
Tyrosine 146 of the Human Na+/Taurocholate Cotransporting Polypeptide (NTCP) Is Essential for Its Hepatitis B Virus (HBV) Receptor Function and HBV Entry into Hepatocytes

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Side-Directed Mutagenesis and Cell Transfection
2.4. Generation of NTCP-HEK293 Stable Cell Lines
2.5. Western Blotting
2.6. Bile Acid Transport Assay
2.7. Myr-PreS1 Peptide Binding Assay
2.8. HBV Infection of HepG2-NTCP Cells
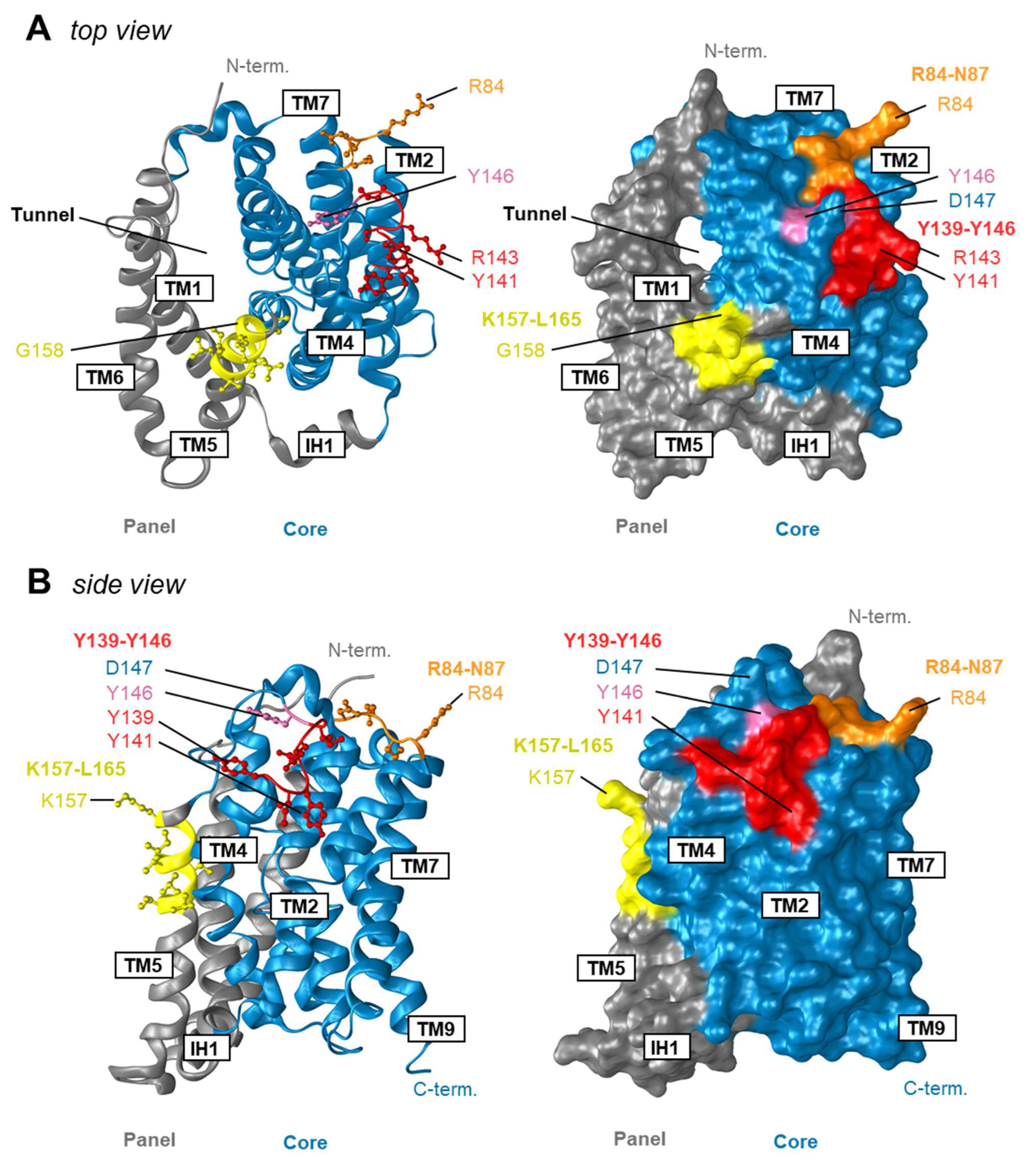
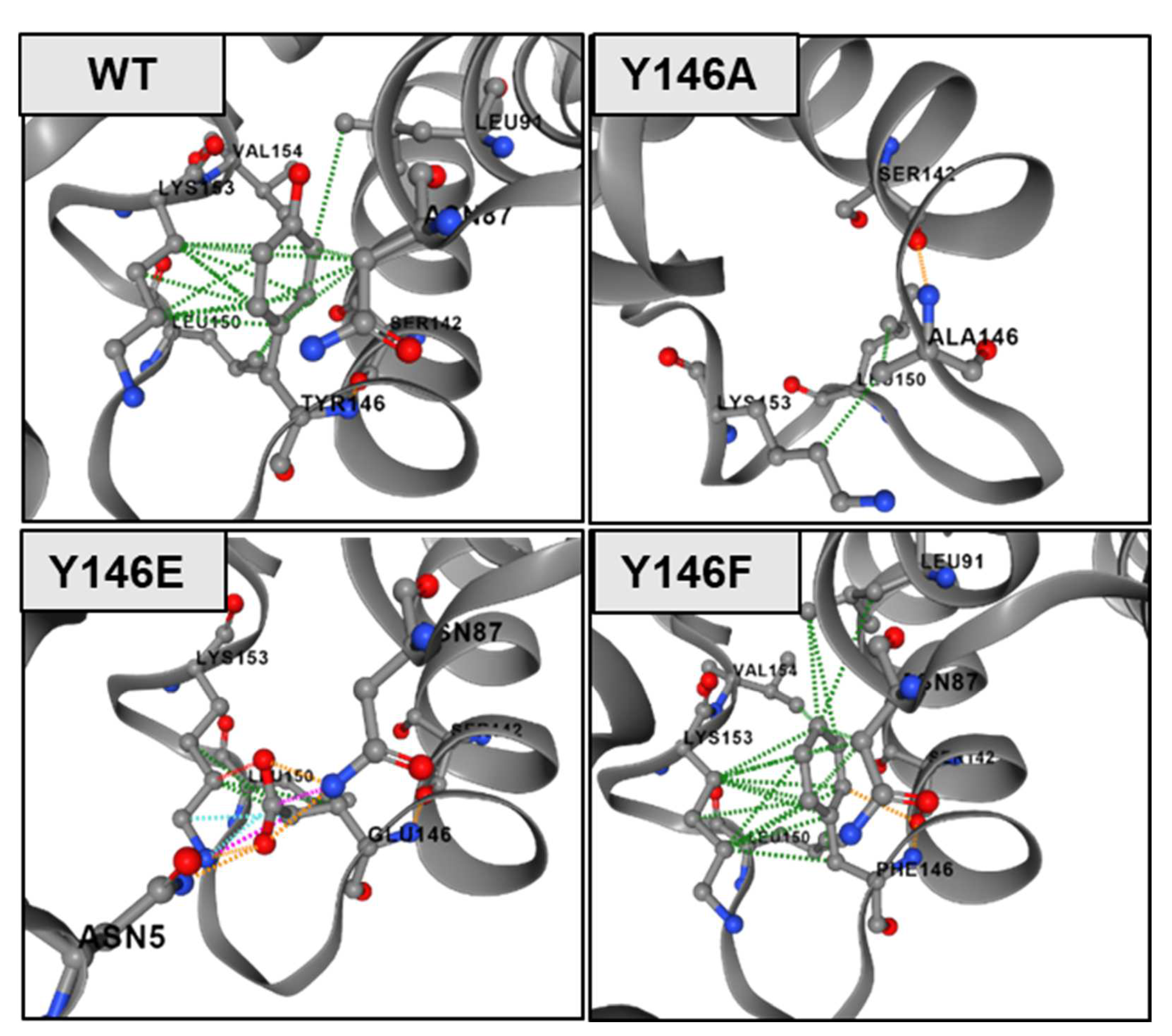
2.9. Modelling of Human NTCP
2.10. Statistics
3. Results
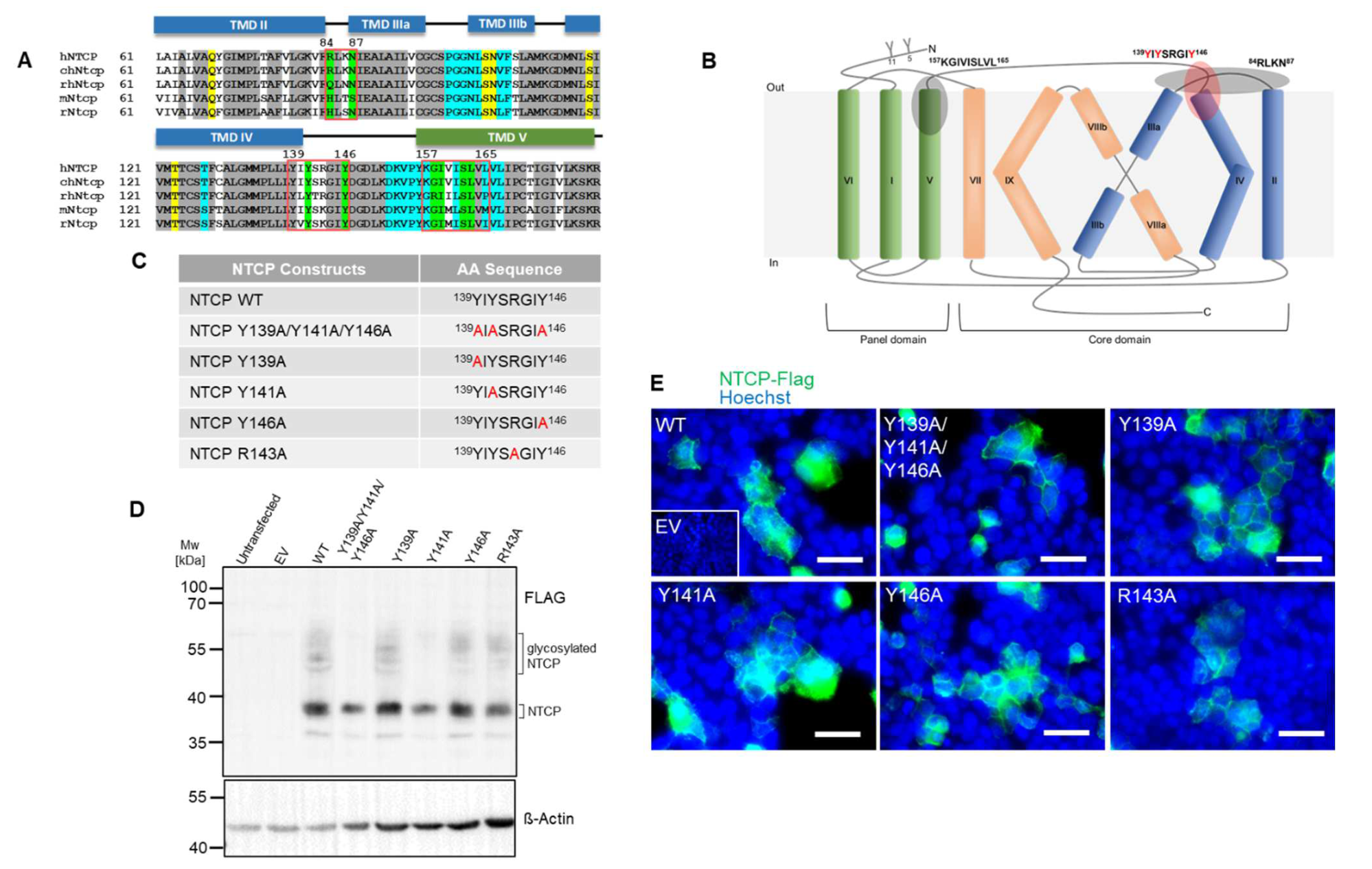
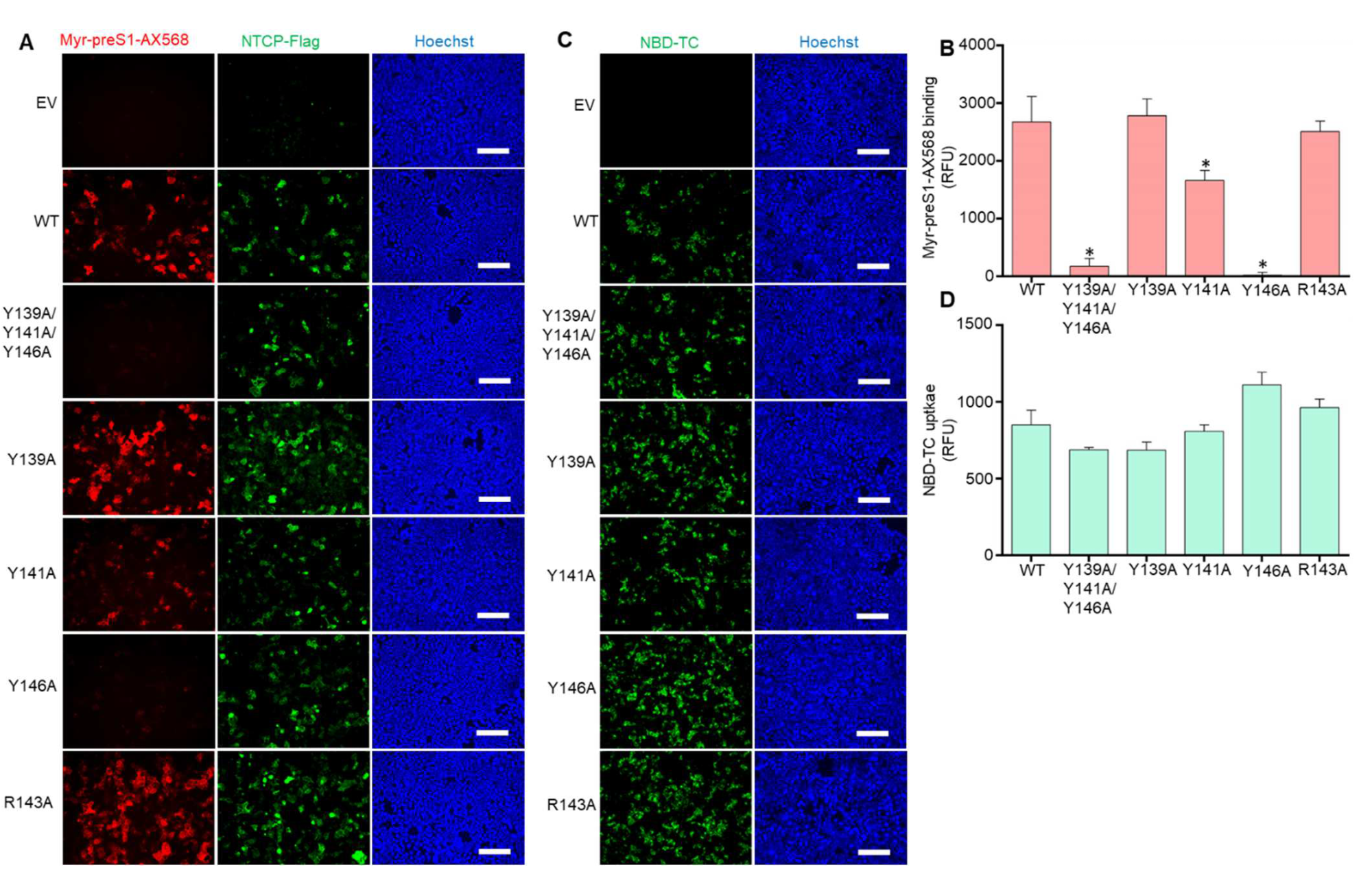
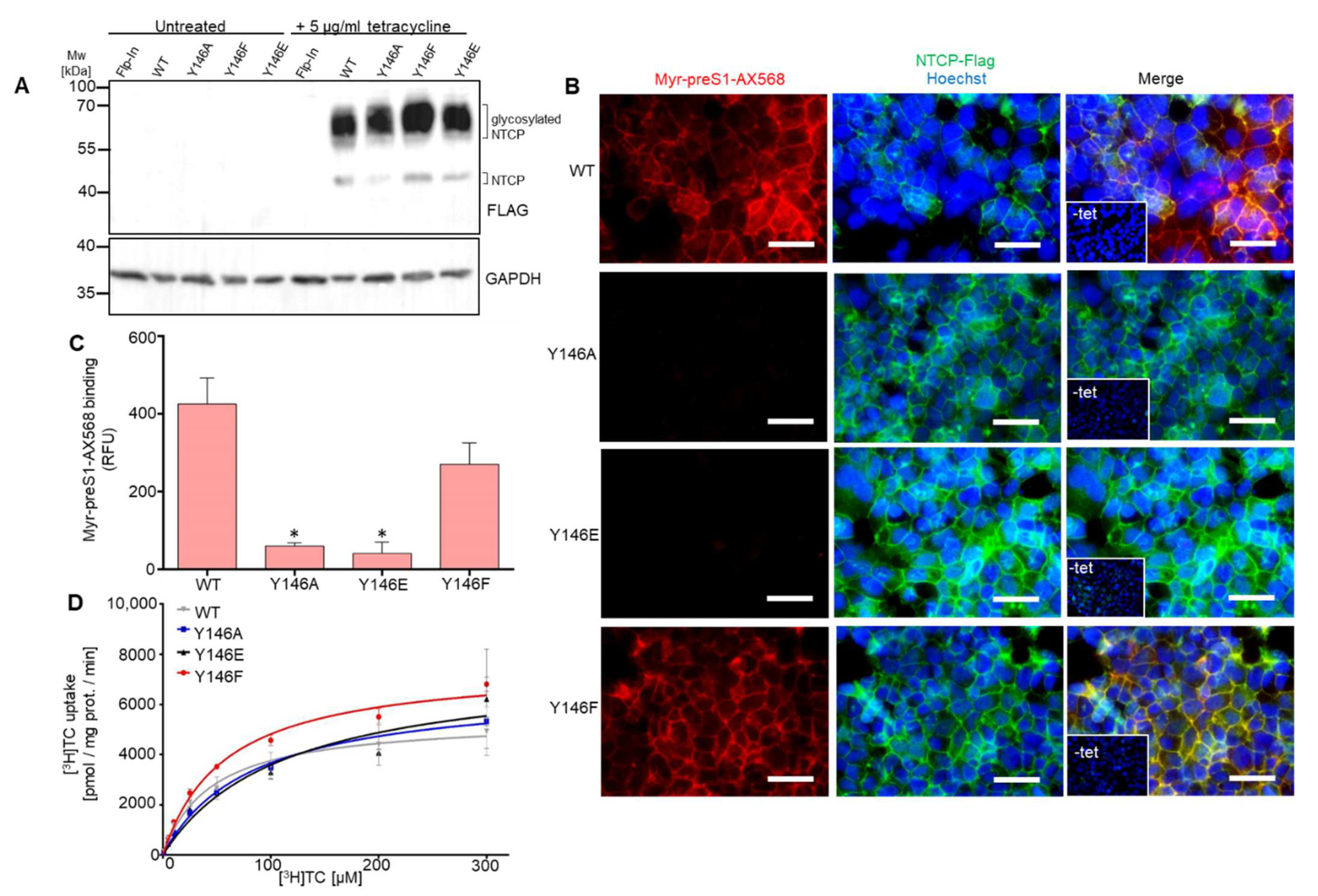
3.1. The NTCP 139YIYSRGIY146 Motif Is Crucial for PreS1-Peptide Binding
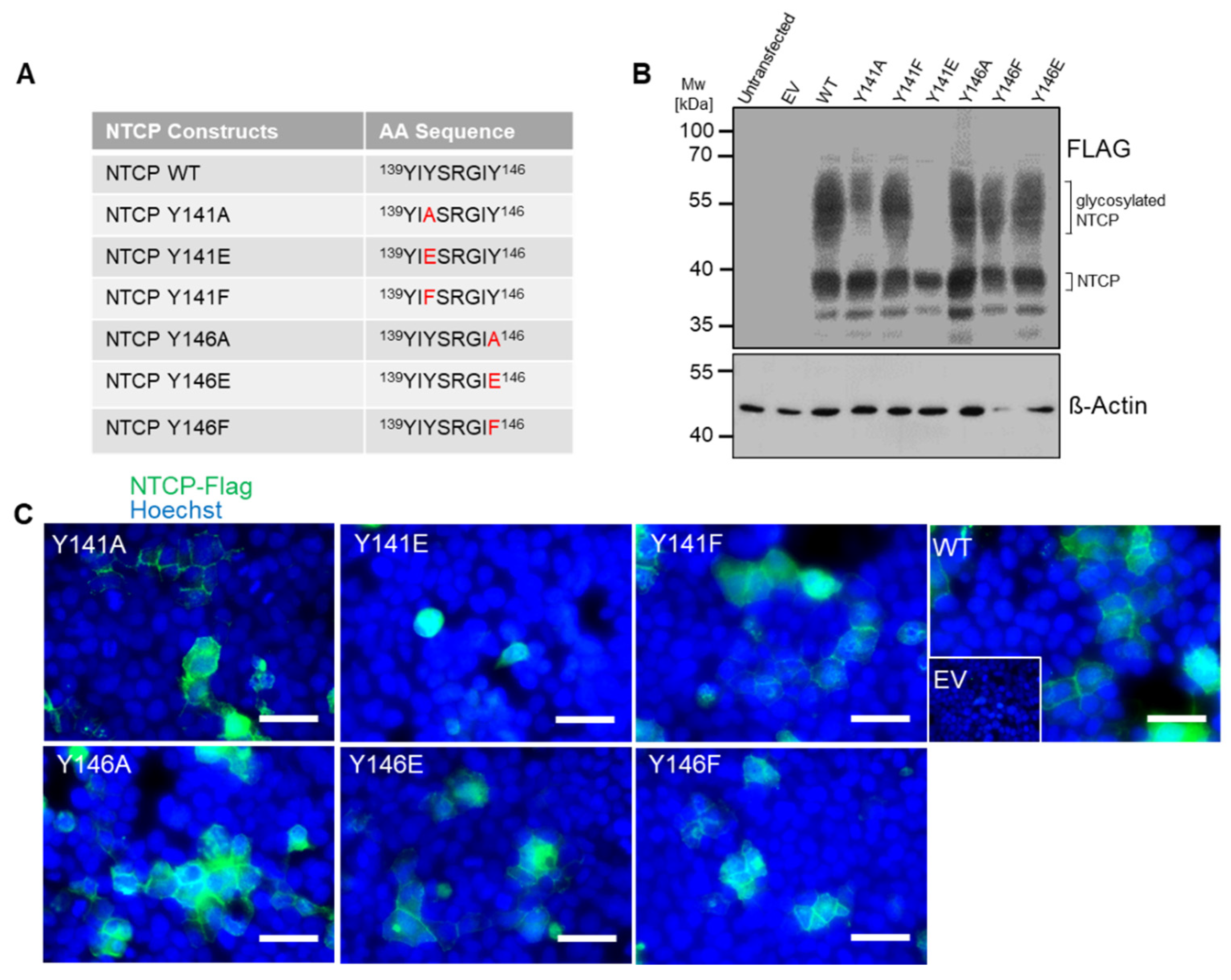
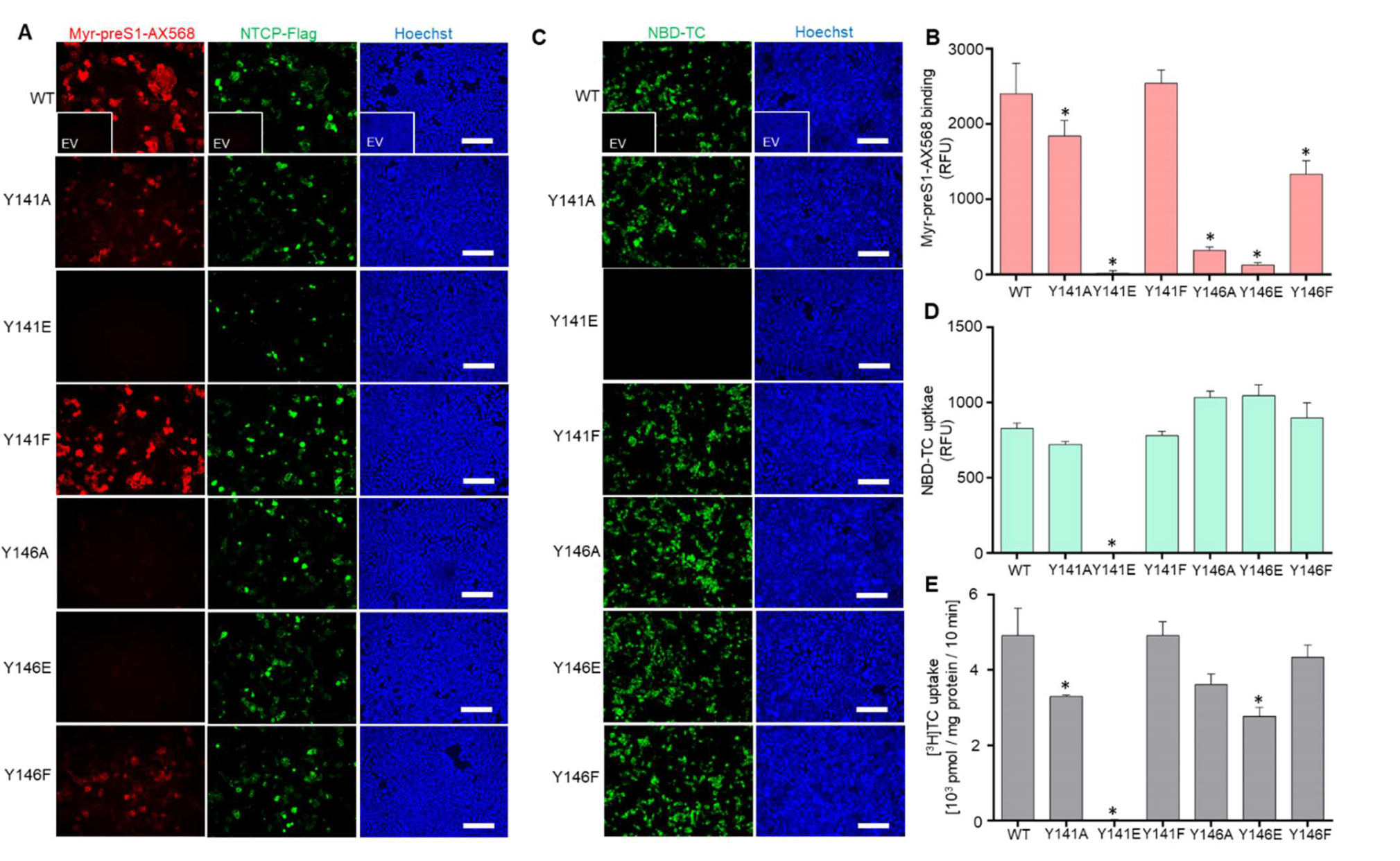
3.2. The Role of Y146 and Y141 in the Transporter and Receptor Function of NTCP
3.3. Y146A, Y146E, and Y146F Mutants Show Similar Transport Kinetics with WT NTCP
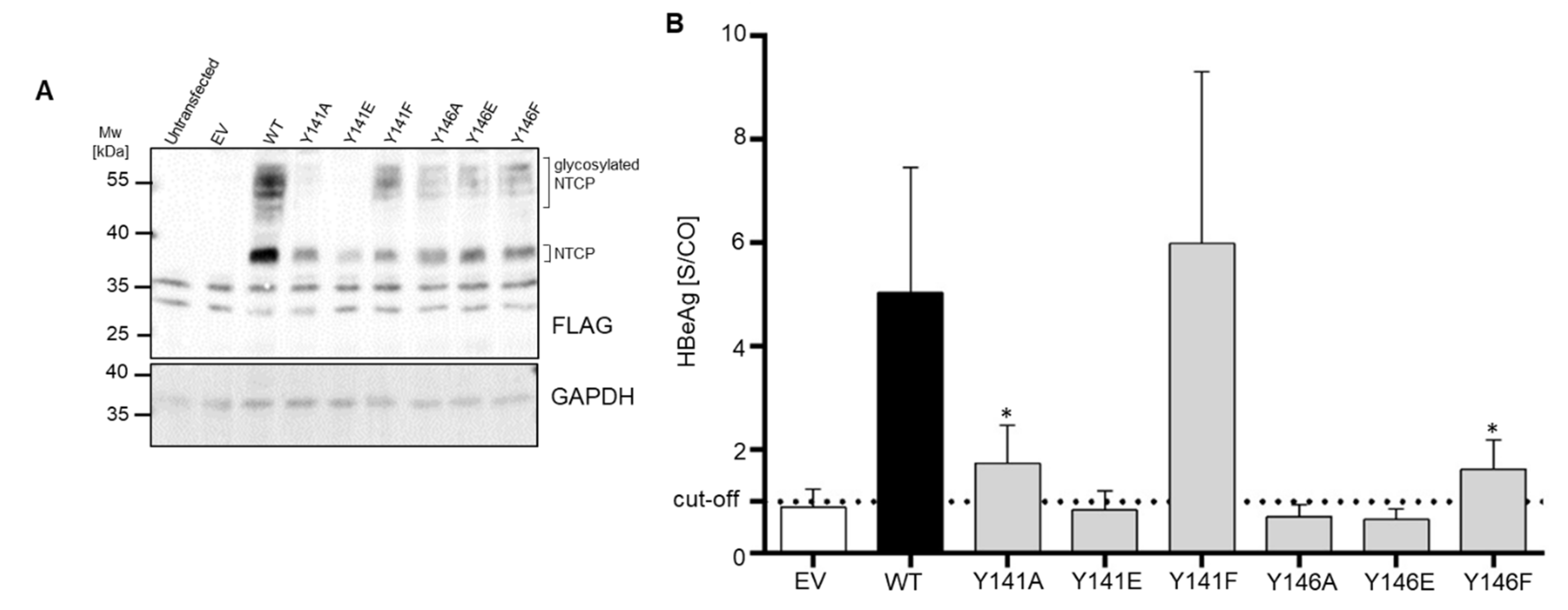
3.4. NTCP Y141E, Y146A and Y146E Mutations Protect against In Vitro HBV Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, J.; Wilke, T.; Petzinger, E. The solute carrier family SLC10: More than a family of bile acid transporters regarding function and phylogenetic relationships. Naunyn-Schmiedebergs Arch. Pharmakol. 2006, 372, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Döring, B.; Lütteke, T.; Geyer, J.; Petzinger, E. The SLC10 carrier family. Curr. Top Membr. 2012, 70, 105–168. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Fälth, M.; Stindt, J.; Königer, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 2014, 146, 1070–1083. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.-F.; Chen, D.-S.; Dusheiko, G.M.; Janssen, H.L.A.; Lau, D.T.Y.; Locarnini, S.A.; Peters, M.G.; Lai, C.-L. Hepatitis B virus infection. Nat. Rev. Dis. Prim. 2018, 4, 18035. [Google Scholar] [CrossRef]
- Urban, S.; Bartenschlager, R.; Kubitz, R.; Zoulim, F. Strategies to inhibit entry of HBV and HDV into hepatocytes. Gastroenterology 2014, 147, 48–64. [Google Scholar] [CrossRef]
- Rasche, A.; Sander, A.-L.; Corman, V.M.; Drexler, J.F. Evolutionary biology of human hepatitis viruses. J. Hepatol. 2019, 70, 501–520. [Google Scholar] [CrossRef] [Green Version]
- Roca Suarez, A.A.; Testoni, B.; Zoulim, F. HBV 2021: New therapeutic strategies against an old foe. Liver Int. 2021, 41 (Suppl. 1), 15–23. [Google Scholar] [CrossRef]
- Appelman, M.D.; Wettengel, J.M.; Protzer, U.; Oude Elferink, R.P.J.; van de Graaf, S.F.J. Molecular regulation of the hepatic bile acid uptake transporter and HBV entry receptor NTCP. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158960. [Google Scholar] [CrossRef]
- Zakrzewicz, D.; Geyer, J. Multitasking Na+/Taurocholate Cotransporting Polypeptide (NTCP) as a Drug Target for HBV Infection: From Protein Engineering to Drug Discovery. Biomedicines 2022, 10, 196. [Google Scholar] [CrossRef]
- Müller, S.F.; König, A.; Döring, B.; Glebe, D.; Geyer, J. Characterisation of the hepatitis B virus cross-species transmission pattern via Na+/taurocholate co-transporting polypeptides from 11 New World and Old World primate species. PLoS ONE 2018, 13, e0199200. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Peng, B.; He, W.; Zhong, G.; Qi, Y.; Ren, B.; Gao, Z.; Jing, Z.; Song, M.; Xu, G.; et al. Molecular determinants of hepatitis b and d virus entry restriction in mouse sodium taurocholate cotransporting polypeptide. J. Virol. 2013, 87, 7977–7991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-H.; Iwamoto, M.; Yun, J.-H.; Uchikubo-Kamo, T.; Son, D.; Jin, Z.; Yoshida, H.; Ohki, M.; Ishimoto, N.; Mizutani, K.; et al. Structural insights into the HBV receptor and bile acid transporter NTCP. Nature 2022. [Google Scholar] [CrossRef] [PubMed]
- Goutam, K.; Ielasi, F.S.; Pardon, E.; Steyaert, J.; Reyes, N. Structural basis of sodium-dependent bile salt uptake into the liver. Nature 2022. [Google Scholar] [CrossRef] [PubMed]
- Asami, J.; Kimura, K.T.; Fujita-Fujiharu, Y.; Ishida, H.; Zhang, Z.; Nomura, Y.; Liu, K.; Uemura, T.; Sato, Y.; Ono, M.; et al. Structure of bile acid transporter NTCP crucial for hepatitis B virus entry. Nature 2022. [Google Scholar] [CrossRef] [PubMed]
- König, A.; Döring, B.; Mohr, C.; Geipel, A.; Geyer, J.; Glebe, D. Kinetics of the bile acid transporter and hepatitis B virus receptor Na+/taurocholate cotransporting polypeptide (NTCP) in hepatocytes. J. Hepatol. 2014, 61, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Peng, B.; Liu, Y.; Xu, G.; He, W.; Ren, B.; Jing, Z.; Sui, J.; Li, W. Viral Entry of hepatitis B and D viruses and bile salts transportation share common molecular determinants on sodium taurocholate cotransporting polypeptide. J. Virol. 2014, 88, 3273–3284. [Google Scholar] [CrossRef] [Green Version]
- Uchida, T.; Park, S.B.; Inuzuka, T.; Zhang, M.; Allen, J.N.; Chayama, K.; Liang, T.J. Genetically edited hepatic cells expressing the NTCP-S267F variant are resistant to hepatitis B virus infection. Mol. Ther. Methods Clin. Dev. 2021, 23, 597–605. [Google Scholar] [CrossRef]
- Binh, M.T.; Hoan, N.X.; Van Tong, H.; Sy, B.T.; Trung, N.T.; Bock, C.-T.; Toan, N.L.; Song, L.H.; Bang, M.H.; Meyer, C.G.; et al. NTCP S267F variant associates with decreased susceptibility to HBV and HDV infection and decelerated progression of related liver diseases. Int. J. Infect. Dis. 2019, 80, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xu, G.; Gao, Z.; Zhou, Z.; Guo, G.; Li, D.; Jing, Z.; Sui, J.; Li, W. The p.Ser267Phe variant of sodium taurocholate cotransporting polypeptide (NTCP) supports HBV infection with a low efficiency. Virology 2018, 522, 168–176. [Google Scholar] [CrossRef]
- Pan, W.; Song, I.-S.; Shin, H.-J.; Kim, M.-H.; Choi, Y.-L.; Lim, S.-J.; Kim, W.-Y.; Lee, S.-S.; Shin, J.-G. Genetic polymorphisms in Na+-taurocholate co-transporting polypeptide (NTCP) and ileal apical sodium-dependent bile acid transporter (ASBT) and ethnic comparisons of functional variants of NTCP among Asian populations. Xenobiotica 2011, 41, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Palatini, M.; Müller, S.F.; Lowjaga, K.A.A.T.; Noppes, S.; Alber, J.; Lehmann, F.; Goldmann, N.; Glebe, D.; Geyer, J. Mutational Analysis of the GXXXG/A Motifs in the Human Na+/Taurocholate Co-Transporting Polypeptide NTCP on Its Bile Acid Transport Function and Hepatitis B/D Virus Receptor Function. Front. Mol. Biosci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Fukano, K.; Oshima, M.; Tsukuda, S.; Aizaki, H.; Ohki, M.; Park, S.-Y.; Wakita, T.; Wakae, K.; Watashi, K.; Muramatsu, M. NTCP Oligomerization Occurs Downstream of the NTCP-EGFR Interaction during Hepatitis B Virus Internalization. J. Virol. 2021, 95, e0093821. [Google Scholar] [CrossRef] [PubMed]
- Noppes, S.; Müller, S.F.; Bennien, J.; Holtemeyer, M.; Palatini, M.; Leidolf, R.; Alber, J.; Geyer, J. Homo- and heterodimerization is a common feature of the solute carrier family SLC10 members. Biol. Chem. 2019, 400, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.H.; Pires, D.E.; Ascher, D.B. DynaMut2: Assessing changes in stability and flexibility upon single and multiple point missense mutations. Protein Sci. 2021, 30, 60–69. [Google Scholar] [CrossRef]
- Petzinger, E.; Wickboldt, A.; Pagels, P.; Starke, D.; Kramer, W. Hepatobiliary transport of bile acid amino acid, bile acid peptide, and bile acid oligonucleotide conjugates in rats. Hepatology 1999, 30, 1257–1268. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Czuba, L.; Hillgren, K.M.; Swaan, P.W. Post-translational modifications of transporters. Pharmacol. Ther. 2018, 192, 88–99. [Google Scholar] [CrossRef]
- Schwartz, D.; Gygi, S.P. An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets. Nat. Biotechnol. 2005, 23, 1391–1398. [Google Scholar] [CrossRef]
- Amanchy, R.; Periaswamy, B.; Mathivanan, S.; Reddy, R.; Tattikota, S.G.; Pandey, A. A curated compendium of phosphorylation motifs. Nat. Biotechnol. 2007, 25, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Saso, W.; Sugiyama, R.; Ishii, K.; Ohki, M.; Nagamori, S.; Suzuki, R.; Aizaki, H.; Ryo, A.; Yun, J.-H.; et al. Epidermal growth factor receptor is a host-entry cofactor triggering hepatitis B virus internalization. Proc. Natl. Acad. Sci. USA 2019, 116, 8487–8492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, J.S.; Fukano, K.; Iwamoto, M.; Tsukuda, S.; Suzuki, R.; Aizaki, H.; Muramatsu, M.; Wakita, T.; Sureau, C.; Watashi, K. A Single Adaptive Mutation in Sodium Taurocholate Cotransporting Polypeptide Induced by Hepadnaviruses Determines Virus Species Specificity. J. Virol. 2019, 93, e01432-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowjaga, K.A.A.; Kirstgen, M.; Müller, S.F.; Goldmann, N.; Lehmann, F.; Glebe, D.; Geyer, J. Long-term trans-inhibition of the hepatitis B and D virus receptor NTCP by taurolithocholic acid. Am. J. Physiol.-Gastrointest. Liver Physiol. 2021, 320, G66–G80. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Urban, S. Entry of hepatitis B and hepatitis D virus into hepatocytes: Basic insights and clinical implications. J. Hepatol. 2016, 64, S32–S40. [Google Scholar] [CrossRef]
- Ruggiero, M.J.; Malhotra, S.; Fenton, A.W.; Swint-Kruse, L.; Karanicolas, J.; Hagenbuch, B. A clinically relevant polymorphism in the Na+/taurocholate cotransporting polypeptide (NTCP) occurs at a rheostat position. J. Biol. Chem. 2021, 296, 100047. [Google Scholar] [CrossRef]
- Zhou, X.; Levin, E.J.; Pan, Y.; McCoy, J.G.; Sharma, R.; Kloss, B.; Bruni, R.; Quick, M.; Zhou, M. Structural basis of the alternating-access mechanism in a bile acid transporter. Nature 2014, 505, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lyu, Y.; Ji, Y.; Sun, Z.; Zhou, X. Substrate binding in the bile acid transporter ASBTYf from Yersinia frederiksenii. Acta Crystallogr. Sect. D Struct. Biol. 2021, 77, 117–125. [Google Scholar] [CrossRef]
- Hu, N.-J.; Iwata, S.; Cameron, A.D.; Drew, D. Crystal structure of a bacterial homologue of the bile acid sodium symporter ASBT. Nature 2011, 478, 408–411. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′→3′) |
|---|---|
| Y139A/Y141A/Y146A * | For: GAT GCC TCT CCT CCT GGC CAT CGC CTC CAG GGG GAT CGC TGA TGG GGA CCT GAA GG Rev: CCT TCA GGT CCC CAT CAG CGA TCC CCC TGG AGG CGA TGG CCA GGA GGA GAG GCA TC |
| Y139A | For: GAT GCC TCT CCT CCT GGC CAT CTA CTC CAG GGG G Rev: CCC CCT GGA GTA GAT GGC CAG GAG GAG AGA GGC ATC |
| Y141A | For: CCT CTC CTC CTG TAC ATC GCC TCC AGG GGG ATC TAT GATGGG Rev: CCC ATC ATA GAT CCC CCT GGA GGC GAT GTA CAG GAG GAG AGG |
| Y141E | For: CCT CTC CTC CTG TAC ATC GAG TCC AGG GGG ATC TAT GAT GGG Rev: CCC ATC ATA GAT CCC CCT GGA CTC GAT GTA CAG GAG GAG AGG |
| Y141F | For: CCT CTC CTC CTG TAC ATC TTC TCC AGG GGG ATC TAT GAT GGG Rev: CCC ATC ATA GAT CCC CCT GGA GAA GAT GTA CAG GAG GAG AGG |
| Y146A | For: CAT CTA CTC CAG GGG GAT CGC TGA TGG GGA CCT GAA GGA C Rev: GTC CTT CAG GTC CCC ATC AGC GAT CCC CCT GGA GTA GAT G |
| Y146E | For: CAT CTA CTC CAG GGG GAT CGA AGA TGG GGA CCT GAA GGA Rev: GTC CTT CAG GTC CCC ATC TTC GAT CCC CCT GGA GTA GAT G |
| Y146F | For: CAT CTA CTC CAG GGG GAT CTT TGA TGG GGA CCT GAA GGA C Rev: GTC CTT CAG GTC CCC ATC AAA GAT CCC CCT GGA GTA GAT G |
| R143A | For: CTC CTG TAC ATC TAC TCC GCG GGG ATC TAT GAT GGG GAC Rev: GTC CCC ATC ATA GAT CCC CGC GGA GTA GAT GTA CAG GAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakrzewicz, D.; Leidolf, R.; Kunz, S.; Müller, S.F.; Neubauer, A.; Leiting, S.; Goldmann, N.; Lehmann, F.; Glebe, D.; Geyer, J. Tyrosine 146 of the Human Na+/Taurocholate Cotransporting Polypeptide (NTCP) Is Essential for Its Hepatitis B Virus (HBV) Receptor Function and HBV Entry into Hepatocytes. Viruses 2022, 14, 1259. https://doi.org/10.3390/v14061259
Zakrzewicz D, Leidolf R, Kunz S, Müller SF, Neubauer A, Leiting S, Goldmann N, Lehmann F, Glebe D, Geyer J. Tyrosine 146 of the Human Na+/Taurocholate Cotransporting Polypeptide (NTCP) Is Essential for Its Hepatitis B Virus (HBV) Receptor Function and HBV Entry into Hepatocytes. Viruses. 2022; 14(6):1259. https://doi.org/10.3390/v14061259
Chicago/Turabian StyleZakrzewicz, Dariusz, Regina Leidolf, Sebastian Kunz, Simon Franz Müller, Anita Neubauer, Silke Leiting, Nora Goldmann, Felix Lehmann, Dieter Glebe, and Joachim Geyer. 2022. "Tyrosine 146 of the Human Na+/Taurocholate Cotransporting Polypeptide (NTCP) Is Essential for Its Hepatitis B Virus (HBV) Receptor Function and HBV Entry into Hepatocytes" Viruses 14, no. 6: 1259. https://doi.org/10.3390/v14061259