li, K.; Yuan, W.; Ye, Y.; Li, H.; Ding, Z.; Song, D.; et al. Prevalence and phylogenetic analysis of porcine diarrhea associated viruses in southern China from 2012 to 2018. BMC Vet. Res. 2019, 15, 470. [Google Scholar] [CrossRef] [Green Version]
- Than, V.T.; Choe, S.E.; Vu, T.T.H.; Do, T.D.; Nguyen, T.L.; Bui, T.T.N.; Mai, T.N.; Cha, R.M.; Song, D.; An, D.J.; et al. Genetic characterization of the spike gene of porcine epidemic diarrhea viruses (PEDVs) circulating in Vietnam from 2015 to 2016. Vet. Med. Sci. 2020, 6, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Lee, J.M.; Jung, J.; Kim, I.J.; Hyun, B.H.; Kim, H.I.; Park, C.K.; Oem, J.K.; Kim, Y.H.; Lee, M.H.; et al. Genetic characterization of porcine epidemic diarrhea virus in Korea from 1998 to 2013. Arch. Virol. 2015, 160, 1055–1064. [Google Scholar] [CrossRef]
- Yu, R.; Zhu, R.; Gao, W.; Zhang, M.; Dong, S.; Chen, B.; Yu, L.; Xie, C.; Jiang, F.; Li, Z. Fine mapping and conservation analysis of linear B-cell epitopes of peste des petits ruminants virus hemagglutinin protein. Vet. Microbiol. 2017, 208, 110–117. [Google Scholar] [CrossRef]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, Y.; Han, X.; Yu, Z.; Wei, Y.; Zhang, G. Porcine epidemic diarrhea virus in Asia: An alarming threat to the global pig industry. Infect. Genet. Evol. 2019, 70, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ma, Z.; Li, Y.; Gao, S.; Xiao, S. Porcine epidemic diarrhea virus: Molecular mechanisms of attenuation and vaccines. Microb. Pathog. 2020, 149, 104553. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.M.; Damschroder, M.M.; Lowe, D.C. Current approaches to fine mapping of antigen-antibody interactions. Immunology 2014, 142, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Cherryholmes, G.A.; Stanton, S.E.; Disis, M.L. Current methods of epitope identification for cancer vaccine design. Vaccine 2015, 33, 7408–7414. [Google Scholar] [CrossRef]
- Potocnakova, L.; Bhide, M.; Pulzova, L.B. An Introduction to B-Cell Epitope Mapping and In Silico Epitope Prediction. J. Immunol. Res. 2016, 2016, 6760830. [Google Scholar] [CrossRef] [Green Version]
- Polyiam, K.; Ruengjitchatchawalya, M.; Mekvichitsaeng, P.; Kaeoket, K.; Hoonsuwan, T.; Joiphaeng, P.; Roshorm, Y.M. Immunodominant and Neutralizing Linear B-Cell Epitopes Spanning the Spike and Membrane Proteins of Porcine Epidemic Diarrhea Virus. Front. Immunol. 2022, 12, 785293. [Google Scholar] [CrossRef]
- Opriessnig, T.; Gerber, P.F.; Shen, H.; de Castro, A.; Zhang, J.; Chen, Q.; Halbur, P. Evaluation of the efficacy of a commercial inactivated genogroup 2b-based porcine epidemic diarrhea virus (PEDV) vaccine and experimental live genogroup 1b exposure against 2b challenge. Vet. Res. 2017, 48, 69. [Google Scholar] [CrossRef]
- Sato, T.; Oroku, K.; Ohshima, Y.; Furuya, Y.; Sasakawa, C. Efficacy of genogroup 1 based porcine epidemic diarrhea live vaccine against genogroup 2 field strain in Japan. Virol. J. 2018, 15, 28. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Yuan, W.; Peng, Q.; Zhang, F.; Ye, Y.; Huang, D.; Ding, Z.; Lin, L.; He, H.; et al. Evaluation of Cross–Protection between G1a and G2a-Genotype Porcine Epidemic Diarrhea Viruses in Suckling Piglets. Animals 2020, 10, 1674. [Google Scholar] [CrossRef]
- Lin, C.M.; Gao, X.; Oka, T.; Vlasova, A.N.; Esseili, M.A.; Wang, Q.; Saif, L.J. Antigenic relationships among porcine epidemic diarrhea virus and transmissible gastroenteritis virus strains. J. Virol. 2015, 89, 3332–3342. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.G.; Li, R.; Xie, S.; Qiao, S.; Li, Q.; Chen, X.X.; Deng, R.; Zhang, G. Identification of a novel linear B-cell epitope within the collagenase equivalent domain of porcine epidemic diarrhea virus spike glycoprotein. Virus Res. 2019, 266, 34–42. [Google Scholar] [CrossRef]
- Liu, J.; Cong, G.; Shi, H.; Wang, Y.; Gao, R.; Chen, J.; Zhang, X.; Shi, D.; Cao, L.; Wang, X.; et al. Neutralizing monoclonal antibody fails to recognize porcine epidemic diarrhea virus with two regions deleted in spike protein. Vet. Microbiol. 2020, 251, 108917. [Google Scholar] [CrossRef]
- Gong, L.; Gong, Y.; Lin, Y.; Qin, J.; Liu, Y.; Qian, X.; Xue, C.; Cao, Y. Identification and characterization of a novel linear epitope in the spike protein of the porcine epidemic diarrhea virus. Acta Virol. 2019, 63, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Zhao, J.; Wang, X.; Zhao, Y.; Wang, C. Development of a hybridoma cell line secreting monoclonal antibody against S protein of a Chinese variant of PEDV. Monoclon. Antib. Immunodiagn. Immunother. 2015, 34, 12–16. [Google Scholar] [CrossRef]
- Li, W.; van Kuppeveld, F.J.M.; He, Q.; Rottier, P.J.M.; Bosch, B.J. Cellular entry of the porcine epidemic diarrhea virus. Virus Res. 2016, 226, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Huan, C.C.; Wang, Y.; Ni, B.; Wang, R.; Huang, L.; Ren, X.F.; Tong, G.Z.; Ding, C.; Fan, H.J.; Mao, X. Porcine epidemic diarrhea virus uses cell-surface heparan sulfate as an attachment factor. Arch. Virol. 2015, 160, 1621–1628. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Bhandari, M.; Martini, O.; Sewall, L.M.; Bangaru, S.; Yoon, K.J.; Ward, A.B. Structure and immune recognition of the porcine epidemic diarrhea virus spike protein. Structure 2021, 29, 385–392. [Google Scholar] [CrossRef]
- Kong, N.; Meng, Q.; Jiao, Y.; Wu, Y.; Zuo, Y.; Wang, H.; Sun, D.; Dong, S.; Zhai, H.; Tong, W.; et al. Identification of a novel B-cell epitope in the spike protein of porcine epidemic diarrhea virus. Virol. J. 2020, 17, 46. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Ge, X.; Gao, Y.; Zarlenga, D.S.; Wang, K.; Li, X.; Qin, Z.; Yin, X.; Liu, J.; Ren, X.; et al. Putative phage-display epitopes of the porcine epidemic diarrhea virus S1 protein and their anti-viral activity. Virus Genes 2015, 51, 217–224. [Google Scholar] [CrossRef]
- Burton, D.R. Antibodies, viruses and vaccines. Nat. Rev. Immunol. 2002, 2, 706–713. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCV777 | SSD2014 | ||||||
|---|---|---|---|---|---|---|---|
| No. | Position | Sequences | Antigenicity a | No. | Position | Sequences | Antigenicity |
| E1 | 19–30 | LPQDVTRCQSTIN | 0.0361 | E1 | 21–28 | QDVTRCSA | −0.0665 |
| E2 | 52–62 | AVVVLGGYLPSMNSSSWY | 0.1657 | E2 | 55–64 | IGENQGVNST | 0.3703 |
| E3 | 65–72 | TGIETDSG | 0.3072 | E3 | 70–76 | QHPTASG | 0.9738 |
| E4 | 91–100 | ISQEPFDPSG | 0.6991 | E4 | 95–105 | ISQEPFDPSGY | 0.6324 |
| E5 | 102–110 | QLYLHKATN | −0.3804 | ||||
| E5 | 111–120 | KATNGNTNAT | 0.4115 | ||||
| E6 | 125–142 | QLYLHKATNGNTSAIARLRICQFPDNKTLGPTVNDVTTGRN | 0.3121 | E6 | 133–146 | TLGPTANNDVTTGR | 1.0705 |
| E7 | 186–192 | SRVATRC | 2.1198 | E7 | 188–194 | DWSRVAT | 0.5834 |
| E8 | 195–202 | KRSCAMQY | 0.2537 | ||||
| E9 | 215–223 | SAGEDGIYY | −0.0055 | E8 | 219–233 | SAGEDGISYQPCTAN | 0.0702 |
| E10 | 226–232 | CTANCSG | −0.4497 | ||||
| E11 | 238–248 | FATDSNGHIPE | −0.7913 | E9 | 244–253 | TEPNGHIPEG | −0.7045 |
| E12 | 297–310 | QTMDGVCNGAAAQR | 0.4745 | E10 | 301–307 | QTIDGVC | 1.0282 |
| E13 | 347–353 | NSSDPHK | 0.2269 | E11 | 350–356 | SNSSNPH | 0.4187 |
| E14 | 372–378 | KVDTYKS | −1.422 | ||||
| E15 | 423–432 | TGHGTDDDVS | 0.5537 | E12 | 427–436 | TGHGTDDDVS | 0.5537 |
| E16 | 461–475 | DDPVSQLKCSQVSFD | 0.8676 | E13 | 465–473 | DDPVSQLKC | 0.4517 |
| E17 | 483–490 | ISSRNLLS | 0.4671 | ||||
| E14 | 520–526 | SFGGHSG | 0.4737 | ||||
| E18 | 528–535 | SDTTINGF | −0.3571 | E15 | 530–539 | IASDTTINGF | −0.4022 |
| E19 | 555–573 | NSYGYVSKSQDSNCPFTLQ | 0.8752 | E16 | 559–575 | NSYGYVSKSQDSNCPFT | 0.7085 |
| E17 | 608–613 | EFGSGV | 0.6906 | ||||
| E20 | 622–634 | ELITGTPKPLQGV | 0.1569 | E18 | 627–640 | LITGTPKPLEGVTD | 0.2387 |
| E21 | 688–694 | GAVYSVT | 0.0153 | ||||
| E22 | 716–728 | LSNSTFNNTRELP | 0.4482 | E19 | 720–732 | LSSSTFNSTRELP | 0.2311 |
| E23 | 734–744 | SNDGSNCTEPV | 0.3639 | E20 | 738–747 | SNDGSNCTEP | 0.4006 |
| E21 | 762–775 | IGYVPSQSGQVKIA | 0.8513 | ||||
| E24 | 800–817 | VSVDCVTYVCNGNSRCKQ | 0.8176 | E22 | 803–819 | PVSVDCATYVCNGNSRC | 0.5376 |
| E25 | 860–869 | ISSFNGDGYN | −0.77 | E23 | 865–873 | SSFNGDGYN | −0.7927 |
| E26 | 875–887 | GVSVYDPASGRVV | 0.5215 | E24 | 879–893 | GVSVYDPASGRVVQR | 0.6336 |
| E27 | 904–921 | NGLGTVDEDYKRCSNGRS | 0.2071 | E25 | 906–925 | VTNGLGTVDEDYKRCSNGRS | 0.2239 |
| E28 | 1013–1024 | VKEAISQTSNGL | 0.0891 | E26 | 1017–1030 | VKEAISQTSKGLNT | 0.3561 |
| E29 | 1037–1043 | VVNSQGS | −0.0001 | ||||
| E27 | 1064–1070 | SSSIDDI | 0.4527 | ||||
| E30 | 1118–1131 | CVKSQSQRYGFCGG | 0.6518 | E28 | 1101–1135 | TLTKYTEVQASRKLAQQKVNECVKSQSQRYGFCGG | 0.2232 |
| E31 | 1205–1214 | MFEPRKPTVS | 0.4124 | E29 | 1194–1218 | LQNHTATEYFVSSRRMFEPRKPTVS | 0.0433 |
| E32 | 1230–1243 | TSDQLPDVIPDYID | 0.4996 | E30 | 1233–1246 | LTRDQLPDVIPDYI | 0.3167 |
| E33 | 1256–1262 | PNRIGPS | 1.1446 | E31 | 1259–1267 | LPNRTGPSL | 0.9542 |
| E34 | 1280–1297 | ADLEQRSESLRNTTEELR | 0.615 | E32 | 1284–1300 | ADLEQRSESLRNTTEEL | 0.5571 |
| E35 | 1348–1373 | TGCCGCCGCCGACFSGCCRGPRLQPY | −0.2869 | E33 | 1351–1361 | STGCCGCCGCC | −0.5985 |
| E34 | 1364–1376 | CFSGCCRGPRLQP | 0.1851 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, R.; Dong, S.; Chen, B.; Liu, Y.; Li, F.; Si, F.; Xie, C.; Li, Z. Antigenicity Alternations of Variant PEDV S Protein Disclosed by Linear B Cell Epitope Mapping. Viruses 2022, 14, 1371. https://doi.org/10.3390/v14071371
Yu R, Dong S, Chen B, Liu Y, Li F, Si F, Xie C, Li Z. Antigenicity Alternations of Variant PEDV S Protein Disclosed by Linear B Cell Epitope Mapping. Viruses. 2022; 14(7):1371. https://doi.org/10.3390/v14071371
Chicago/Turabian StyleYu, Ruisong, Shijuan Dong, Bingqing Chen, Yingjie Liu, Fengping Li, Fusheng Si, Chunfang Xie, and Zhen Li. 2022. "Antigenicity Alternations of Variant PEDV S Protein Disclosed by Linear B Cell Epitope Mapping" Viruses 14, no. 7: 1371. https://doi.org/10.3390/v14071371