
Occurrence and Characterization of Wheat Streak Mosaic Virus Found in Mono- and Mixed Infection with High Plains Wheat Mosaic Virus in Winter Wheat in Ukraine

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Enzyme-Linked Immunosobent Assay (ELISA)
2.3. Transmission Electron Microscopy (TEM)
2.4. RNA Extraction, Reverse Transcription-Polymerase Chain Reaction (RT-PCR), and Sequencing
2.5. Phylogenetic Analysis
2.6. Statistical Analysis
3. Results
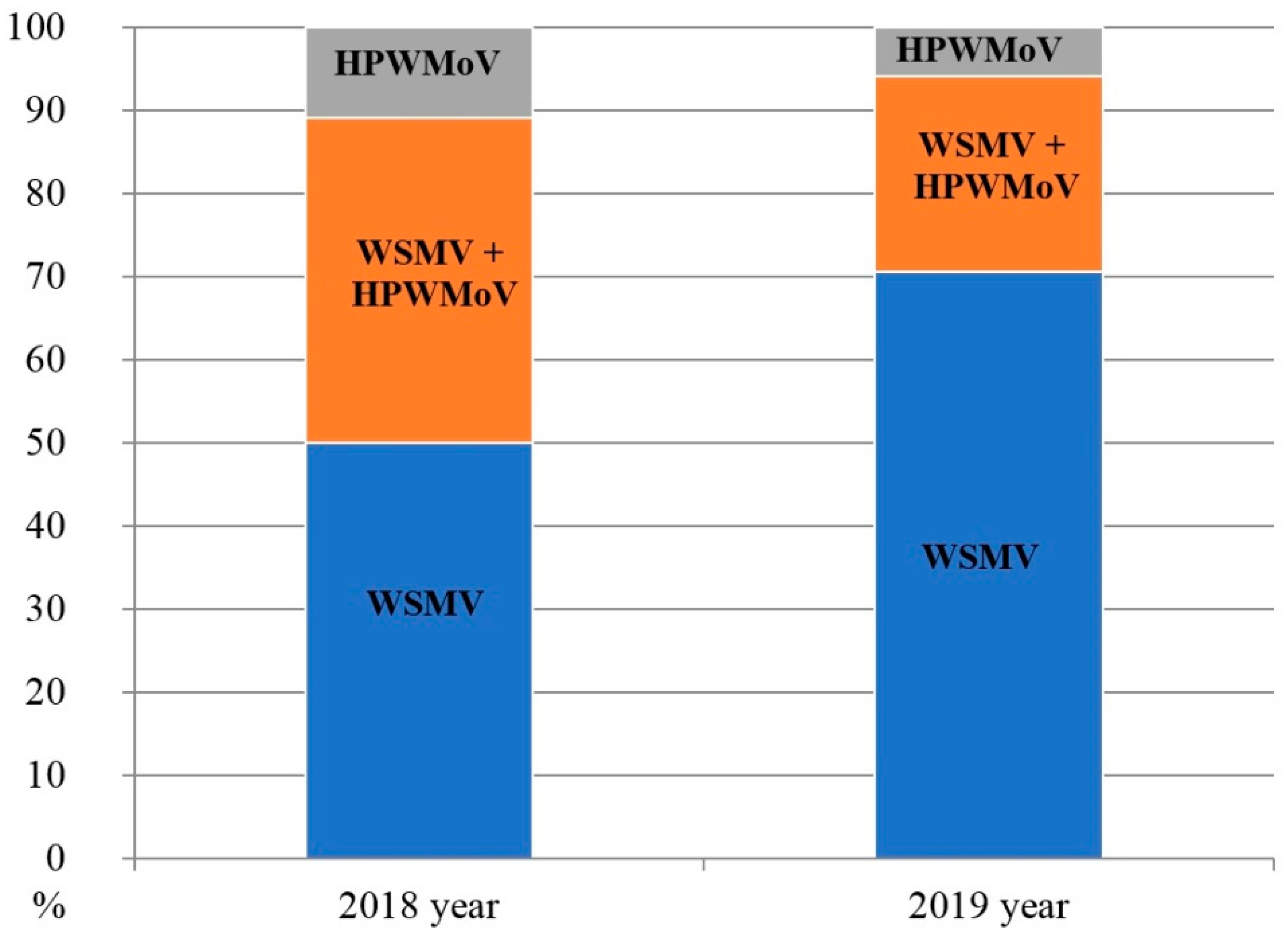
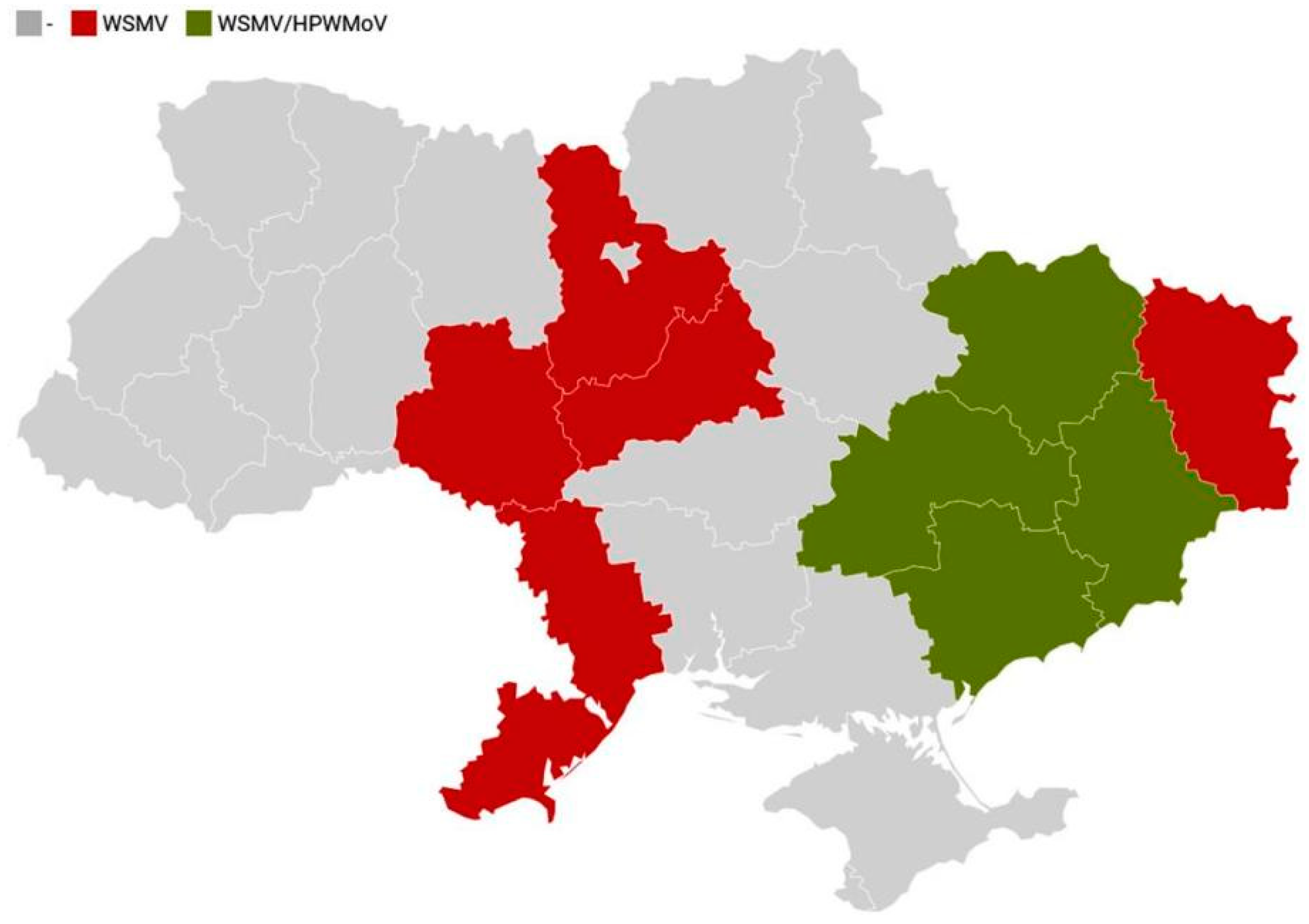
3.1. Surveys and Virus Detection by DAS-ELISA
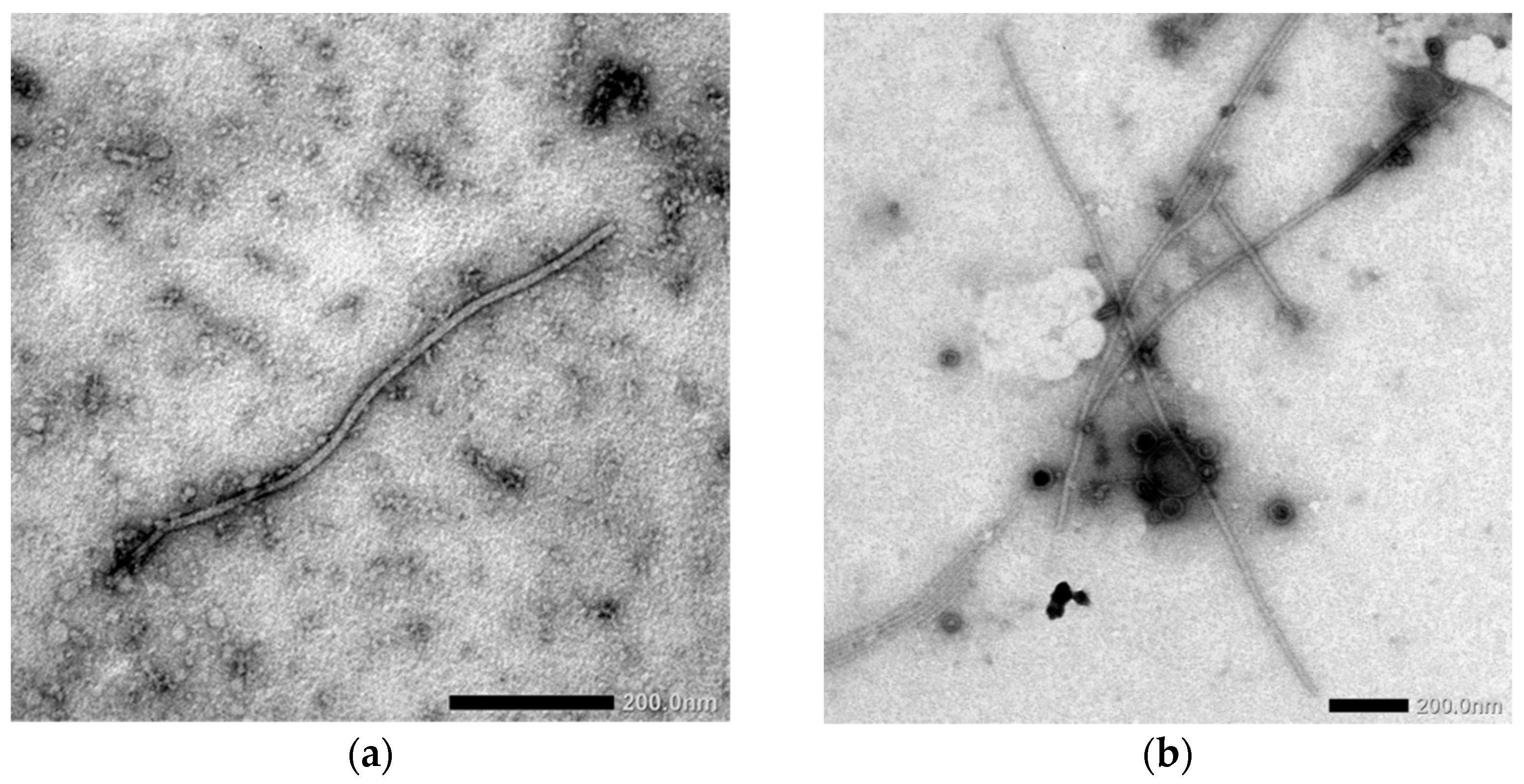
3.2. Direct Virus Indication Using Electron Microscopy
3.3. Virus Detection by RT-PCR in ELISA-Positive Samples
3.4. Phylogenetic Relationships and Sequence Similarity
4. Discussion
4.1. Occurrence of WSMV and HPWMoV
4.2. Patterns of Mono- and Mixed Infection of Winter Wheat with WSMV and HPWMoV
4.3. Susceptibility of Different Varieties of Winter Wheat to WSMV and HPWMoV Infections
4.4. Phylogenetic Analysis of WSMV and HPWMoV Isolates from Ukraine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabenstein, F.; Seifers, D.L.; Schubert, J.; French, R.; Stenger, D.C. Phylogenetic relationships, strain diversity and biogeography of tritimoviruses. J. Gen. Virol. 2002, 83, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, G.I.; Gibbs, M.J.; Gibbs, A.J.; Jones, R.A.C. Wheat streak mosaic virus in Australia: Relationship to isolates from the Pacific Northwest of the USA and its dispersion via seed transmission. Plant Dis. 2007, 91, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbanaviciene, L.; Šneideris, D.; Žizyte, M. Wheat streak mosaic virus detected in winter wheat in Lithuania. Zemdirb.-Agric. 2015, 102, 111–114. [Google Scholar] [CrossRef]
- Bennypaul, H.; Abdullahi, I.; Harding, M.; Neeser, C. First detection of Wheat streak mosaic virus in two perennial weed species, Agropyron cristatum (L.) Gaertn. and Hordeum jubatum subsp. intermedium Bowden, in Canada. Plant Dis. 2019, 103, 1441. [Google Scholar] [CrossRef]
- Rabenstein, F.; Stenger, D.C.; French, R. Genus Tritimovirus. In Viruses and virus diseases of Poaceae (Gramineae); Lapierre, H., Signoret, P.A., Eds.; INRA: Paris, France, 2004; pp. 398–402. [Google Scholar]
- Choi, I.R.; Horken, K.M.; Stenger, D.C.; French, R. Mapping of the P1 proteinase cleavage site in the polyprotein of Wheat streak mosaic virus (genus Tritimovirus). J. Gen. Virol. 2002, 83, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Stenger, D.C.; Hein, G.L.; French, R. Nested deletion analysis of Wheat streak mosaic virus HC-Pro: Mapping of domains affecting polyprotein processing and eriophyid mite transmission. Virology 2006, 350, 465–474. [Google Scholar] [CrossRef] [Green Version]
- CABI/EPPO. Wheat Streak Mosaic Virus; Distribution Maps of Plant Diseases, No. 443; CAB International: Wallingford, UK, 2005. [Google Scholar]
- Ellis, M.H.; Rebetzke, G.J.; Kelman, W.M.; Moore, C.S.; Hyles, J.E. Detection of Wheat streak mosaic virus in four pasture grass species in Australia. Plant Pathol. 2004, 53, 239. [Google Scholar] [CrossRef]
- Singh, K.; Wegulo, S.N.; Skoracka, A.; Kundu, J.K. Wheat streak mosaic virus: A century old virus with rising importance worldwide. Mol. Plant Pathol. 2018, 19, 2193–2206. [Google Scholar] [CrossRef] [Green Version]
- Divis, L.A.; Graybosch, R.A.; Peterson, C.J.; Baenziger, P.S.; Hein, G.L.; Beecher, B.B.; Martin, T.J. Agronomic and quality effects in winter wheat of a gene conditioning resistance to wheat streak mosaic virus. Euphytica 2006, 152, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Miller, Z.; Menalled, F.; Ito, D.; Moffet, M.; Burrows, M. Impacts of crop variety and time of inoculation on the susceptibility and tolerance of winter wheat to Wheat streak mosaic virus. Plant Dis. 2014, 98, 1060–1065. [Google Scholar] [CrossRef] [Green Version]
- Olejnyk, A.N. Wheat Streak in Ukraine. Ph.D. Dissertation, 1968; 15s. (In Russian). [Google Scholar]
- Shevchenko, G.P.; Helman, L.V.; Nedvyga, O.E.; Onyshchenko, A.M.; Porembska, N.B. Viral and Mycoplasma Diseases of Field Crops. K.: Urogaj. 1995. 304s. Available online: https://vdoc.pub/documents/-2c1k6ogj62hg (accessed on 1 April 2022). (In Ukrainian).
- Mischenko, L.T.; Bojko, A.L. Wheat streak mosaic virus in Ukraine. Rep. Natl. Acad. Sci. Ukr. 2004, 2, 176–180. (In Ukrainian) [Google Scholar]
- Mischenko, L.T. Viral Disease on Winter Wheat. K.: Fitosociocentr. 2009. 352s. Available online: https://www.twirpx.com/file/2565923/ (accessed on 1 April 2022). (In Ukrainian).
- McMechan, A.J. Over-Summering Ecology of the Wheat Curl Mite (Aceria tosichella Keifer). Ph.D. Dissertation, University of Nebraska, Lincoln, NE, USA, 2016. [Google Scholar]
- Wosula, E.N.; Tatineni, S.; Wegulo, S.N.; Hein, G.L. Effect of Temperature on Wheat Streak Mosaic Disease Development in Winter Wheat. Plant Dis. 2017, 101, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velandia, M.; Rejesus, R.M.; Jones, D.C.; Price, J.A.; Workneh, F.; Rush, C.M. Economic impact of Wheat streak mosaic virus in the Texas High Plains. Crop Prot. 2010, 29, 699–703. [Google Scholar] [CrossRef]
- French, R.; Stenger, D.C. Evolution of wheat streak mosaic virus: Dynamics of population growth within plants may explain limited variation. Annu. Rev. Phytopathol. 2003, 41, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Hadi, B.A.R.; Langhm, M.A.C.; Osborne, L.; Tilmon, K.J. Wheat streak mosaic virus on wheat: Biology and management. J. Integr. Pest Manag. 2011, 2, J1–J5. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-Hofman, C.; Wegulo, S.N.; Tatineni, S.; Hein, G.L. Impact of Wheat streak mosaic virus and Triticum mosaic virus coinfection of wheat on transmission rates by wheat curl mites. Plant Dis. 2015, 99, 1170–1174. [Google Scholar] [CrossRef] [Green Version]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [Green Version]
- Navia, D.; de Mendonca, R.S.; Skoracka, A.; Szydlo, W.; Knihinicki, D.; Hein, G.L.; da Silva Pereira, P.R.V.; Truol, G.; Lau, D. Wheat curl mite, Aceria tosichella, and transmitted viruses: An expanding pest complex affecting cereal crops. Exp. Appl. Acarol. 2013, 59, 95–143. [Google Scholar] [CrossRef] [Green Version]
- Coutts, B.A.; Banovic, M.; Kehoe, M.A.; Severtson, D.L.; Jones, R.A.C. Epidemiology of Wheat streak mosaic virus in wheat in a Mediterranean-type environment. Eur. J. Plant. Pathol. 2014, 140, 797–813. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Coulls, B.A.; Mackie, A.E.; Dwyer, G.I. Seed transmission of Wheat streak mosaic virus shown unequivocally in wheat. Plant Dis. 2005, 89, 1048–1050. [Google Scholar] [CrossRef] [Green Version]
- Lanoiselet, V.M.; Hind-Lanoiselet, T.L.; Murray, G.M. Studies on the seed transmission of Wheat streak mosaic virus. Australas. Plant Pathol. 2008, 37, 584–588. [Google Scholar] [CrossRef]
- Forster, R.L.; Seifers, D.L.; Strausbaugh, C.A.; Jensen, S.G.; Ball, E.M.; Harvey, T.L. Seed transmission of the High Plains virus in sweet corn. Plant Dis. 2001, 85, 696–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatineni, S.; Hein, G.L. High Plains wheat mosaic virus: An enigmatic disease of wheat and corn causing the High Plains disease. Mol. Plant Pathol. 2021, 22, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, E.-H.; Chu, Y.-M.; Shin, Y.-G.; Ahn, T.-Y. Development of PCR Diagnosis System for Plant Quarantine Seed-borne Wheat Streak Mosaic Virus. Korean J. Microbiol. 2013, 49, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Redila, C.D.; Prakash, V.; Nouri, S. Metagenomics Analysis of the Wheat Virome Identifies Novel Plant and Fungal-Associated Viral Sequences. Viruses 2021, 13, 2457. [Google Scholar] [CrossRef] [PubMed]
- Snihur, H.O.; Bosiuk, V.J.; Petrenko, S.M.; Shevchenko, O.V.; Polischuk, V.P. Spread of cereal viruses in some agriecosystems of Ukraine. Acta Physiol. Plant. 2009, 31 (Suppl. 1), S49. [Google Scholar]
- Snihur, H.; Petrenko, S.; Kot, T.; Shevchenko, O.; Polischuk, V. Widespread viral diseases endangering cereal crops in Ukraine. Mikrobiolohichnyi Zhurnal 2018, 80, 103–114. [Google Scholar] [CrossRef]
- Mishchenko, L.T.; Dunich, A.A.; Skrypkina, I.Y.; Kozub, N.O. Phylogenetic analysis of two Ukrainian isolates of Wheat streak mosaic virus. Biopolym. Cell 2019, 35, 64–77. [Google Scholar] [CrossRef]
- Snihur, H.; Pozhylov, I.; Budzanivska, I.; Shevchenko, O. First report of occurrence of High Plains wheat mosaic virus on different hosts in Ukraine. J. Plant Pathol. 2020, 102, 545–546. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.M. Characteristics of the microplate method of enzyme-linked immunosorbent assay for detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Kitajima, E. Electron Microscopy in Plant Virology: Past, Present and Future. Microsc. Microanal. 2004, 10, 212–213. [Google Scholar] [CrossRef]
- Richert-Pöggeler, K.R.; Franzke, K.; Hipp, K.; Kleespies, R.G. Electron Microscopy Methods for Virus Diagnosis and High Resolution Analysis of Viruses. Front. Microbiol. 2019, 9, 3255. [Google Scholar] [CrossRef] [PubMed]
- Kudela, O.; Kudelova, M.; Novakova, S.; Glasa, M. First report of Wheat streak mosaic virus in Slovakia. Plant Dis. 2008, 92, 1365. [Google Scholar] [CrossRef] [PubMed]
- Lebas, B.S.M.; Ochoa-Corona, F.M.; Elliott, D.R.; Tang, Z.; Alexander, B.J.R. Development of an RT-PCR for High Plains virus indexing scheme in New Zealand post-entry quarantine. Plant Dis. 2005, 89, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- Byamukama, E.; Tatineni, S.; Hein, G.L.; McMechan, J.A.; Wegulo, S.N. Incidence of Wheat streak mosaic virus, Triticum mosaic virus, and Wheat mosaic virus in wheat curl mites recovered from maturing winter wheat spikes. Plant Dis. 2016, 100, 318–323. [Google Scholar] [CrossRef] [Green Version]
- Gadiou, S.; Kúdela, O.; Ripl, J.; Rabenstein, F.; Kundu, J.K.; Glasa, M. An amino acid deletion in Wheat streak mosaic virus capsid protein distinguishes a homogeneous group of European isolates and facilitates their specific detection. Plant Dis. 2009, 93, 1209–1213. [Google Scholar] [CrossRef] [Green Version]
- Uyeda, I.; Masuta, C. Plant Virology Protocols: New Approaches to Detect Viruses and Host Responses. In Methods in Molecular Biology, 3rd ed.; Humana Press: New York, NY, USA, 2015; 292p. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Dayhoff, M.O.; Barker, W.C.; Hunt, L.T. Establishing homologies in protein sequences. Method Enzymol. 1983, 91, 524–545. [Google Scholar]
- Shevchenko, O.; Yasaka, R.; Tymchyshyn, O.; Shevchenko, T.; Ohshima, K. First evidence of the occurrence of Turnip mosaic virus in Ukraine and molecular characterization of its isolate. J. Phytopathol. 2018, 166, 429–437. [Google Scholar] [CrossRef]
- Valouzi, H.; Shahmohammadi, N.; Golnaraghi, A.; Moosavi, M.R.; Ohshima, K. Genetic diversity and evolutionary analyses of potyviruses infecting narcissus in Iran. J. Plant Pathol. 2022, 104, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Kawakubo, S.; Gao, F.; Li, S.; Tan, Z.; Huang, Y.K.; Adkar-Purushothama, C.R.; Gurikar, C.; Maneechoat, P.; Chiemsombat, P.; Aye, S.S.; et al. Genomic analysis of the brassica pathogen turnip mosaic potyvirus reveals its spread along the former trade routes of the Silk Road. Proc. Natl. Acad. Sci. USA 2021, 118, e2021221118. [Google Scholar] [CrossRef] [PubMed]
- Overmyer, L.M. Factors Influencing Wheat Curl Mite Aceria tosichella Keifer Dispersal. Diss. Stud. Res. Entomol. 2020, 65, 1–110. [Google Scholar]
- Karpicka-Ignatowska, K.; Laska, A.; Rector, B.G.; Skoracka, A.; Kuczyński, L. Temperature-dependent development and survival of an invasive genotype of wheat curl mite, Aceria Tosichella. Exp. Appl. Acarol. 2021, 83, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Burrows, M.; Franc, G.; Rush, C.; Blunt, T.; Ito, D.; Kinzer, K.; Olson, J.; O’Mara, J.; Price, J.; Tande, C.; et al. Occurrence of viruses in wheat in the Great Plains region, 2008. Plant Health Prog. 2009, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Harvey, T.L.; Seifers, D.L.; Martin, T.J.; Brown-Guedira, G.; Gill, B.S. Survival of wheat curl mites on different sources of resistance in wheat. Crop Sci. 1999, 39, 1887–1889. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Gene/Region | Forward Primer | Reverse Primer | Expected Product Length | Reference |
|---|---|---|---|---|---|
| WSMV | Coat protein | GAG AGC AAT ACT GCG TGT ACG | GCA TAA TGG CTC GAA GTG ATG | 750 bp | [39] |
| HPWMoV | Nucleoprotein | TTT ATG GCT CTT TGT ATT GG | TAT GTT TCC CCT CTT TGT G | 339 bp | [40] |
| TriMV | PIPO | CTT AAG CAC ATG TTA CAA TC | GTC CCT GAT AAC TAA TTC TA | 1200 bp | [41] |
| Region | District | Year of Sampling | BSMV * | WSMV * | HPWMoV * |
|---|---|---|---|---|---|
| Vinnytsia | Bershadskiy | 2017 | 0/5 | 5/5 | - |
| Dnipropetrovsk | Petropavlivskiy | 2017 | 0/15 | 15/15 | - |
| Pokrovskiy | 2017 | 0/10 | 10/10 | - | |
| Donetsk | Maryinskiy | 2017 | 0/5 | 0/5 | - |
| Kyiv | Kyiv-Svyatoshinskiy | 2017 | 0/10 | 5/10 | - |
| Luhansk | Novoaidarskiy | 2017 | 0/5 | 5/5 | - |
| Odessa | Ovidiopolskiy | 2017 | 0/10 | 5/10 | - |
| Kharkiv | Chuguyivskiy | 2017 | 0/10 | 5/10 | - |
| Vinnytsia | Bershadskiy | 2018 | 0/5 | 5/5 | 0/5 |
| Dnipropetrovsk | Mezhivskiy | 2018 | 0/5 | 5/5 ** | 5/5 ** |
| Pokrovskiy | 2018 | 0/5 | 0/5 | 5/5 | |
| Donetsk | Oleksandrivskiy | 2018 | 0/15 | 10/15 ** | 15/15 ** |
| Pokrovskiy | 2018 | 0/5 | 5/5 ** | 5/5 ** | |
| Zaporizhia | Vasylivskiy | 2018 | 0/10 | 10/10 ** | 10/10 ** |
| Kyiv | Kyiv-Svyatoshinskiy | 2018 | 0/10 | 10/10 | 0/10 |
| Kharkiv | Kupyanskiy | 2018 | 0/20 | 20/20 ** | 5/20 ** |
| Kharkivskiy | 2018 | 0/15 | 15/15 | 0/15 | |
| Vinnytsia | Bershadskiy | 2019 | 0/10 | 10/10 | 0/10 |
| Dnipropetrovsk | Mezhivskiy | 2019 | 0/30 | 25/30 ** | 20/30 ** |
| Pavlogradskiy | 2019 | 0/5 | 5/5 | 0/5 | |
| Synelnykivskiy | 2019 | 0/5 | 5/5 | 0/5 | |
| Zaporizhia | Bilmatskiy | 2019 | 0/5 | 5/5 | 0/5 |
| Rozivskiy | 2019 | 0/5 | 5/5 | 0/5 | |
| Kyiv | Kyiv-Svyatoshinskiy | 2019 | 0/5 | 5/5 | 0/5 |
| Kharkiv | Pechenizkiy | 2019 | 0/15 | 15/15 ** | 5/15 ** |
| Cherkassy | Umanskiy | 2019 | 0/5 | 5/5 | 0/5 |
| Total | 0/245 | 210/245 | 70/175 | ||
| Percentage of infected plants | 0 | 85.7 | 40 | ||
| Cultivar | Country of Origin of Cultivar | Virus Infection * | |
|---|---|---|---|
| WSMV | HPWMoV | ||
| Novosmuhlyanka | Ukraine | − | + |
| Garantiya Odesska | Ukraine | + | + |
| Donetska 48 | Ukraine | + | + |
| Dostatok | Ukraine | + | + |
| Podolyanka | Ukraine | + | + |
| Bohdana | Ukraine | + | − |
| Zymoyarka | Ukraine | + | − |
| Zolotokolosa | Ukraine | + | − |
| Misiya Odesska | Ukraine | + | − |
| Smuhlyanka | Ukraine | + | − |
| Emil | Germany | + | − |
| Cubus | Germany | + | − |
| Skagen | Germany | + | + |
| Isolate Name (GenBank Accession Number) | WSMV-UA-2017 (MK167470) | UA-WSMV-CP-ZP-18 (OM927716) | UA-WSMV-CP-ZP-21 (OM927719) | UA-WSMV-CP-DN-19 (OM927718) | UA-WSMV-CP-KH-18 (OM927717) | P-Ch-Vas-Ukr-2020 (MW072787.1) | Ukraine-Ep-18 (MH523357.1) | Ukraine-Mal-18 (MH523356.1) | Z-Vas-Ukr-2020 (MW072786.1) | DSMZ_PV-0356 (MZ202336.1) |
|---|---|---|---|---|---|---|---|---|---|---|
| WSMV-UA-2017 (MK167470) | - | 0.022 | 0.016 | 0.011 | 0.021 | 0.021 | 0.142 | 0.074 | 0.016 | 0.016 |
| UA-WSMV-CP-ZP-18 (OM927716) | 0.026 | - | 0.016 | 0.011 | 0.011 | 0.017 | 0.135 | 0.064 | 0.005 | 0.005 |
| UA-WSMV-CP-ZP-21 (OM927719) | 0.017 | 0.020 | - | 0.005 | 0.016 | 0.021 | 0.124 | 0.068 | 0.011 | 0.011 |
| UA-WSMV-CP-DN-19 (OM927718) | 0.013 | 0.016 | 0.004 | - | 0.011 | 0.016 | 0.130 | 0.062 | 0.005 | 0.005 |
| UA-WSMV-CP-KH-18 (OM927717) | 0.023 | 0.022 | 0.013 | 0.010 | - | 0.016 | 0.130 | 0.062 | 0.005 | 0.005 |
| P-Ch-Vas-Ukr-2020 (MW072787.1) | 0.027 | 0.026 | 0.017 | 0.013 | 0.019 | - | 0.137 | 0.068 | 0.011 | 0.011 |
| Ukraine-Ep-18 (MH523357.1) | 0.100 | 0.102 | 0.087 | 0.087 | 0.093 | 0.098 | - | 0.062 | 0.124 | 0.124 |
| Ukraine-Mal-18 (MH523356.1) | 0.064 | 0.063 | 0.053 | 0.049 | 0.055 | 0.060 | 0.039 | - | 0.056 | 0.056 |
| Z-Vas-Ukr-2020 (MW072786.1) | 0.021 | 0.020 | 0.012 | 0.008 | 0.013 | 0.017 | 0.087 | 0.049 | - | 0.000 |
| DSMZ_PV-0356 (MZ202336.1) | 0.021 | 0.020 | 0.012 | 0.008 | 0.013 | 0.017 | 0.087 | 0.045 | 0.012 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozhylov, I.; Snihur, H.; Shevchenko, T.; Budzanivska, I.; Liu, W.; Wang, X.; Shevchenko, O. Occurrence and Characterization of Wheat Streak Mosaic Virus Found in Mono- and Mixed Infection with High Plains Wheat Mosaic Virus in Winter Wheat in Ukraine. Viruses 2022, 14, 1220. https://doi.org/10.3390/v14061220
Pozhylov I, Snihur H, Shevchenko T, Budzanivska I, Liu W, Wang X, Shevchenko O. Occurrence and Characterization of Wheat Streak Mosaic Virus Found in Mono- and Mixed Infection with High Plains Wheat Mosaic Virus in Winter Wheat in Ukraine. Viruses. 2022; 14(6):1220. https://doi.org/10.3390/v14061220
Chicago/Turabian StylePozhylov, Illia, Halyna Snihur, Tetiana Shevchenko, Irena Budzanivska, Wenwen Liu, Xifeng Wang, and Oleksiy Shevchenko. 2022. "Occurrence and Characterization of Wheat Streak Mosaic Virus Found in Mono- and Mixed Infection with High Plains Wheat Mosaic Virus in Winter Wheat in Ukraine" Viruses 14, no. 6: 1220. https://doi.org/10.3390/v14061220