
Bovine Astrovirus—A Comprehensive Review


Abstract
:1. Introduction
2. Discovery and Epidemiology

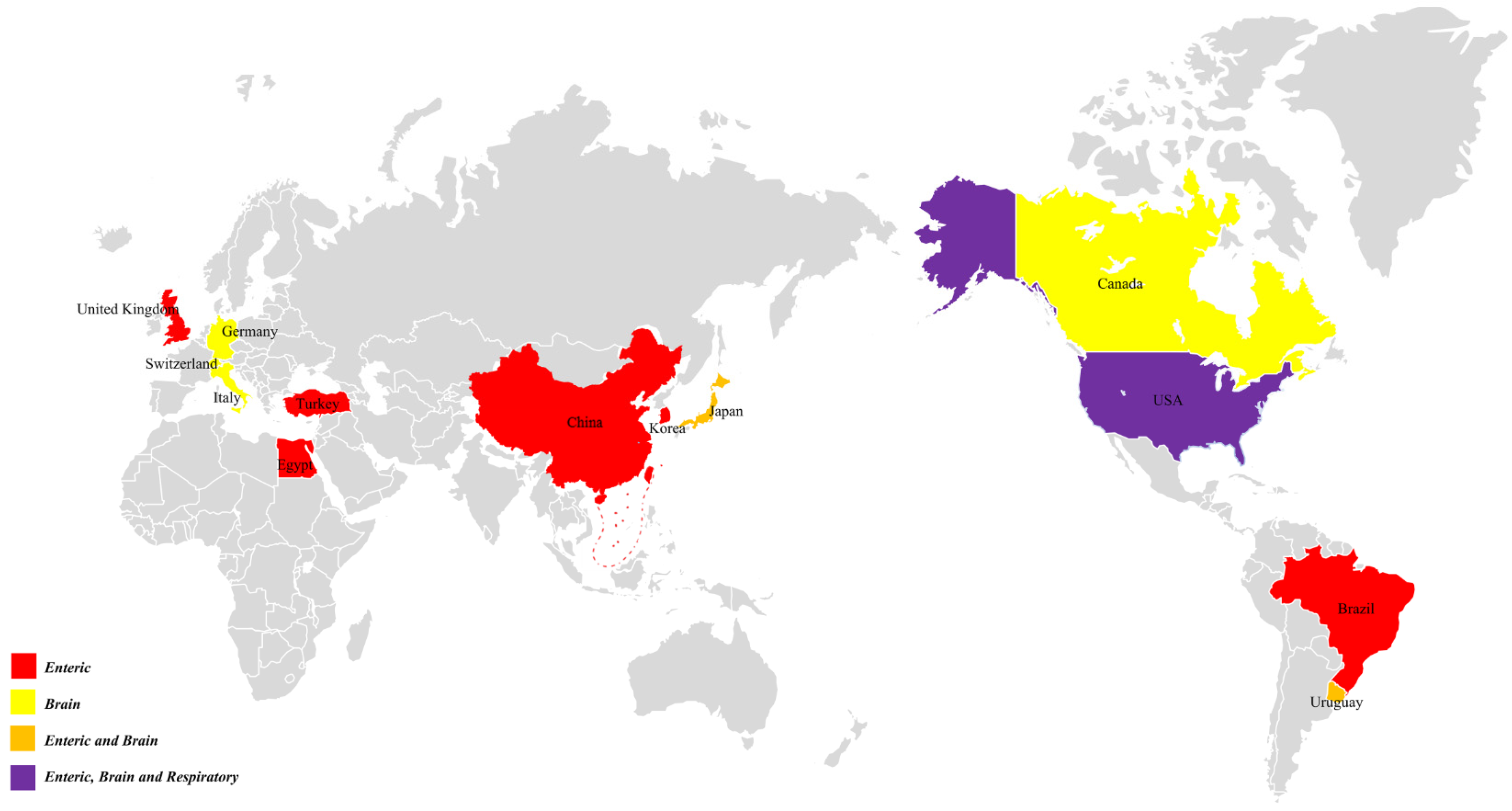{kind=link}
{kind=link}
{kind=link}
| Sample Type | Year | Country | Positive Rate | References |
|---|---|---|---|---|
| blood samples | 1978 | United Kingdom | 27.12% (16/59) | [9] |
| fecal samples | 1985 | USA | 2.64% (28/1060) | [10] |
| 2011 | China | 2.39% (5/209) | [14] | |
| 2014 | Korea | 7.83% (9/115) | [6] | |
| 2015 | Japan | 10.27% (15/146) | [15] | |
| United Kingdom | 65.19% (88/135) | [8] | ||
| China | 43.6% (92/211) | [16] | ||
| Brazil | 14.34% (39/272) | [4] | ||
| 2017 | Egypt | 32% (8/25) | [5] | |
| 2018 | Turkey | 3.15% (4/127) | [17] | |
| 2019 | Uruguay | 25.6% (128/500) | [18] | |
| 2021 | China | 46.34% (76/164) | [11] | |
| brain tissue samples | 2013 | USA | 12.5% (4/32) | [19] |
| 2014 | Switzerland | 22.73% (5/22) | [2] | |
| Germany | - | [20] | ||
| 2016 | Switzerland | 85.71% (12/14) | [26] | |
| Switzerland | 34.02% (33/97) | [7] | ||
| Canada | 44.44% (4/9) | [21,22] | ||
| Japan | 0.67% (1/150) | [23] | ||
| 2017 | Switzerland | 0.11% (2/1816) | [12] | |
| 2018 | Uruguay | 2.7% (1/37) | [24] | |
| 2019 | Italy | 0.36% (1/280) | [25] | |
| nasal swab samples | 2015 | USA | 8% (4/50) | [27] |
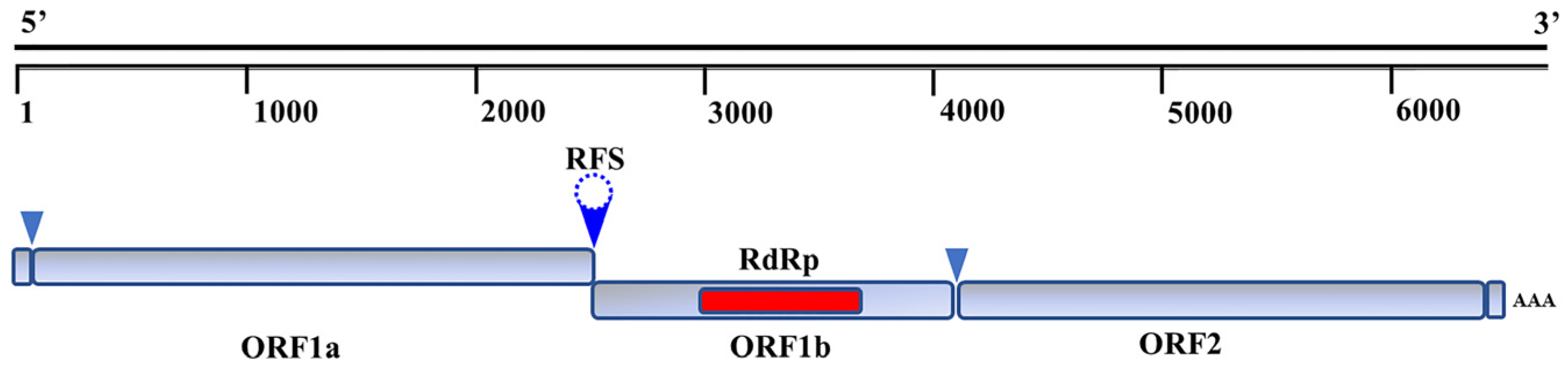
3. Genomic Characteristics
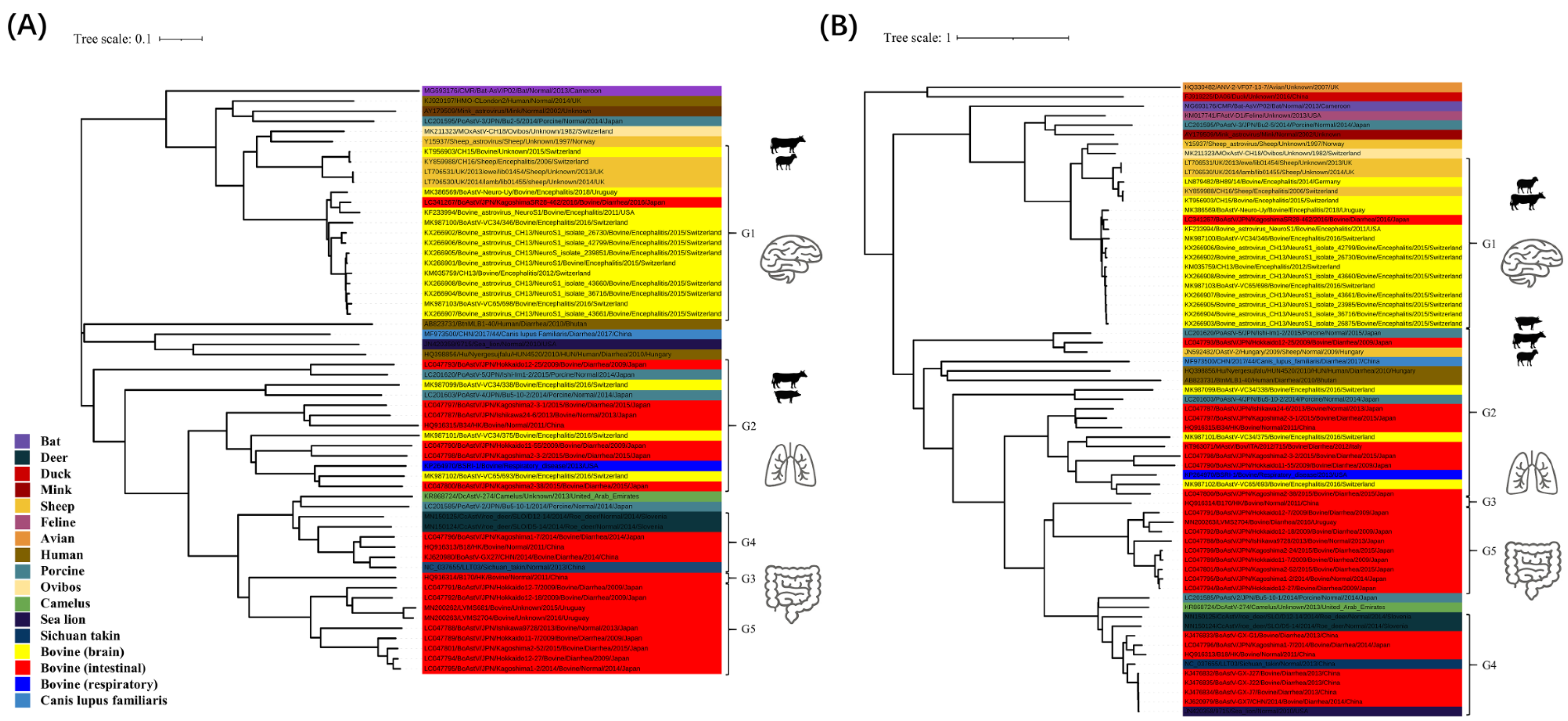
4. Phylogenetics and Evolution
5. Detection Methods
6. Coinfection of BoAstV with Typical Bovine Enteric, Neurotropic and Respiratory Viruses
7. Pathogenesis of BoAstV
8. Cross Species Transmission of BoAstV
9. Conclusions and Future Prospects for BoAstV
9.1. Establishment of a BoAstV Culture System
9.2. Potential Tropism of Other Organs
9.3. Pathogenic Significance of BoAstV
9.4. Exploration and Future Direction of Astrovirus Infection Spectrum
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouzalas, I.G.; Wuthrich, D.; Walland, J.; Drogemuller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffarena, R.D.; Casaux, M.L.; Schild, C.O.; Fraga, M.; Castells, M.; Colina, R.; Maya, L.; Corbellini, L.G.; Riet-Correa, F.; Giannitti, F. Causes of neonatal calf diarrhea and mortality in pasture-based dairy herds in Uruguay: A farm-matched case-control study. Braz. J. Microbiol. 2021, 52, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Candido, M.; Alencar, A.L.; Almeida-Queiroz, S.R.; Buzinaro Mda, G.; Munin, F.S.; de Godoy, S.H.; Livonesi, M.C.; Fernandes, A.M.; de Sousa, R.L. Molecular detection and phylogenetic analysis of bovine astrovirus in Brazil. Arch. Virol. 2015, 160, 1519–1525. [Google Scholar] [CrossRef]
- Mohamed, F.F.; Mansour, S.M.G.; El-Araby, I.E.; Mor, S.K.; Goyal, S.M. Molecular detection of enteric viruses from diarrheic calves in Egypt. Arch. Virol. 2017, 162, 129–137. [Google Scholar] [CrossRef]
- Oem, J.K.; An, D.J. Phylogenetic analysis of bovine astrovirus in Korean cattle. Virus Genes 2014, 48, 372–375. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and Pathological Phenotype of Bovine Astrovirus CH13/NeuroS1 Infection in Neurologically-Diseased Cattle: Towards Assessment of Causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef]
- Sharp, C.P.; Gregory, W.F.; Mason, C.; deC Bronsvoort, B.M.; Beard, P.M. High prevalence and diversity of bovine astroviruses in the faeces of healthy and diarrhoeic calves in South West Scotland. Vet. Microbiol. 2015, 178, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Woode, G.N.; Bridger, J.C. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J. Med. Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Woode, G.N.; Gourley, N.E.; Pohlenz, J.F.; Liebler, E.M.; Mathews, S.L.; Hutchinson, M.P. Serotypes of bovine astrovirus. J. Clin. Microbiol. 1985, 22, 668–670. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Qi, M.; Jiang, C.; Peng, Y.; Peng, Q.; Chen, Y.; Hu, C.; Chen, J.; Chen, X.; Chen, H.; et al. Prevalence of bovine astroviruses and their genotypes in sampled Chinese calves with and without diarrhoea. J. Gen. Virol. 2021, 102, 001640. [Google Scholar] [CrossRef]
- Truchet, L.; Walland, J.; Wuthrich, D.; Boujon, C.L.; Posthaus, H.; Bruggmann, R.; Schupbach-Regula, G.; Oevermann, A.; Seuberlich, T. Neuropathological survey reveals underestimation of the prevalence of neuroinfectious diseases in cattle in Switzerland. Vet. Microbiol. 2017, 208, 137–145. [Google Scholar] [CrossRef]
- Wuthrich, D.; Boujon, C.L.; Truchet, L.; Selimovic-Hamza, S.; Oevermann, A.; Bouzalas, I.G.; Bruggmann, R.; Seuberlich, T. Exploring the virome of cattle with non-suppurative encephalitis of unknown etiology by metagenomics. Virology 2016, 493, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Tse, H.; Chan, W.M.; Tsoi, H.W.; Fan, R.Y.Y.; Lau, C.C.Y.; Lau, S.K.P.; Woo, P.C.Y.; Yuen, K.Y. Rediscovery and genomic characterization of bovine astroviruses. J. Gen. Virol. 2011, 92 Pt 8, 1888–1898. [Google Scholar] [CrossRef]
- Nagai, M.; Omatsu, T.; Aoki, H.; Otomaru, K.; Uto, T.; Koizumi, M.; Minami-Fukuda, F.; Takai, H.; Murakami, T.; Masuda, T.; et al. Full genome analysis of bovine astrovirus from fecal samples of cattle in Japan: Identification of possible interspecies transmission of bovine astrovirus. Arch. Virol. 2015, 160, 2491–2501. [Google Scholar] [CrossRef]
- Alfred, N.; Liu, H.; Li, M.L.; Hong, S.F.; Tang, H.B.; Wei, Z.Z.; Chen, Y.; Li, F.K.; Zhong, Y.Z.; Huang, W.J. Molecular epidemiology and phylogenetic analysis of diverse bovine astroviruses associated with diarrhea in cattle and water buffalo calves in China. J. Vet. Med. Sci. 2015, 77, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Isidan, H.; Turan, T.; Atasoy, M.O.; Sozdutmaz, I.; Irehan, B. Detection and first molecular characterisation of three picornaviruses from diarrhoeic calves in Turkey. Acta Vet. Hung. 2019, 67, 463–476. [Google Scholar] [CrossRef]
- Castells, M.; Bertoni, E.; Caffarena, R.D.; Casaux, M.L.; Schild, C.; Victoria, M.; Riet-Correa, F.; Giannitti, F.; Parreno, V.; Colina, R. Bovine Astrovirus Surveillance in Uruguay Reveals High Detection Rate of a Novel Mamastrovirus Species. Viruses 2019, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Hoper, D.; Beer, M.; Hoffmann, B. Detection of a Novel Bovine Astrovirus in a Cow with Encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Sanchez, S.; Philibert, H.; Clark, E.G.; Seuberlich, T. Bovine astrovirus infection in feedlot cattle with neurological disease in western Canada. Can. Vet. J. 2017, 58, 601–603. [Google Scholar]
- Spinato, M.T.; Vince, A.; Cai, H.; Ojkic, D. Identification of bovine astrovirus in cases of bovine non-suppurative encephalitis in eastern Canada. Can. Vet. J. 2017, 58, 607–609. [Google Scholar]
- Hirashima, Y.; Okada, D.; Shibata, S.; Yoshida, S.; Fujisono, S.; Omatsu, T.; Mizutani, T.; Nagai, M. Whole genome analysis of a novel neurotropic bovine astrovirus detected in a Japanese black steer with non-suppurative encephalomyelitis in Japan. Arch. Virol. 2018, 163, 2805–2810. [Google Scholar] [CrossRef]
- Giannitti, F.; Caffarena, R.D.; Pesavento, P.; Uzal, F.A.; Maya, L.; Fraga, M.; Colina, R.; Castells, M. The First Case of Bovine Astrovirus-Associated Encephalitis in the Southern Hemisphere (Uruguay), Uncovers Evidence of Viral Introduction to the Americas from Europe. Front. Microbiol. 2019, 10, 1240. [Google Scholar] [CrossRef] [Green Version]
- Zaccaria, G.; Lorusso, A.; Hierweger, M.M.; Malatesta, D.; Defourny, S.V.; Ruggeri, F.; Camma, C.; Ricci, P.; Domenico, M.D.; Rinaldi, A.; et al. Detection of Astrovirus in a Cow with Neurological Signs by Nanopore Technology, Italy. Viruses 2020, 12, 530. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Bouzalas, I.G.; Vandevelde, M.; Oevermann, A.; Seuberlich, T. Detection of Astrovirus in Historical Cases of European Sporadic Bovine Encephalitis, Switzerland 1958–1976. Front. Vet. Sci. 2016, 3, 91. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.F.; Kondov, N.O.; Deng, X.; Van Eenennaam, A.; Neibergs, H.L.; Delwart, E. A metagenomics and case-control study to identify viruses associated with bovine respiratory disease. J. Virol. 2015, 89, 5340–5349. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, R.L.T.; Bogdanoff, W.A.; Espinoza, M.A.; López, S.; DuBois, R.M.; Arias, C.F. Isolation of Neutralizing Monoclonal Antibodies to Human Astrovirus and Characterization of Virus Variants That Escape Neutralization. J. Virol. 2019, 93, e01465-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, T. Novel human astroviruses: Challenges for developing countries. VirusDisease 2014, 25, 208–214. [Google Scholar] [CrossRef]
- Shi, Z.H.; Xu, Z.X.; Lan, Y.L.; Zhang, B.; Meng, H.L.; Wang, Y.Z.; Jin, L.; Wang, W.J. Establishment and application of a multiple PCR for detection of BNoV, BoAstV and BToV. Chin. J. Vet. Sci. 2019, 40, 1924–1927. [Google Scholar]
- Luthi, R.; Boujon, C.L.; Kauer, R.; Koch, M.C.; Bouzalas, I.G.; Seuberlich, T. Accurate and precise real-time RT-PCR assays for the identification of astrovirus associated encephalitis in cattle. Sci. Rep. 2018, 8, 9215. [Google Scholar] [CrossRef] [Green Version]
- Boujon, C.L.; Selimovic-Hamza, S.; Bouzalas, I.; Seuberlich, T. Development and validation of an immunohistochemistry procedure for the detection of a neurotropic bovine astrovirus. J. Virol. Methods 2017, 239, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Roach, S.N.; Langlois, R.A. Intra- and Cross-Species Transmission of Astroviruses. Viruses 2021, 13, 1127. [Google Scholar] [CrossRef]
- Mansour, S.M.G.; Mohamed, F.F.; Abdallah, F.M.; El-Araby, I.E.; Mor, S.K.; Goyal, S.M. Genomic features of first bovine astrovirus detected in Egypt. VirusDisease 2021, 32, 601–607. [Google Scholar] [CrossRef]
- Hierweger, M.M.; Werder, S.; Seuberlich, T. Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle. Int. J. Mol. Sci. 2020, 21, 498. [Google Scholar] [CrossRef] [Green Version]
- Janowski, A.B. Beyond the Gastrointestinal Tract: The Emerging and Diverse Tissue Tropisms of Astroviruses. Viruses 2021, 13, 732. [Google Scholar] [CrossRef]
- Woode, G.N.; Pohlenz, J.F.; Gourley, N.E.; Fagerland, J.A. Astrovirus and Breda virus infections of dome cell epithelium of bovine ileum. J. Clin. Microbiol. 1984, 19, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Koci, M.D.; Moser, L.A.; Kelley, L.A.; Larsen, D.; Brown, C.C.; Schultz-Cherry, S. Astrovirus induces diarrhea in the absence of inflammation and cell death. J. Virol. 2003, 77, 11798–11808. [Google Scholar] [CrossRef] [Green Version]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Blikslager, A.; Reddivari, M.; Heath, R.J.; Koci, M.D.; et al. Oral Administration of Astrovirus Capsid Protein Is Sufficient to Induce Acute Diarrhea In Vivo. mBio 2016, 7, e01494-16. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, D.L.; Saif, Y.M. Astrovirus: A cause of an enteric disease in turkey poults. Avian Dis. 1986, 30, 8. [Google Scholar] [CrossRef]
- Cortez, V.; Boyd, D.F.; Crawford, J.C.; Sharp, B.; Livingston, B.; Rowe, H.M.; Davis, A.; Alsallaq, R.; Robinson, C.G.; Vogel, P.; et al. Astrovirus infects actively secreting goblet cells and alters the gut mucus barrier. Nat. Commun. 2020, 11, 2097. [Google Scholar] [CrossRef]
- Wildi, N.; Seuberlich, T. Neurotropic Astroviruses in Animals. Viruses 2021, 13, 1201. [Google Scholar] [CrossRef]
- Guan, T.P.; Teng, J.L.L.; Yeong, K.Y.; You, Z.Q.; Liu, H.; Wong, S.S.Y.; Lau, S.K.P.; Woo, P.C.Y. Metagenomic analysis of Sichuan takin fecal sample viromes reveals novel enterovirus and astrovirus. Virology 2018, 521, 77–91. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Hoper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Rawal, G.; Ferreyra, F.M.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Mueller, A.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Detection and Cellular Tropism of Porcine Astrovirus Type 3 on Breeding Farms. Viruses 2019, 11, 1051. [Google Scholar] [CrossRef] [Green Version]
- Kauer, R.V.; Koch, M.C.; Schonecker, L.; Becker, J.; Holwerda, M.; Glaus, A.N.; Hierweger, M.M.; Werder, S.; Dijkman, R.; Meylan, M.; et al. Fecal Shedding of Bovine Astrovirus CH13/NeuroS1 in Veal Calves. J. Clin. Microbiol. 2020, 58, e01964-19. [Google Scholar] [CrossRef]
- Lukashov, V.V.; Goudsmit, J. Evolutionary relationships among Astroviridae. J. Gen. Virol. 2002, 83 Pt 6, 1397–1405. [Google Scholar] [CrossRef]
- Mendenhall, I.H.; Smith, G.J.; Vijaykrishna, D. Ecological Drivers of Virus Evolution: Astrovirus as a Case Study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [Green Version]
- Hata, A.; Kitajima, M.; Haramoto, E.; Lee, S.; Ihara, M.; Gerba, C.P.; Tanaka, H. Next-generation amplicon sequencing identifies genetically diverse human astroviruses, including recombinant strains, in environmental waters. Sci. Rep. 2018, 8, 11837. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, N.; Honce, R.; Schultz-Cherry, S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rodriguez, F.J.; Vieille, G.; Turin, L.; Yildiz, S.; Tapparel, C.; Kaiser, L. Fecal Components Modulate Human Astrovirus Infectivity in Cells and Reconstituted Intestinal Tissues. Msphere 2019, 4, e00568-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanoff, W.A.; Perez, E.I.; Lopez, T.; Arias, C.F.; DuBois, R.M. Structural Basis for Escape of Human Astrovirus from Antibody Neutralization: Broad Implications for Rational Vaccine Design. J. Virol. 2018, 92, e01546-17. [Google Scholar] [CrossRef] [Green Version]
- Acharya, M.; Arsi, K.; Donoghue, A.M.; Liyanage, R.; Rath, N.C. Production and characterization of avian crypt-villus enteroids and the effect of chemicals. BMC Vet. Res. 2020, 16, 179. [Google Scholar] [CrossRef]
- Zhang, Q.; Cao, Y.; Wang, J.; Fu, G.; Sun, M.; Zhang, L.; Meng, L.; Cui, G.; Huang, Y.; Hu, X.; et al. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings. Emerg. Microbes Infect. 2018, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Kolawole, A.O.; Mirabelli, C.; Hill, D.R.; Svoboda, S.A.; Janowski, A.B.; Passalacqua, K.D.; Rodriguez, B.N.; Dame, M.K.; Freiden, P.; Berger, R.P.; et al. Astrovirus replication in human intestinal enteroids reveals multi-cellular tropism and an intricate host innate immune landscape. PLoS Pathog. 2019, 15, e1008057. [Google Scholar] [CrossRef] [Green Version]
- Makimaa, H.; Ingle, H.; Baldridge, M.T. Enteric Viral Co-Infections: Pathogenesis and Perspective. Viruses 2020, 12, 904. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Yue, H. Molecular detection and genomic characteristics of bovine kobuvirus from dairy calves in China. Infect. Genet. Evol. 2019, 74, 103939. [Google Scholar] [CrossRef]
- Zhou, J.; Li, C.; Liu, X.; Chiu, M.C.; Zhao, X.; Wang, D.; Wei, Y.; Lee, A.; Zhang, A.J.; Chu, H.; et al. Infection of bat and human intestinal organoids by SARS-CoV-2. Nat. Med. 2020, 26, 1077–1083. [Google Scholar] [CrossRef]
- Fu, Y.; Pan, M.; Wang, X.; Xu, Y.; Xie, X.; Knowles, N.J.; Yang, H.; Zhang, D. Complete sequence of a duck astrovirus associated with fatal hepatitis in ducklings. J. Gen. Virol. 2009, 90 Pt 5, 1104–1108. [Google Scholar] [CrossRef]
- Imada, T.; Yamaguchi, S.; Mase, M.; Tsukamoto, K.; Kubo, M.; Morooka, A. Avian nephritis virus (ANV) as a new member of the family Astroviridae and construction of infectious ANV cDNA. J. Virol. 2000, 74, 8487–8493. [Google Scholar] [CrossRef] [Green Version]
- Wunderli, W.; Meerbach, A.; Gungor, T.; Berger, C.; Greiner, O.; Caduff, R.; Trkola, A.; Bossart, W.; Gerlach, D.; Schibler, M.; et al. Astrovirus infection in hospitalized infants with severe combined immunodeficiency after allogeneic hematopoietic stem cell transplantation. PLoS ONE 2011, 6, e27483. [Google Scholar] [CrossRef] [Green Version]
- Rawal, G.; Ferreyra, F.M.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Ecology of Porcine Astrovirus Type 3 in a Herd with Associated Neurologic Disease. Viruses 2020, 12, 992. [Google Scholar] [CrossRef]
- Bridger, J.C.; Hall, G.A.; Brown, J.F. Characterization of a calici-like virus (Newbury agent) found in association with astrovirus in bovine diarrhea. Infect. Immun. 1984, 43, 133–138. [Google Scholar] [CrossRef] [Green Version]



| Order | Year | Country | Sample Type | Co-Infection | References |
|---|---|---|---|---|---|
| 1 | 1978 | United Kingdom | stool samples/rectal swab | BEC, BRV | [9] |
| 2 | 2014 | Korea | stool samples/rectal swab | BcoV, BVDV, BKV, BRV | [6] |
| 3 | 2015 | China | stool samples/rectal swab | BtoV, BVDV, BEV, BcoV BRV | [16] |
| 4 | 2016 | Switzerland | brain tissue samples | BoHV-6 | [13] |
| 5 | 2017 | Egypt | stool samples/rectal swab | BnoV, BRV | [5] |
| 6 | 2021 | China | stool samples/rectal swab | BRV, BKV, BEV | [11] |
| 7 | 2016 | Switzerland | brain tissue samples | BoHV-6 | [13] |
| 8 | 2020 | Switzerland | brain tissue samples | PIV-5, BpyV-2, BoHV6 | [37] |
| 9 | 2015 | USA | nasal swabs samples | BadV-3, BRAV, BRBV, IDV | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Li, B.; Sun, D. Bovine Astrovirus—A Comprehensive Review. Viruses 2022, 14, 1217. https://doi.org/10.3390/v14061217
Zhu Q, Li B, Sun D. Bovine Astrovirus—A Comprehensive Review. Viruses. 2022; 14(6):1217. https://doi.org/10.3390/v14061217
Chicago/Turabian StyleZhu, Qinghe, Bin Li, and Dongbo Sun. 2022. "Bovine Astrovirus—A Comprehensive Review" Viruses 14, no. 6: 1217. https://doi.org/10.3390/v14061217