
Fusion Proteins CLD and CLDmut Demonstrate Potent and Broad Neutralizing Activity against HIV-1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. HIV-1 Production
2.3. Design and Genetic Engineering of Protein Expression Constructs
2.4. Expression and Purification of CLD and CLDmut Proteins
2.5. SDS-PAGE, Native PAGE and Western Blot
2.6. HIV-1 Neutralization Assay
2.7. Bio-Layer Interferometry (BLI)
2.8. Cytotoxicity Assay
2.9. Statistical Analysis
3. Results
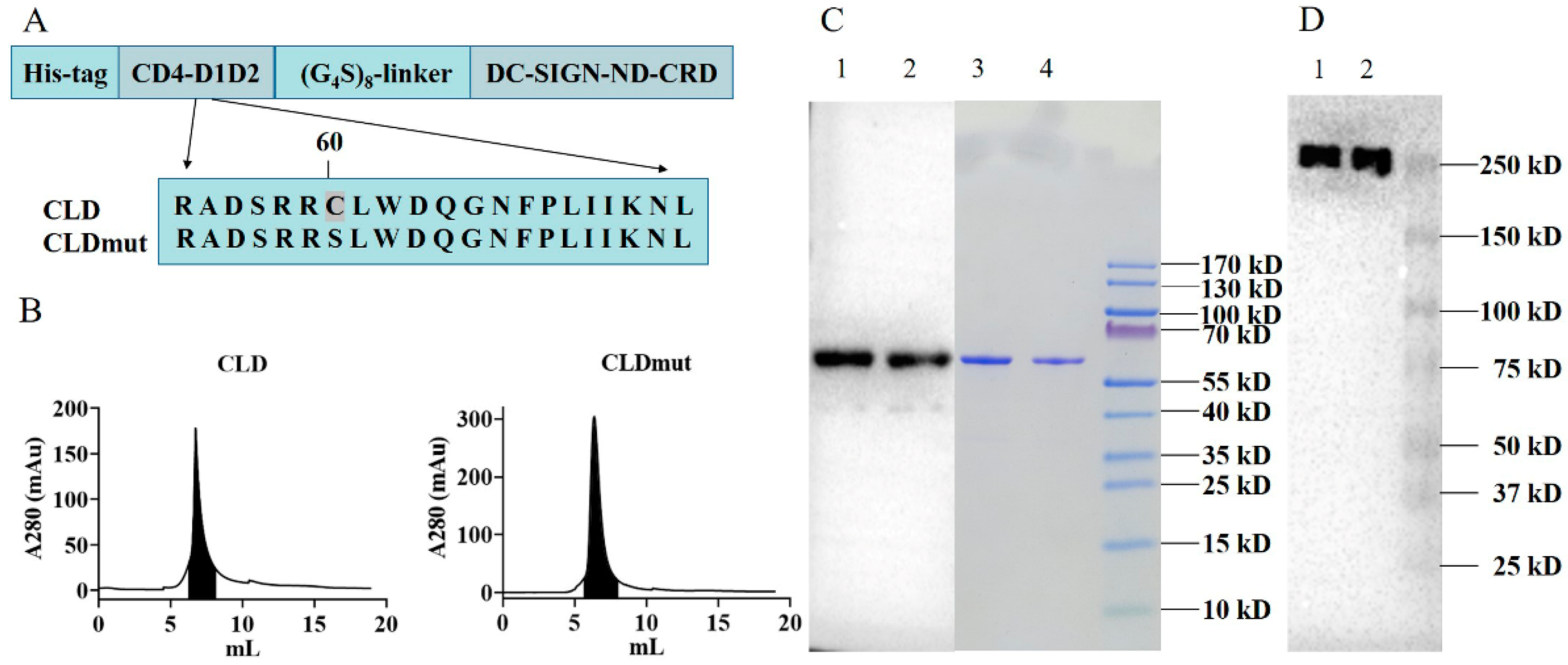
3.1. Production and Purification of CLD and CLDmut Proteins in 293F Cells
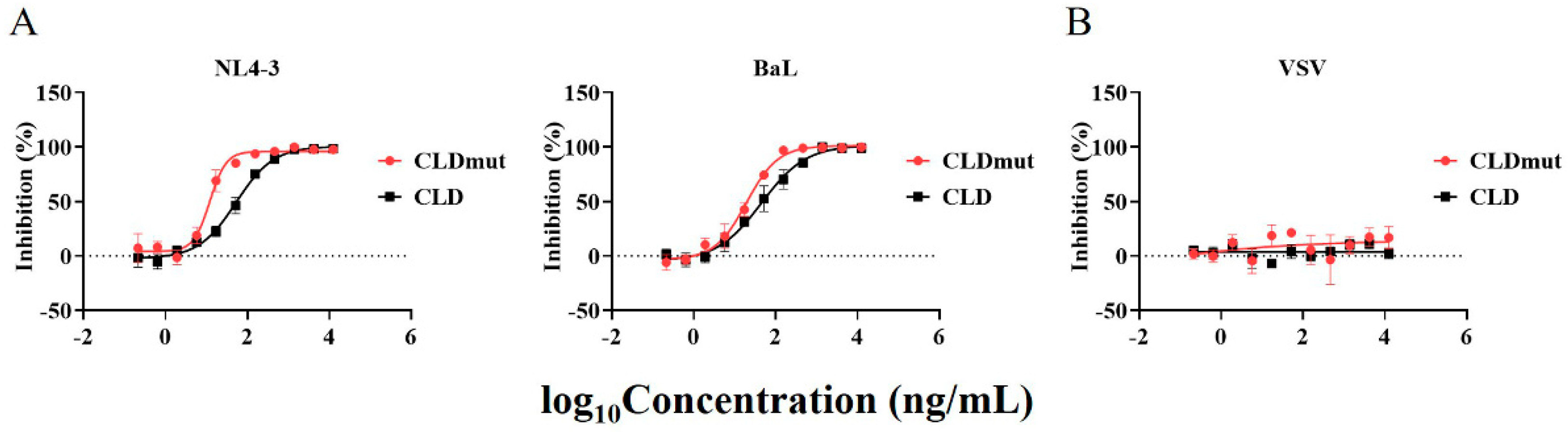
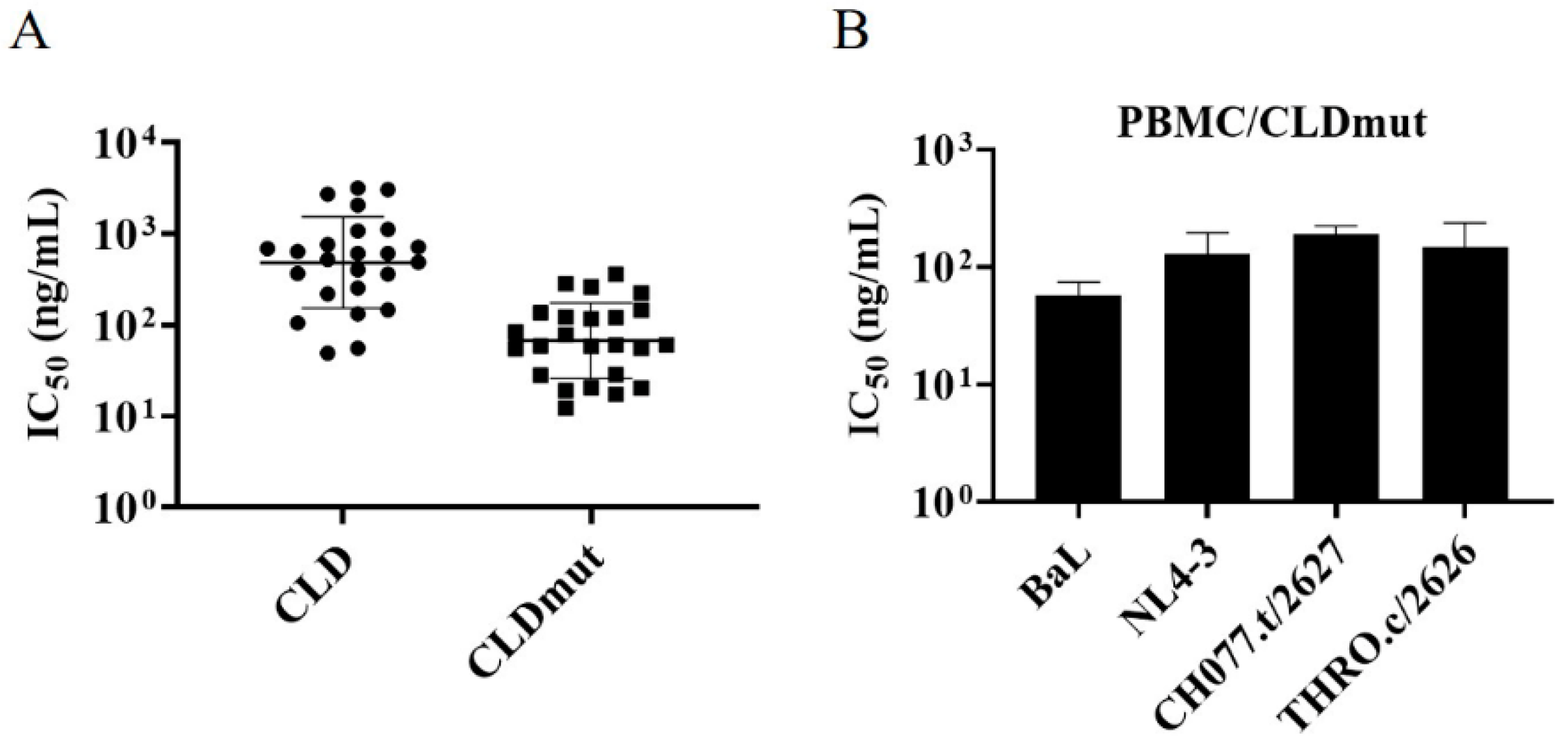
3.2. CLD and CLDmut Proteins Block the Infection of a Wide Range of HIV-1 Isolates
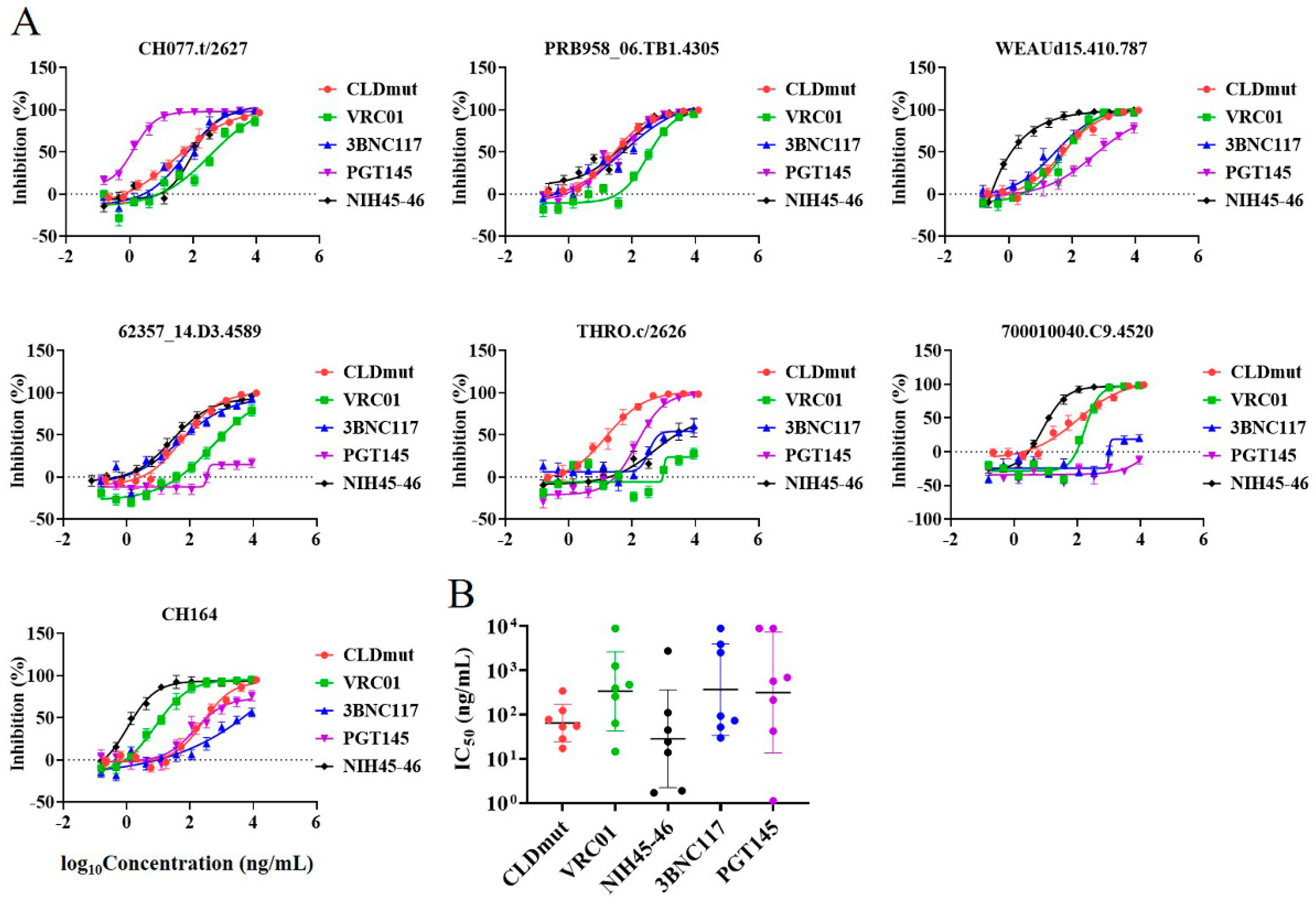
3.3. CLDmut Has More Potent or Comparable Neutralizing Activity against HIV-1 in Comparison with bNAbs
3.4. CLDmut Has Strong Binding Affinity to HIV-1 gp140
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heiken, H. Antiretroviral treatment for HIV: Is it time for 2 drug regimens? MMW Fortschr. Med. 2019, 161, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.; Ho, D.D.; Abdool Karim, Q. HIV/AIDS epidemiology, pathogenesis, prevention, and treatment. Lancet 2006, 368, 489–504. [Google Scholar] [CrossRef] [Green Version]
- Montessori, V.; Press, N.; Harris, M.; Akagi, L.; Montaner, J.S. Adverse effects of antiretroviral therapy for HIV infection. Can. Med. Assoc. J. 2004, 170, 229–238. [Google Scholar]
- Nolan, D.; Reiss, P.; Mallal, S. Adverse effects of antiretroviral therapy for HIV infection: A review of selected topics. Expert Opin. Drug Saf. 2005, 4, 201–218. [Google Scholar] [CrossRef]
- Rodriguez Cumplido, D.; Asensio Ostos, C. Biological and biosimilar drugs: Clarifying concepts. Aten. Prim. 2018, 50, 323–324. [Google Scholar] [CrossRef]
- Kos, J. Biological drugs. Farm. Vestn. 2009, 60, 61–66. [Google Scholar]
- Fernández-Cruz, E.; Alecsandru, D.; Rodríguez-Sainz, C. Introduction to biological drugs. Actas Dermosifiliogr. 2008, 99 (Suppl. S4), 2–6. [Google Scholar]
- Lalezari, J.P.; Luber, A.D. Enfuvirtide. Drugs Today 2004, 40, 259–269. [Google Scholar] [CrossRef]
- Joly, V.; Jidar, K.; Tatay, M.; Yeni, P. Enfuvirtide: From basic investigations to current clinical use. Expert Opin. Pharmacother. 2010, 11, 2701–2713. [Google Scholar] [CrossRef]
- Huang, J.; Ofek, G.; Laub, L.; Louder, M.K.; Doria-Rose, N.A.; Longo, N.S.; Imamichi, H.; Bailer, R.T.; Chakrabarti, B.; Sharma, S.K.; et al. Broad and potent neutralization of HIV-1 by a gp41-specific human antibody. Nature 2012, 491, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caskey, M.; Klein, F.; Lorenzi, J.C.; Seaman, M.S.; West, A.P., Jr.; Buckley, N.; Kremer, G.; Nogueira, L.; Braunschweig, M.; Scheid, J.F.; et al. Viraemia suppressed in HIV-1-infected humans by broadly neutralizing antibody 3BNC117. Nature 2015, 522, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalgleish, A.G.; Beverley, P.C.; Clapham, P.R.; Crawford, D.H.; Greaves, M.F.; Weiss, R.A. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature 1984, 312, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, Y.; Gong, R.; Zhu, Z.; Wang, Y.; Zhao, Q.; Dimitrov, D.S. Engineered single human CD4 domains as potent HIV-1 inhibitors and components of vaccine immunogens. J. Virol. 2011, 85, 9395–9405. [Google Scholar] [CrossRef] [Green Version]
- Traunecker, A.; Lüke, W.; Karjalainen, K. Soluble CD4 molecules neutralize human immunodeficiency virus type 1. Nature 1988, 331, 84–86. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Feinberg, H.; Guo, Y.; Mitchell, D.A.; Drickamer, K.; Weis, W.I. Extended neck regions stabilize tetramers of the receptors DC-SIGN and DC-SIGNR. J. Biol. Chem. 2005, 280, 1327–1335. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Hu, K.; Yang, J.; Jin, J.; Li, C.; Stieh, D.; Griffin, G.E.; Shattock, R.J.; Hu, Q. Bifunctional CD4-DC-SIGN fusion proteins demonstrate enhanced avidity to gp120 and inhibit HIV-1 infection and dissemination. Antimicrob. Agents Chemother. 2012, 56, 4640–4649. [Google Scholar] [CrossRef] [Green Version]
- Hoorelbeke, B.; Van Montfort, T.; Xue, J.; LiWang, P.J.; Tanaka, H.; Igarashi, Y.; van Damme, E.J.; Sanders, R.W.; Balzarini, J. HIV-1 envelope trimer has similar binding characteristics for carbohydrate-binding agents as monomeric gp120. FEBS Lett. 2013, 587, 860–866. [Google Scholar] [CrossRef] [Green Version]
- Peakman, M.; Senaldi, G.; Foote, N.; McManus, T.J.; Vergani, D. Naturally occurring soluble CD4 in patients with human immunodeficiency virus infection. J. Infect. Dis. 1992, 165, 799–804. [Google Scholar] [CrossRef]
- Plazolles, N.; Humbert, J.-M.; Vachot, L.; Verrier, B.; Hocke, C.; Halary, F. Pivotal advance: The promotion of soluble DC-SIGN release by inflammatory signals and its enhancement of cytomegalovirus-mediated cis-infection of myeloid dendritic cells. J. Leukoc. Biol. 2011, 89, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Ceballos-Alcantarilla, E.; Merkx, M. Understanding and applications of Ser/Gly linkers in protein engineering. Methods Enzymol. 2021, 647, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Rinas, U.; Garcia-Fruitos, E.; Corchero, J.L.; Vazquez, E.; Seras-Franzoso, J.; Villaverde, A. Bacterial Inclusion Bodies: Discovering Their Better Half. Trends Biochem. Sci. 2017, 42, 726–737. [Google Scholar] [CrossRef]
- Cerutti, N.; Mendelow, B.V.; Napier, G.B.; Papathanasopoulos, M.A.; Killick, M.; Khati, M.; Stevens, W.; Capovilla, A. Stabilization of HIV-1 gp120-CD4 receptor complex through targeted interchain disulfide exchange. J. Biol. Chem. 2010, 285, 25743–25752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Napier, K.B.; Trent, J.O.; Wang, Z.; Taylor, S.; Griffin, G.E.; Peiper, S.C.; Shattock, R.J. Restricted variable residues in the C-terminal segment of HIV-1 V3 loop regulate the molecular anatomy of CCR5 utilization. J. Mol. Biol. 2005, 350, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.X.; Barry, A.P.; Wang, Z.X.; Connolly, S.M.; Peiper, S.C.; Greenberg, M.L. Evolution of the human immunodeficiency virus type 1 envelope during infection reveals molecular corollaries of specificity for coreceptor utilization and AIDS pathogenesis. J. Virol. 2000, 74, 11858–11872. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Jin, W.; Hu, K.; Luo, S.; Du, T.; Griffin, G.E.; Shattock, R.J.; Hu, Q. Highly conserved HIV-1 gp120 glycans proximal to CD4-binding region affect viral infectivity and neutralizing antibody induction. Virology 2012, 423, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.; Hu, K.; Hu, H.; Ni, F.; Du, T.; Shattock, R.J.; Hu, Q. Antigenicity and immunogenicity of HIV-1 gp140 with different combinations of glycan mutation and V1/V2 region or V3 crown deletion. Vaccine 2019, 37, 7501–7508. [Google Scholar] [CrossRef]
- Ingale, J.; Wyatt, R.T. Kinetic Analysis of Monoclonal Antibody Binding to HIV-1 gp120-derived Hyperglycosylated Cores. Bio-Protoc. 2015, 5, e1615. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Du, T.; Li, C.; Luo, S.; Liu, Y.; Huang, X.; Hu, Q. Sensitivity of transmitted and founder human immunodeficiency virus type 1 envelopes to carbohydrate-binding agents griffithsin, cyanovirin-N and Galanthus nivalis agglutinin. J. Gen. Virol. 2015, 96, 3660–3666. [Google Scholar] [CrossRef]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatell, J.M. Antiretroviral therapy for HIV: Do subtypes matter? Clin. Infect. Dis. 2011, 53, 1153–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geretti, A.M.; Harrison, L.; Green, H.; Sabin, C.; Hill, T.; Fearnhill, E.; Pillay, D.; Dunn, D. Effect of HIV-1 subtype on virologic and immunologic response to starting highly active antiretroviral therapy. Clin. Infect. Dis. 2009, 48, 1296–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemelaar, J. The origin and diversity of the HIV-1 pandemic. Trends Mol. Med. 2012, 18, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Kantor, R. Impact of HIV-1 pol diversity on drug resistance and its clinical implications. Curr. Opin. Infect. Dis. 2006, 19, 594–606. [Google Scholar] [CrossRef]
- Khattar, S.K.; Samal, S.; LaBranche, C.C.; Montefiori, D.C.; Collins, P.L.; Samal, S.K. Comparative immunogenicity of HIV-1 gp160, gp140 and gp120 expressed by live attenuated newcastle disease virus vector. PLoS ONE 2013, 8, e78521. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.J.; Bai, J.; Liu, F.L.; Zhang, X.Y.; Wang, J.Z. Focus on the therapeutic efficacy of 3BNC117 against HIV-1: In vitro studies, in vivo studies, clinical trials and challenges. Int. Immunopharmacol. 2017, 52, 44–50. [Google Scholar] [CrossRef]
- Roy, C.N.; Khandaker, I.; Oshitani, H. Evolutionary Dynamics of Tat in HIV-1 Subtypes B and C. PLoS ONE 2015, 10, e0129896. [Google Scholar] [CrossRef]
- Lynch, R.M.; Boritz, E.; Coates, E.E.; DeZure, A.; Madden, P.; Costner, P.; Enama, M.E.; Plummer, S.; Holman, L.; Hendel, C.S.; et al. Virologic effects of broadly neutralizing antibody VRC01 administration during chronic HIV-1 infection. Sci. Transl. Med. 2015, 7, 319ra206. [Google Scholar] [CrossRef] [Green Version]
- Bar-On, Y.; Grüll, H.; Schoofs, T.; Pai, J.A.; Nogueira, L.; Butler, A.L.; Millard, K.; Lehmann, C.; Suárez, I.; Oliveira, T.; et al. Safety and antiviral activity of combination HIV-1 broadly neutralizing antibodies in viremic individuals. Nat. Med. 2018, 24, 1701–1707. [Google Scholar] [CrossRef]
- Mendoza, P.; Grüll, H.; Nogueira, L.; Pai, J.A.; Butler, A.L.; Millard, K.; Lehmann, C.; Suárez, I.; Oliveira, T.; Lorenzi, J.C.C.; et al. Combination therapy with anti-HIV-1 antibodies maintains viral suppression. Nature 2018, 561, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Caskey, M.; Schoofs, T.; Grüll, H.; Settler, A.; Karagounis, T.; Kreider, E.F.; Murrell, B.; Pfeifer, N.; Nogueira, L.; Oliveira, T.; et al. Antibody 10-1074 suppresses viremia in HIV-1-infected individuals. Nat. Med. 2017, 23, 185–191. [Google Scholar] [CrossRef]
- Stephenson, K.E.; Julg, B.; Tan, C.S.; Zash, R.; Walsh, S.R.; Rolle, C.P.; Monczor, A.N.; Lupo, S.; Gelderblom, H.C.; Ansel, J.L.; et al. Safety, pharmacokinetics and antiviral activity of PGT121, a broadly neutralizing monoclonal antibody against HIV-1: A randomized, placebo-controlled, phase 1 clinical trial. Nat. Med. 2021, 27, 1718–1724. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.R.; Farzan, M. Engineering antibody-like inhibitors to prevent and treat HIV-1 infection. Curr. Opin. HIV AIDS 2017, 12, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Gilbert, P.B.; Juraska, M.; Montefiori, D.C.; Morris, L.; Karuna, S.T.; Edupuganti, S.; Mgodi, N.M.; Decamp, A.C.; Rudnicki, E.; et al. Two Randomized Trials of Neutralizing Antibodies to Prevent HIV-1 Acquisition. N. Engl. J. Med. 2021, 384, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Kolchinsky, P.; Mirzabekov, T.; Farzan, M.; Kiprilov, E.; Cayabyab, M.; Mooney, L.J.; Choe, H.; Sodroski, J. Adaptation of a CCR5-using, primary human immunodeficiency virus type 1 isolate for CD4-independent replication. J. Virol. 1999, 73, 8120–8126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, T.L.; LaBranche, C.C.; Zhang, W.; Canziani, G.; Robinson, J.; Chaiken, I.; Hoxie, J.A.; Doms, R.W. Stable exposure of the coreceptor-binding site in a CD4-independent HIV-1 envelope protein. Proc. Natl. Acad. Sci. USA 1999, 96, 6359–6364. [Google Scholar] [CrossRef] [Green Version]
- Dumonceaux, J.; Nisole, S.; Chanel, C.; Quivet, L.; Amara, A.; Baleux, F.; Briand, P.; Hazan, U. Spontaneous mutations in the env gene of the human immunodeficiency virus type 1 NDK isolate are associated with a CD4-independent entry phenotype. J. Virol. 1998, 72, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.M.; Foulke, J.S.; Flinko, R.; Heredia, A.; DeVico, A.; Reitz, M. An alteration of human immunodeficiency virus gp41 leads to reduced CCR5 dependence and CD4 independence. J. Virol. 2008, 82, 5460–5471. [Google Scholar] [CrossRef] [Green Version]
- Xiao, P.; Usami, O.; Suzuki, Y.; Ling, H.; Shimizu, N.; Hoshino, H.; Zhuang, M.; Ashino, Y.; Gu, H.; Hattori, T. Characterization of a CD4-independent clinical HIV-1 that can efficiently infect human hepatocytes through chemokine (C-X-C motif) receptor 4. AIDS 2008, 22, 1749–1757. [Google Scholar] [CrossRef]
- Zerhouni, B.; Nelson, J.A.; Saha, K. Isolation of CD4-independent primary human immunodeficiency virus type 1 isolates that are syncytium inducing and acutely cytopathic for CD8+ lymphocytes. J. Virol. 2004, 78, 1243–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haim, H.; Strack, B.; Kassa, A.; Madani, N.; Wang, L.; Courter, J.R.; Princiotto, A.; McGee, K.; Pacheco, B.; Seaman, M.S.; et al. Contribution of intrinsic reactivity of the HIV-1 envelope glycoproteins to CD4-independent infection and global inhibitor sensitivity. PLoS Pathog. 2011, 7, e1002101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaway, G.P.; Davis-Bruno, K.L.; Beaudry, G.A.; Garcia, E.B.; Wong, E.L.; Ryder, A.M.; Hasel, K.W.; Gauduin, M.-C.; Koup, R.A.; McDougal, J.S.; et al. Expression and characterization of CD4-IgG2, a novel heterotetramer that neutralizes primary HIV type 1 isolates. AIDS Res. Hum. Retrovir. 1995, 11, 533–539. [Google Scholar] [CrossRef] [PubMed]
- West, A.P., Jr.; Galimidi, R.P.; Gnanapragasam, P.N.; Bjorkman, P.J. Single-chain Fv-based anti-HIV proteins: Potential and limitations. J. Virol. 2012, 86, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthos, J.; Cicala, C.; Steenbeke, T.D.; Chun, T.W.; Dela Cruz, C.D.; Hanback, D.B.; Khazanie, P.; Nam, D.; Schuck, P.; Selig, S.M.; et al. Biochemical and biological characterization of a dodecameric CD4-Ig fusion protein: Implications for therapeutic and vaccine strategies. J. Biol. Chem. 2002, 277, 11456–11464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastie, E.; Samulski, R.J. Adeno-associated virus at 50: A golden anniversary of discovery, research, and gene therapy success-a personal perspective. Hum Gene Ther. 2015, 26, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, S.P.; Martinez-Navio, J.M.; Piatak, M., Jr.; Lifson, J.D.; Gao, G.; Desrosiers, R.C. AAV-Delivered Antibody Mediates Significant Protective Effects against SIVmac239 Challenge in the Absence of Neutralizing Activity. PLoS Pathog. 2015, 11, e1005090. [Google Scholar] [CrossRef]
- Nieto, K.; Salvetti, A. AAV Vectors Vaccines Against Infectious Diseases. Front. Immunol. 2014, 5, 5. [Google Scholar] [CrossRef]
- DeVico, A.; Fouts, T.; Lewis, G.K.; Gallo, R.C.; Godfrey, K.; Charurat, M.; Harris, I.; Galmin, L.; Pal, R. Antibodies to CD4-induced sites in HIV gp120 correlate with the control of SHIV challenge in macaques vaccinated with subunit immunogens. Proc. Natl. Acad. Sci. USA 2007, 104, 17477–17482. [Google Scholar] [CrossRef] [Green Version]
- Fouts, T.; Godfrey, K.; Bobb, K.; Montefiori, D.; Hanson, C.V.; Kalyanaraman, V.S.; DeVico, A.; Pal, R. Crosslinked HIV-1 envelope-CD4 receptor complexes elicit broadly cross-reactive neutralizing antibodies in rhesus macaques. Proc. Natl. Acad. Sci. USA 2002, 99, 11842–11847. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, E.; Mingozzi, F.; Montane, J.; Leon, X.; Anguela, X.; Haurigot, V.A.; A Edmonson, S.; Africa, L.; Zhou, S.; High, K.; et al. High AAV vector purity results in serotype- and tissue-independent enhancement of transduction efficiency. Gene Ther. 2010, 17, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Veillette, M.; Coutu, M.; Richard, J.; Batraville, L.A.; Dagher, O.; Bernard, N.; Tremblay, C.; Kaufmann, D.E.; Roger, M.; Finzi, A. The HIV-1 gp120 CD4-bound conformation is preferentially targeted by antibody-dependent cellular cytotoxicity-mediating antibodies in sera from HIV-1-infected individuals. J. Virol. 2015, 89, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Hijazi, K.; Wang, Y.; Scala, C.; Jeffs, S.; Longstaff, C.; Stieh, D.; Haggarty, B.; Vanham, G.; Schols, D.; Balzarini, J.; et al. DC-SIGN increases the affinity of HIV-1 envelope glycoprotein interaction with CD4. PLoS ONE 2011, 6, e28307. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, M.; Xiao, Y.; Du, T.; Hu, H.; Ni, F.; Hu, K.; Hu, Q. Fusion Proteins CLD and CLDmut Demonstrate Potent and Broad Neutralizing Activity against HIV-1. Viruses 2022, 14, 1365. https://doi.org/10.3390/v14071365
Fu M, Xiao Y, Du T, Hu H, Ni F, Hu K, Hu Q. Fusion Proteins CLD and CLDmut Demonstrate Potent and Broad Neutralizing Activity against HIV-1. Viruses. 2022; 14(7):1365. https://doi.org/10.3390/v14071365
Chicago/Turabian StyleFu, Ming, Yingying Xiao, Tao Du, Huimin Hu, Fengfeng Ni, Kai Hu, and Qinxue Hu. 2022. "Fusion Proteins CLD and CLDmut Demonstrate Potent and Broad Neutralizing Activity against HIV-1" Viruses 14, no. 7: 1365. https://doi.org/10.3390/v14071365