
Human Beta Papillomavirus Type 8 E1 and E2 Proteins Suppress the Activation of the RIG-I-Like Receptor MDA5

 , , and
, , and 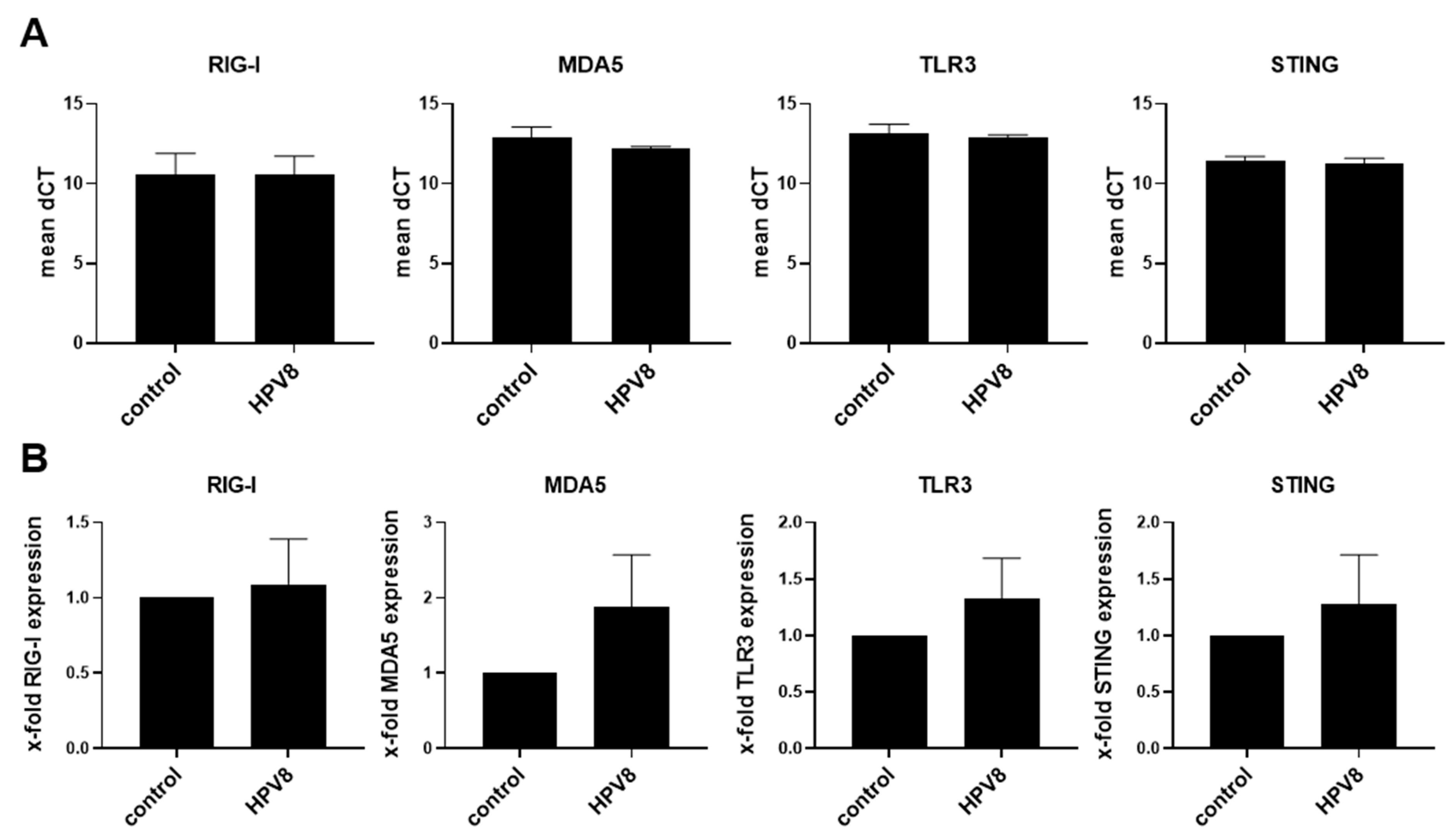{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Isolation and qPCR Analysis
2.3. Measurement of Cytokine/Chemokine Secretion
2.4. Cell Viability
2.5. Statistical Analysis
3. Results
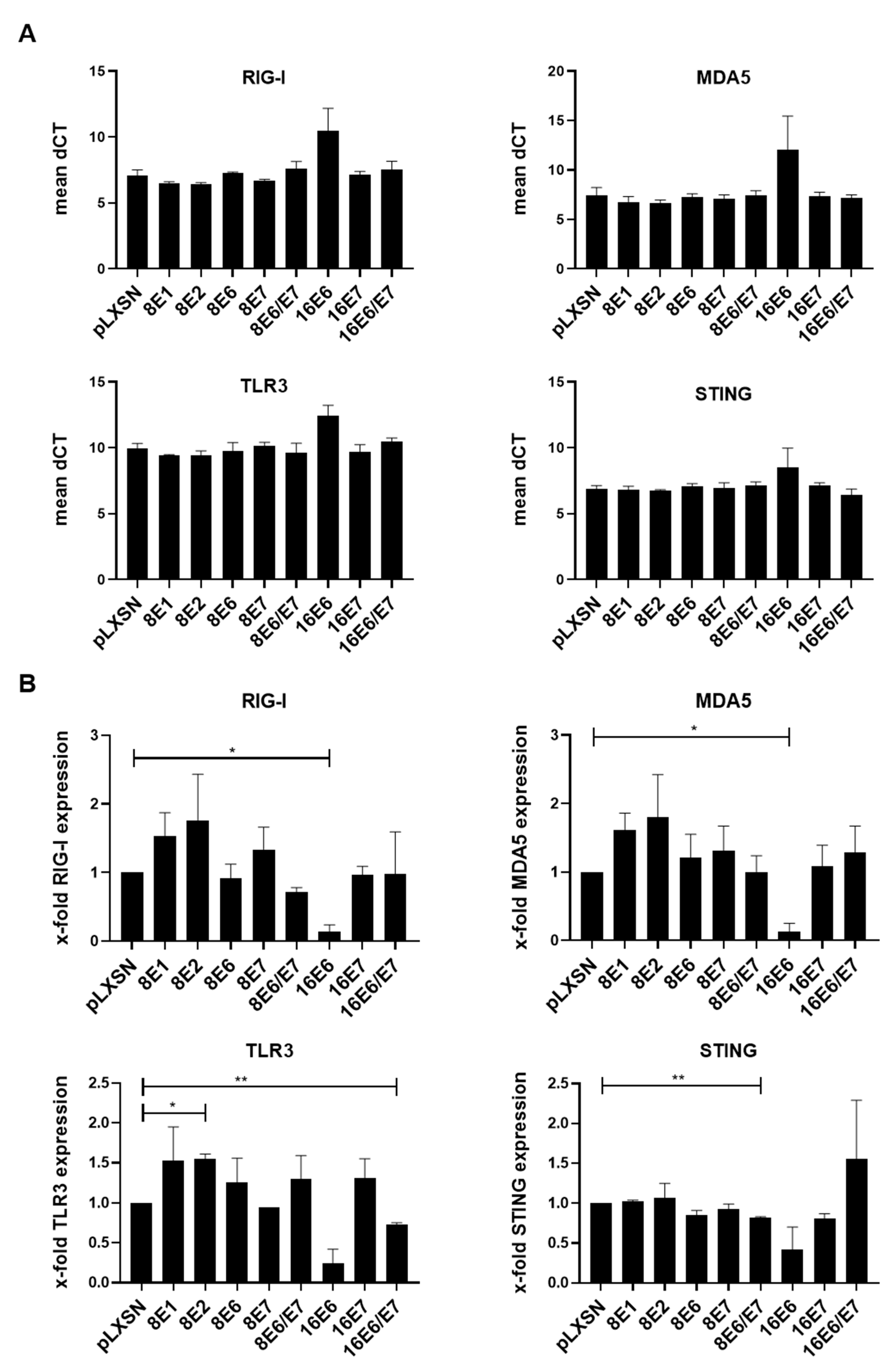
3.1. Expression Levels of RIG-I, MDA5, TLR3, and STING Are Downregulated by HPV16-E6 but Not by HPV8 Early Proteins
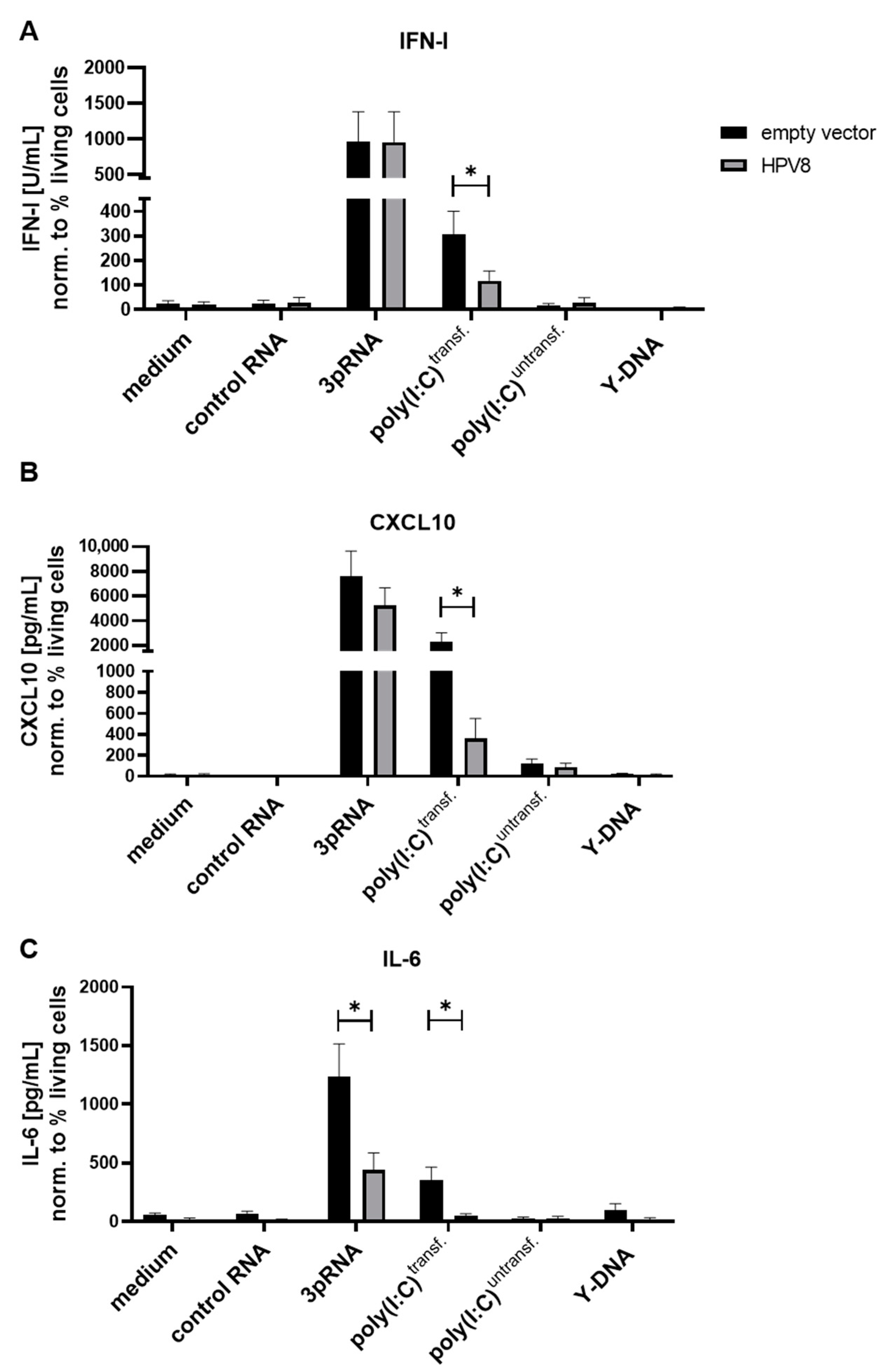
3.2. MDA5-Induced Cytokine Secretion in U2OS Cells Is Diminished in the Presence of HPV8
3.3. Expression of HPV8-E1 and HPV8-E2 Suppress MDA5-Induced Cytokines in Primary Human Keratinocytes
3.4. PBMCs Secret Elevated Cytokine Levels upon Stimulation with Total RNA Isolated from HPV8-E2-Expressing Keratinocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fera, A.N.D.; Warburton, A.; Coursey, T.L.; Khurana, S.; McBride, A.A. Persistent Human Papillomavirus Infection. Viruses 2021, 13, 321. [Google Scholar] [CrossRef]
- Van Doorslaer, K.; Li, Z.; Xirasagar, S.; Maes, P.; Kaminsky, D.; Liou, D.; Sun, Q.; Kaur, R.; Huyen, Y.; McBride, A.A. The Papillomavirus Episteme: A Major Update to the Papillomavirus Sequence Database. Nucleic Acids Res. 2017, 45, D499–D506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Villiers, E.-M.; Fauquet, C.; Broker, T.R.; Bernard, H.-U.; zur Hausen, H. Classification of Papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human Papillomavirus Molecular Biology and Disease Association. Rev. Med. Virol. 2015, 25, 2–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffman, M.; Doorbar, J.; Wentzensen, N.; de Sanjosé, S.; Fakhry, C.; Monk, B.J.; Stanley, M.A.; Franceschi, S. Carcinogenic Human Papillomavirus Infection. Nat. Rev. Dis. Primers 2016, 2, 16086. [Google Scholar] [CrossRef]
- Hübbers, C.U.; Akgül, B. HPV and Cancer of the Oral Cavity. Virulence 2015, 6, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Howley, P.M.; Pfister, H.J. Beta Genus Papillomaviruses and Skin Cancer. Virology 2015, 479, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Hufbauer, M.; Akgül, B. Molecular Mechanisms of Human Papillomavirus Induced Skin Carcinogenesis. Viruses 2017, 9, 187. [Google Scholar] [CrossRef] [Green Version]
- Tommasino, M. The Biology of Beta Human Papillomaviruses. Virus Res. 2017, 231, 128–138. [Google Scholar] [CrossRef]
- Stanley, M. Pathology and Epidemiology of HPV Infection in Females. Gynecol. Oncol. 2010, 117, S5–S10. [Google Scholar] [CrossRef]
- Egawa, N.; Doorbar, J. The Low-Risk Papillomaviruses. Virus Res. 2017, 231, 119–127. [Google Scholar] [CrossRef] [PubMed]
- McBride, A.A.; Sakakibara, N.; Stepp, W.H.; Jang, M.K. Hitchhiking on Host Chromatin: How Papillomaviruses Persist. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venuti, A.; Lohse, S.; Tommasino, M.; Smola, S. Cross-Talk of Cutaneous Beta Human Papillomaviruses and the Immune System: Determinants of Disease Penetrance. Philos. Trans. R. Soc. B 2019, 374, 20180287. [Google Scholar] [CrossRef] [Green Version]
- Okude, H.; Ori, D.; Kawai, T. Signaling through Nucleic Acid Sensors and Their Roles in Inflammatory Diseases. Front. Immunol. 2021, 11, 625833. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Barchet, W.; Wimmenauer, V.; Schlee, M.; Hartmann, G. Accessing the Therapeutic Potential of Immunostimulatory Nucleic Acids. Curr. Opin. Immunol. 2008, 20, 389–395. [Google Scholar] [CrossRef]
- Kalali, B.N.; Köllisch, G.; Mages, J.; Müller, T.; Bauer, S.; Wagner, H.; Ring, J.; Lang, R.; Mempel, M.; Ollert, M. Double-Stranded RNA Induces an Antiviral Defense Status in Epidermal Keratinocytes through TLR3-, PKR-, and MDA5/RIG-I-Mediated Differential Signaling. J. Immunol. 2008, 181, 2694–2704. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, G.; Krieg, A.M. Mechanism and Function of a Newly Identified CpG DNA Motif in Human Primary B Cells. J. Immunol. 2000, 164, 944–953. [Google Scholar] [CrossRef]
- Krug, A.; Rothenfusser, S.; Hornung, V.; Jahrsdörfer, B.; Blackwell, S.; Ballas, Z.K.; Endres, S.; Krieg, A.M.; Hartmann, G. Identification of CpG Oligonucleotide Sequences with High Induction of IFN-α/β in Plasmacytoid Dendritic Cells. Eur. J. Immunol. 2001, 31, 2154–2163. [Google Scholar] [CrossRef]
- Hornung, V.; Guenthner-Biller, M.; Bourquin, C.; Ablasser, A.; Schlee, M.; Uematsu, S.; Noronha, A.; Manoharan, M.; Akira, S.; de Fougerolles, A.; et al. Sequence-Specific Potent Induction of IFN-α by Short Interfering RNA in Plasmacytoid Dendritic Cells through TLR7. Nat. Med. 2005, 11, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Bartok, E.; Hartmann, G. Immune Sensing Mechanisms That Discriminate Self from Altered Self and Foreign Nucleic Acids. Immunity 2020, 53, 54–77. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, G. Chapter Four Nucleic Acid Immunity. Adv. Immunol. 2017, 133, 121–169. [Google Scholar] [CrossRef]
- Schlee, M.; Roth, A.; Hornung, V.; Hagmann, C.A.; Wimmenauer, V.; Barchet, W.; Coch, C.; Janke, M.; Mihailovic, A.; Wardle, G.; et al. Recognition of 5′ Triphosphate by RIG-I Helicase Requires Short Blunt Double-Stranded RNA as Contained in Panhandle of Negative-Strand Virus. Immunity 2009, 31, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Schwerd, T.; Hamm, W.; Hellmuth, J.C.; Cui, S.; Wenzel, M.; Hoffmann, F.S.; Michallet, M.-C.; Besch, R.; Hopfner, K.-P.; et al. 5′-Triphosphate RNA Requires Base-Paired Structures to Activate Antiviral Signaling via RIG-I. Proc. Natl. Acad. Sci. USA 2009, 106, 12067–12072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlee, M. Master Sensors of Pathogenic RNA–RIG-I like Receptors. Immunobiology 2013, 218, 1322–1335. [Google Scholar] [CrossRef]
- Gitlin, L.; Barchet, W.; Gilfillan, S.; Cella, M.; Beutler, B.; Flavell, R.A.; Diamond, M.S.; Colonna, M. Essential Role of Mda-5 in Type I IFN Responses to Polyriboinosinic: Polyribocytidylic Acid and Encephalomyocarditis Picornavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 8459–8464. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Uematsu, S.; Matsui, K.; Tsujimura, T.; Takeda, K.; Fujita, T.; Takeuchi, O.; et al. Cell Type-Specific Involvement of RIG-I in Antiviral Response. Immunity 2005, 23, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential Roles of MDA5 and RIG-I Helicases in the Recognition of RNA Viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Kato, H.; Takeuchi, O.; Mikamo-Satoh, E.; Hirai, R.; Kawai, T.; Matsushita, K.; Hiiragi, A.; Dermody, T.S.; Fujita, T.; Akira, S. Length-Dependent Recognition of Double-Stranded Ribonucleic Acids by Retinoic Acid–Inducible Gene-I and Melanoma Differentiation–Associated Gene 5. J. Exp. Med. 2008, 205, 1601–1610. [Google Scholar] [CrossRef]
- Yoneyama, M.; Kikuchi, M.; Matsumoto, K.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Foy, E.; Loo, Y.-M.; Gale, M.; Akira, S.; et al. Shared and Unique Functions of the DExD/H-Box Helicases RIG-I, MDA5, and LGP2 in Antiviral Innate Immunity. J. Immunol. 2005, 175, 2851–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP Synthase Is a Cytosolic DNA Sensor That Activates the Type I Interferon Pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Sun, L.; Chen, X.; Du, F.; Shi, H.; Chen, C.; Chen, Z.J. Cyclic GMP-AMP Is an Endogenous Second Messenger in Innate Immune Signaling by Cytosolic DNA. Science 2013, 339, 826–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Ascano, M.; Wu, Y.; Barchet, W.; Gaffney, B.L.; Zillinger, T.; Serganov, A.A.; Liu, Y.; Jones, R.A.; Hartmann, G.; et al. Cyclic [G(2′,5′)PA(3′,5′)p] Is the Metazoan Second Messenger Produced by DNA-Activated Cyclic GMP-AMP Synthase. Cell 2013, 153, 1094–1107. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Bruneau, R.C.; Brennan, G.; Rothenburg, S. Battle Royale: Innate Recognition of Poxviruses and Viral Immune Evasion. Biomed 2021, 9, 765. [Google Scholar] [CrossRef]
- Ablasser, A.; Goldeck, M.; Cavlar, T.; Deimling, T.; Witte, G.; Röhl, I.; Hopfner, K.-P.; Ludwig, J.; Hornung, V. CGAS Produces a 2′-5′-Linked Cyclic Dinucleotide Second Messenger that Activates STING. Nature 2013, 498, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Cigno, I.L.; Calati, F.; Albertini, S.; Gariglio, M. Subversion of Host Innate Immunity by Human Papillomavirus Oncoproteins. Pathogens 2020, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Reiser, J.; Hurst, J.; Voges, M.; Krauss, P.; Münch, P.; Iftner, T.; Stubenrauch, F. High-Risk Human Papillomaviruses Repress Constitutive Kappa Interferon Transcription via E6 to Prevent Pathogen Recognition Receptor and Antiviral-Gene Expression. J. Virol. 2011, 85, 11372–11380. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.; Pauli, E.-K.; Biryukov, J.; Feister, K.F.; Meng, M.; White, E.A.; Münger, K.; Howley, P.M.; Meyers, C.; Gack, M.U. The Human Papillomavirus E6 Oncoprotein Targets USP15 and TRIM25 to Suppress RIG-I-Mediated Innate Immune Signaling. J. Virol. 2018, 92, e01737-17. [Google Scholar] [CrossRef] [Green Version]
- Ronco, L.V.; Karpova, A.Y.; Vidal, M.; Howley, P.M. Human Papillomavirus 16 E6 Oncoprotein Binds to Interferon Regulatory Factor-3 and Inhibits Its Transcriptional Activity. Gene Dev. 1998, 12, 2061–2072. [Google Scholar] [CrossRef] [Green Version]
- Perea, S.E.; Massimi, P.; Banks, L. Human Papillomavirus Type 16 E7 Impairs the Activation of the Interferon Regulatory Factor-1. Int. J. Mol. Med. 2000, 5, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Um, S.-J.; Rhyu, J.-W.; Kim, E.-J.; Jeon, K.-C.; Hwang, E.-S.; Park, J.-S. Abrogation of IRF-1 Response by High-Risk HPV E7 Protein in Vivo. Cancer Lett. 2002, 179, 205–212. [Google Scholar] [CrossRef]
- Park, J.-S.; Kim, E.-J.; Kwon, H.-J.; Hwang, E.-S.; Namkoong, S.-E.; Um, S.-J. Inactivation of Interferon Regulatory Factor-1 Tumor Suppressor Protein by HPV E7 Oncoprotein implication for the e7-mediated immune evasion mechanism in cervical carcinogenesis. J. Biol. Chem. 2000, 275, 6764–6769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, U.A.; Zannetti, C.; Parroche, P.; Goutagny, N.; Malfroy, M.; Roblot, G.; Carreira, C.; Hussain, I.; Müller, M.; Taylor-Papadimitriou, J.; et al. The Human Papillomavirus Type 16 E7 Oncoprotein Induces a Transcriptional Repressor Complex on the Toll-like Receptor 9 Promoter. J. Exp. Med. 2013, 210, 1369–1387. [Google Scholar] [CrossRef]
- Luo, X.; Donnelly, C.R.; Gong, W.; Heath, B.R.; Hao, Y.; Donnelly, L.A.; Moghbeli, T.; Tan, Y.S.; Lin, X.; Bellile, E.; et al. HPV16 Drives Cancer Immune Escape via NLRX1-Mediated Degradation of STING. J. Clin. Invest. 2019, 130, 1635–1652. [Google Scholar] [CrossRef] [Green Version]
- Lau, L.; Gray, E.E.; Brunette, R.L.; Stetson, D.B. DNA Tumor Virus Oncogenes Antagonize the CGAS-STING DNA-Sensing Pathway. Science 2015, 350, 568–571. [Google Scholar] [CrossRef] [Green Version]
- Albertini, S.; Cigno, I.L.; Calati, F.; Andrea, M.D.; Borgogna, C.; Dell’Oste, V.; Landolfo, S.; Gariglio, M. HPV18 Persistence Impairs Basal and DNA Ligand–Mediated IFN-β and IFN-Λ1 Production through Transcriptional Repression of Multiple Downstream Effectors of Pattern Recognition Receptor Signaling. J. Immunol. 2018, 200, 2076–2089. [Google Scholar] [CrossRef] [Green Version]
- Cigno, I.L.; Calati, F.; Borgogna, C.; Zevini, A.; Albertini, S.; Martuscelli, L.; Andrea, M.D.; Hiscott, J.; Landolfo, S.; Gariglio, M. Human Papillomavirus E7 Oncoprotein Subverts Host Innate Immunity via SUV39H1-Mediated Epigenetic Silencing of Immune Sensor Genes. J. Virol. 2020, 94, e01812-19. [Google Scholar] [CrossRef]
- Heuser, S.; Hufbauer, M.; Steiger, J.; Marshall, J.; Sterner-Kock, A.; Mauch, C.; Zigrino, P.; Akgül, B. The Fibronectin/A3β1 Integrin Axis Serves as Molecular Basis for Keratinocyte Invasion Induced by ΒHPV. Oncogene 2016, 35, 4529–4539. [Google Scholar] [CrossRef]
- Marcuzzi, G.P.; Hufbauer, M.; Kasper, H.U.; Weißenborn, S.J.; Smola, S.; Pfister, H. Spontaneous Tumour Development in Human Papillomavirus Type 8 E6 Transgenic Mice and Rapid Induction by UV-Light Exposure and Wounding. J. Gen. Virol. 2009, 90, 2855–2864. [Google Scholar] [CrossRef]
- Pfefferle, R.; Marcuzzi, G.P.; Akgül, B.; Kasper, H.U.; Schulze, F.; Haase, I.; Wickenhauser, C.; Pfister, H. The Human Papillomavirus Type 8 E2 Protein Induces Skin Tumors in Transgenic Mice. J. Investig. Dermatol. 2008, 128, 2310–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akgül, B.; Bostanci, N.; Westphal, K.; Nindl, I.; Navsaria, H.; Storey, A.; Pfister, H. Human Papillomavirus 5 and 8 E6 Downregulate Interleukin-8 Secretion in Primary Human Keratinocytes. J. Gen. Virol. 2010, 91, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Pacini, L.; Ceraolo, M.G.; Venuti, A.; Melita, G.; Hasan, U.A.; Accardi, R.; Tommasino, M. UV Radiation Activates Toll-Like Receptor 9 Expression in Primary Human Keratinocytes, an Event Inhibited by Human Papillomavirus 38 E6 and E7 Oncoproteins. J. Virol. 2017, 91, e01123-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geimanen, J.; Isok-Paas, H.; Pipitch, R.; Salk, K.; Laos, T.; Orav, M.; Reinson, T.; Ustav, M.; Ustav, M.; Ustav, E. Development of a Cellular Assay System to Study the Genome Replication of High- and Low-Risk Mucosal and Cutaneous Human Papillomaviruses. J. Virol. 2011, 85, 3315–3329. [Google Scholar] [CrossRef] [Green Version]
- Hufbauer, M.; Biddle, A.; Borgogna, C.; Gariglio, M.; Doorbar, J.; Storey, A.; Pfister, H.; Mackenzie, I.; Akgül, B. Expression of Betapapillomavirus Oncogenes Increases the Number of Keratinocytes with Stem Cell-Like Properties. J. Virol. 2013, 87, 12158–12165. [Google Scholar] [CrossRef] [Green Version]
- Akgül, B.; Ghali, L.; Davies, D.; Pfister, H.; Leigh, I.M.; Storey, A. HPV8 Early Genes Modulate Differentiation and Cell Cycle of Primary Human Adult Keratinocytes. Exp. Dermatol. 2007, 16, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Leverrier, S.; Bergamaschi, D.; Ghali, L.; Ola, A.; Warnes, G.; Akgül, B.; Blight, K.; García-Escudero, R.; Penna, A.; Eddaoudi, A.; et al. Role of HPV E6 Proteins in Preventing UVB-Induced Release of pro-Apoptotic Factors from the Mitochondria. Apoptosis 2007, 12, 549–560. [Google Scholar] [CrossRef]
- Akgül, B.; García-Escudero, R.; Ghali, L.; Pfister, H.J.; Fuchs, P.G.; Navsaria, H.; Storey, A. The E7 Protein of Cutaneous Human Papillomavirus Type 8 Causes Invasion of Human Keratinocytes into the Dermis in Organotypic Cultures of Skin. Cancer Res. 2005, 65, 2216–2223. [Google Scholar] [CrossRef] [Green Version]
- Akgül, B.; García-Escudero, R.; Ekechi, C.; Steger, G.; Navsaria, H.; Pfister, H.; Storey, A. The E2 Protein of Human Papillomavirus Type 8 Increases the Expression of Matrix Metalloproteinase-9 in Human Keratinocytes and Organotypic Skin Cultures. Med. Microbiol. Immunol. 2011, 200, 127–135. [Google Scholar] [CrossRef]
- Daßler-Plenker, J.; Paschen, A.; Putschli, B.; Rattay, S.; Schmitz, S.; Goldeck, M.; Bartok, E.; Hartmann, G.; Coch, C. Direct RIG-I Activation in Human NK Cells Induces TRAIL-dependent Cytotoxicity toward Autologous Melanoma Cells. Int. J. Cancer 2019, 144, 1645–1656. [Google Scholar] [CrossRef]
- Glas, M.; Coch, C.; Trageser, D.; Daßler, J.; Simon, M.; Koch, P.; Mertens, J.; Quandel, T.; Gorris, R.; Reinartz, R.; et al. Targeting the Cytosolic Innate Immune Receptors RIG-I and MDA5 Effectively Counteracts Cancer Cell Heterogeneity in Glioblastoma. Stem Cells 2013, 31, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Isok-Paas, H.; Männik, A.; Ustav, E.; Ustav, M. The Transcription Map of HPV11 in U2OS Cells Adequately Reflects the Initial and Stable Replication Phases of the Viral Genome. Virol. J. 2015, 12, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschamps, T.; Kalamvoki, M. Impaired STING Pathway in Human Osteosarcoma U2OS Cells Contributes to the Growth of ICP0-Null Mutant Herpes Simplex Virus. J. Virol. 2017, 91, e00006-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, C.D.; Fontan, C.T.; Otoa, R.; Das, D.; Prabhakar, A.T.; Wang, X.; Bristol, M.L.; Morgan, I.M. Human Papillomavirus 16 E6 and E7 Synergistically Repress Innate Immune Gene Transcription. Msphere 2020, 5, e00828-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Koning, H.D.; Rodijk-Olthuis, D.; van Vlijmen-Willems, I.M.J.J.; Joosten, L.A.B.; Netea, M.G.; Schalkwijk, J.; Zeeuwen, P.L.J.M. A Comprehensive Analysis of Pattern Recognition Receptors in Normal and Inflamed Human Epidermis: Upregulation of Dectin-1 in Psoriasis. J. Investig. Dermatol. 2010, 130, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Matsuzaki, Y.; Nishikawa, Y.; Kitamura, H.; Akasaka, E.; Rokunohe, D.; Nakano, H.; Imaizumi, T.; Satoh, K.; Sawamura, D. Characterization of Retinoic Acid-Inducible Gene-I (RIG-I) Expression Corresponding to Viral Infection and UVB in Human Keratinocytes. J. Dermatol. Sci. 2012, 66, 64–70. [Google Scholar] [CrossRef]
- Lebre, M.C.; van der Aar, A.M.G.; van Baarsen, L.; van Capel, T.M.M.; Schuitemaker, J.H.N.; Kapsenberg, M.L.; de Jong, E.C. Human Keratinocytes Express Functional Toll-Like Receptor 3, 4, 5, and 9. J. Investig. Dermatol. 2007, 127, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Köllisch, G.; Kalali, B.N.; Voelcker, V.; Wallich, R.; Behrendt, H.; Ring, J.; Bauer, S.; Jakob, T.; Mempel, M.; Ollert, M. Various Members of the Toll-like Receptor Family Contribute to the Innate Immune Response of Human Epidermal Keratinocytes. Immunology 2005, 114, 531–541. [Google Scholar] [CrossRef]
- Castro-Munoz, L.J.; Manzo-Merino, J.; Munoz-Bello, J.O.; Olmedo-Nieva, L.; Cedro-Tanda, A.; Alfaro-Ruiz, L.A.; Hidalgo-Miranda, A.; Madrid-Marina, V.; Lizano, M. The Human Papillomavirus (HPV) E1 Protein Regulates the Expression of Cellular Genes Involved in Immune Response. Sci. Rep. 2019, 9, 13620. [Google Scholar] [CrossRef]
- Sunthamala, N.; Thierry, F.; Teissier, S.; Pientong, C.; Kongyingyoes, B.; Tangsiriwatthana, T.; Sangkomkamhang, U.; Ekalaksananan, T. E2 Proteins of High Risk Human Papillomaviruses Down-Modulate STING and IFN-κ Transcription in Keratinocytes. PLoS ONE 2014, 9, e91473. [Google Scholar] [CrossRef]
- Evans, M.R.; James, C.D.; Bristol, M.L.; Nulton, T.J.; Wang, X.; Kaur, N.; White, E.A.; Windle, B.; Morgan, I.M. Human Papillomavirus 16 E2 Regulates Keratinocyte Gene Expression Relevant to Cancer and the Viral Life Cycle. J. Virol. 2018, 93, e01941-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.R.; James, C.D.; Bristol, M.L.; Nulton, T.J.; Wang, X.; Kaur, N.; White, E.A.; Windle, B.; Morgan, I.M. Correction for Evans et al., “Human Papillomavirus 16 E2 Regulates Keratinocyte Gene Expression Relevant to Cancer and the Viral Life Cycle”. J. Virol. 2019, 93, e01067-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Gao, C. E3 Ubiquitin Ligases, the Powerful Modulator of Innate Antiviral Immunity. Cell. Immunol. 2019, 340, 103915. [Google Scholar] [CrossRef] [PubMed]
- Oshiumi, H.; Kouwaki, T.; Seya, T. Accessory Factors of Cytoplasmic Viral RNA Sensors Required for Antiviral Innate Immune Response. Front. Immunol. 2016, 7, 200. [Google Scholar] [CrossRef]
- Fuchs, P.G.; Iftner, T.; Weninger, J.; Pfister, H. Epidermodysplasia Verruciformis-Associated Human Papillomavirus 8: Genomic Sequence and Comparative Analysis. J. Virol. 1986, 58, 626–634. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rattay, S.; Hufbauer, M.; Hagen, C.; Putschli, B.; Coch, C.; Akgül, B.; Hartmann, G. Human Beta Papillomavirus Type 8 E1 and E2 Proteins Suppress the Activation of the RIG-I-Like Receptor MDA5. Viruses 2022, 14, 1361. https://doi.org/10.3390/v14071361
Rattay S, Hufbauer M, Hagen C, Putschli B, Coch C, Akgül B, Hartmann G. Human Beta Papillomavirus Type 8 E1 and E2 Proteins Suppress the Activation of the RIG-I-Like Receptor MDA5. Viruses. 2022; 14(7):1361. https://doi.org/10.3390/v14071361
Chicago/Turabian StyleRattay, Stephanie, Martin Hufbauer, Christian Hagen, Bastian Putschli, Christoph Coch, Baki Akgül, and Gunther Hartmann. 2022. "Human Beta Papillomavirus Type 8 E1 and E2 Proteins Suppress the Activation of the RIG-I-Like Receptor MDA5" Viruses 14, no. 7: 1361. https://doi.org/10.3390/v14071361