
Rotaviruses: From Pathogenesis to Disease Control—A Critical Review


Abstract
:1. Introduction
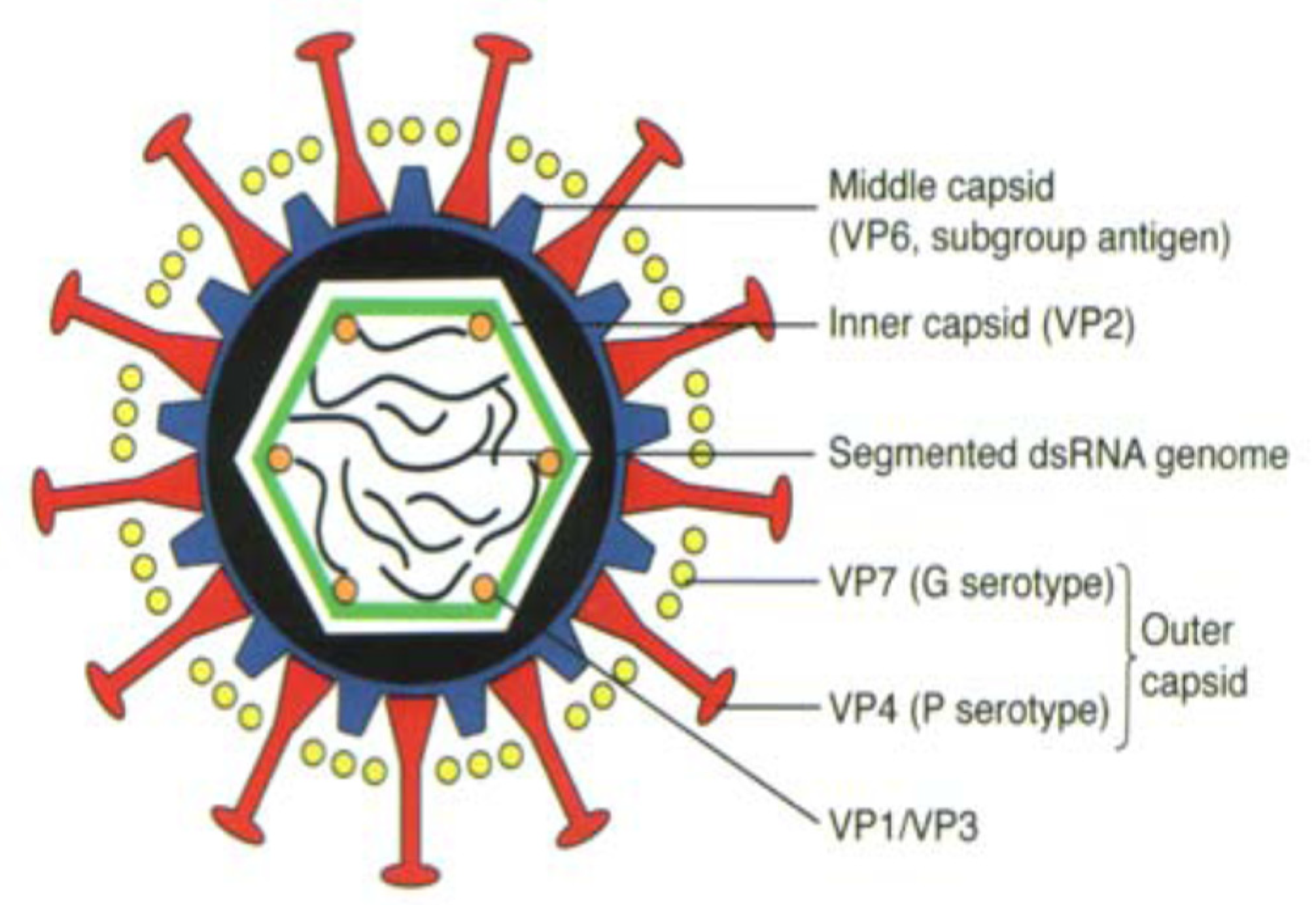
2. Etymology and Biology of Rotavirus
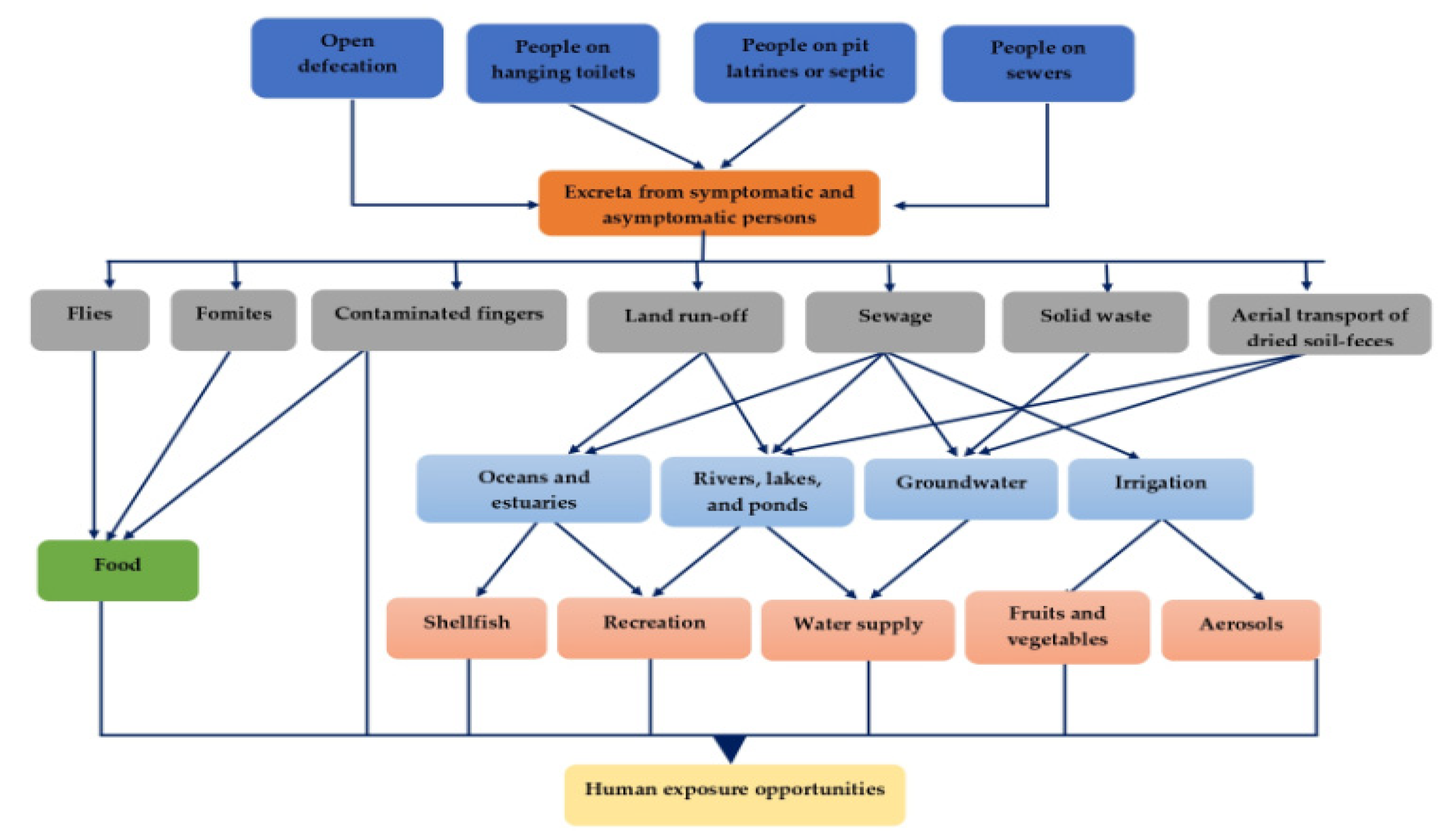
3. Exposure Pathways in Developing Countries
4. Pathogenesis of Rotavirus Infection
4.1. Viral Entry and Site of Primary Replication
4.2. Local Intestinal Infection and Disease Mechanisms
4.3. Systemic Infection
4.4. Host and Viral Factors Influencing Pathogenesis
4.5. Clinical Features
5. Immunity to Rotavirus
5.1. Innate Immune Response
5.2. Humoral Immunity
5.3. Cell-Mediated Immunity
6. Laboratory Diagnosis of Rotavirus Infection
7. Epidemiology and Molecular Diversity
7.1. Morbidity and Mortality in Children
7.2. Age and Sex Incidence Distribution
7.3. Seasonal Patterns of Infection
7.4. Nosocomially-Acquired Infection
8. Prevention and Control
8.1. Vaccination
Factors Influencing RV Vaccine Efficacy and Effectiveness in Poor Socioeconomic Settings
8.2. Nonvaccine Approaches
8.2.1. Good Hygiene
8.2.2. Breastfeeding
8.2.3. Probioses
8.2.4. Antiviral Drugs
9. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea among Children Younger Than 5 Years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef] [Green Version]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D.; World Health Organization–Coordinated Global Rotavirus Surveillance Network. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. 2016, 62, S96–S105. [Google Scholar] [CrossRef] [Green Version]
- Moraga, P.; GBD 2016 Causes of Death Collaborators. Global, regional, and national age-sex specific mortality for 264 causes of death, 1980–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1151–1210. [Google Scholar]
- World Health Organization. Summary of Key Characteristics of Currently WHO-Pre-Qualified Rotavirus Vaccines, Version 1.4 Dated 26 February 2019. Available online: https://www.who.int/publications/i/item/WHO-IVB-2021.03 (accessed on 15 July 2021).
- Kim, A.H.; Hogarty, M.P.; Harris, V.C.; Baldridge, M.T. The Complex Interactions between Rotavirus and the Gut Microbiota. Front. Cell Infect. Microbiol. 2021, 10, 586751. [Google Scholar] [CrossRef]
- Lee, B. Update on rotavirus vaccine underperformance in low- to middle-income countries and next-generation vaccines. Hum. Vaccines Immunother. 2021, 17, 1787–1802. [Google Scholar] [CrossRef]
- Saha, D.; Ota, M.O.C.; Pereira, P.; Buchy, P.; Badur, S. Rotavirus vaccines performance: Dynamic interdependence of host, pathogen and environment. Expert Rev. Vaccines 2021, 20, 945–957. [Google Scholar] [CrossRef]
- Adams, W.R.; Kraft, L.M. Epizootic diarrhea of infant mice: Indentification of the etiologic agent. Science 1963, 141, 359–360. [Google Scholar] [CrossRef]
- Bishop, R.F.; Davidson, G.P.; Holmes, I.H.; Ruck, B.J. Virus particles in epithelial cells of duodenal mucosa from children with viral gastroenteritis. Lancet 1973, 1, 1281–1283. [Google Scholar] [CrossRef]
- Bishop, R.F.; Davidson, G.P.; Holmes, I.H.; Ruck, B.J. Detection of a new virus by electron microscopy of faecal extracts from children with acute gastroenteritis. Lancet 1974, 1, 149–151. [Google Scholar] [CrossRef]
- Flewett, T.H.; Woode, G.N. The rotaviruses. Arch. Virol. 1978, 57, 1–23. [Google Scholar] [CrossRef]
- Estes, M.K. Rotaviruses and their replication. In Fields Virology, 3rd ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Chanock, R.M., Monath, T.P., Melnick, J.L., Eds.; Raven Press: New York, NY, USA, 1996; Volume 2, pp. 1625–1655. [Google Scholar]
- National Center for Immunization and Respiratory Diseases/CDC. Rotavirus Vaccination. Last Reviewed: 25 July 2018. Available online: https://www.cdc.gov/vaccines/vpd/rotavirus/index.html (accessed on 8 August 2020).
- World Health Organization Causes of Child Mortality (WHO 2021). Available online: https://www.who.int/gho/child_health/mortality/causes/en/ (accessed on 23 February 2021).
- Long, C.P.; McDonald, S.M. Rotavirus genome replication: Some assembly required. PLoS Pathog. 2017, 13, e1006242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estes, M.K.; Kapikian, A. Rotaviruses. In Fields Virology; Knipe, D., Griffin, D., Lamb, R., Martin, M., Roizman, B., Straus, S., Eds.; Wolters Kluwer Health; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1917–1975. [Google Scholar]
- Gómez-Rial, J.; Rivero-Calle, I.; Salas, A.; Martinón-Torres, F. Rotavirus and autoimmunity. J. Inf. 2020, 81, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Surendran, S. Review article: Rotavirus infection: Molecular changes and Pathophysiology. EXCLI J. 2008, 7, 154. [Google Scholar]
- Crawford, S.E.; Ramani, S.; Tate, J.E.; Parashar, U.D.; Svensson, L.; Hagbom, M.; Franco, M.A.; Greenberg, H.B.; O’Ryan, M.; Kang, G.; et al. Rotavirus infection. Nat. Rev. Dis. Primers 2017, 3, 17083. [Google Scholar] [CrossRef] [Green Version]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Steger, C.L.; Boudreaux, C.E.; LaConte, L.E.; Pease, J.B.; McDonald, S.M. Group a Rotavirus VP1 Polymerase and VP2 Core Shell Proteins: Intergenotypic Sequence Variation and In Vitro Functional Compatibility. J. Virol. 2019, 93, e01642-18. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Rotavirus Vaccines: WHO Position Paper—July 2021. Wkly. Epidemiol. Rec. 2021, 28, 301–320. Available online: http://www.who.int/wer (accessed on 16 September 2021).
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Bányai, K.; Brister, J.R. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [Green Version]
- Cunliffe, N.A.; Bresee, J.S.; Gentsch, J.R.; Glass, R.I.; Hart, C.A. The expanding diversity of rotaviruses. Lancet 2002, 359, 640–642. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, A.E.; Iturriza-Gómara, M.; Maes, P.; Patton, J.T.; et al. Full genome-based classification of rotaviruses reveals common origin between human Wa-like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef] [Green Version]
- RCWG. Newly Assigned Genotypes: List of Accepted Genotypes. 2019. Available online: https://rega.kuleuven.be/cev/viralmetagenomics/virus-classification/rcwg (accessed on 2 March 2021).
- World Health Organization (WHO). Vaccine-Preventable Diseases Surveillance Standards. Rotavirus. Last Updated: 5 September 2018. Available online: https://www.who.int/publications/i/item/surveillance-standards-for-vaccine-preventable-diseases-2nd-edition (accessed on 20 April 2020).
- Julian, T.R. Environmental transmission of diarrheal pathogens in low and middle income countries. Environ. Sci. Process. Impacts 2016, 18, 944–955. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Sustained Decrease in Laboratory Detection of Rotavirus after Implementation of Routine Vaccination—United States 2004–2014. CDC. 2015. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6413a1.htm (accessed on 16 September 2021).
- Burke, R.M.; Tate, J.E.; Barin, N.; Bock, C.; Bowen, M.D.; Chang, D.; Gautam, R.; Han, G.; Holguin, J.; Huynh, T.; et al. Three rotavirus outbreaks in the postvaccine era—California, 2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilde, J.; Van, R.; Pickering, L.; Eiden, J.; Yolken, R. Detection of rotaviruses in the day care environment by reverse transcriptase polymerase chain reaction. J. Infect. Dis. 1992, 166, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Meštrović, T. Rotavirus Transmission. News Medical.Net. Last Updated: 2 March 2021. Available online: https://www.newsmedical.net/health/Rotavirus-Transmission.aspx (accessed on 16 September 2021).
- Dennehy, P.H. Transmission of rotavirus and other enteric pathogens in the home. Pediatr. Infect. Dis. J. 2000, 19, S103–S105. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.; Weinstock, D.M.; Eagan, J.; Kiehn, T.; Armstrong, D.; Sepkowitz, K.A. Rotavirus outbreak on a pediatric oncology floor: Possible association with toys. Amer. J. Inf. Control. 2000, 28, 378–380. [Google Scholar] [CrossRef]
- Centers for Disease Prevention and Control. Epidemiology and Prevention of Vaccine-Preventable Diseases; Cortese, M.M., Haber, P., Eds.; National Center for Immunization and Respiratory Diseases: Atlanta, GA, USA, 2005; Chaper 19: Rotavirus. Available online: https://www.cdc.gov/vaccines/pubs/pinkbook/rota.html (accessed on 3 November 2020).
- Abad, F.X.; Pinto, R.M.; Bosch, A. Survival of enteric viruses on environmental fomites. Appl. Environ. Microbiol. 1994, 60, 3704–3710. [Google Scholar] [CrossRef] [Green Version]
- Gallimore, C.I.; Pipkin, C.; Shrimpton, H.; Green, A.D.; Pickford, Y.; McCartney, C. Detection of multiple enteric virus strains within a foodborne outbreak of gastroenteritis: An indication of the source of contamination. Epidemiol. Infect. 2005, 133, 41–47. [Google Scholar] [CrossRef]
- Quiroz-Santiago, C.; Vázquez-Salinas, C.; Natividad-Bonifacio, I.; Barrón-Romero, B.L.; Quiñones-Ramírez, E.I. Rotavirus G2P4 detection in fresh vegetables and oysters in Mexico City. J. Food Prot. 2014, 77, 1953–1959. [Google Scholar] [CrossRef]
- van Zyl, W.B.; Page, N.A.; Grabow, W.O.K.; Steele, A.D.; Taylor, M.B. Molecular Epidemiology of Group A Rotaviruses in Water Sources and Selected Raw Vegetables in Southern Africa. Appl. Environ. Microbiol. 2006, 72, 4554–4560. [Google Scholar] [CrossRef] [Green Version]
- NCID; National Institute of Infectious Diseases. An Outbreak of Group A Rotavirus Infections among Adults from Eating Meals Prepared at a Restaurant, April 2000—Shimane; Japan Ministry of Health and Welfare, Infectious Agents and Surveillance Report: Tokyo, Japan, 2000; Volume 27, p. 145.
- Le Guyader, F.S.; Le Saux, J.C.; Ambert-Balay, K.; Krol, J.; Serais, O.; Parnaudeau, S.; Giraudon, H.; Delmas, G.; Pommepuy, M.; Pothier, P. Aichi virus, norovirus, astrovirus, enterovirus, and rotavirus involved in clinical cases from a French oyster-related gastroenteritis outbreak. J. Clin. Microbiol. 2008, 46, 4011–4017. [Google Scholar] [CrossRef] [Green Version]
- Iritani, N.; Kaida, A.; Abe, N.; Kubo, H.; Sekiguchi, J.; Yamamoto, S.P.; Goto, K.; Tanaka, T.; Noda, M. Detection and genetic characterization of human enteric viruses in oyster-associated gastroenteritis outbreaks between 2001 and 2012 in Osaka City, Japan. J. Med. Virol. 2014, 86, 2019–2025. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Foodborne outbreak of Group A rotavirus gastroenteritis among college students—District of Columbia, March–April 2000. MMWR Morb. Mortal. Wkly. Rep. 2000, 49, 1131–1133. [Google Scholar]
- Mizukoshi, F.; Kuroda, M.; Tsukagoshi, H.; Sekizuka, T.; Funatogawa, K.; Morita, Y.; Noda, M.; Katayama, K.; Kimura, H. A food-borne outbreak of gastroenteritis due to genotype G1P[8] rotavirus among adolescents in Japan. Microbiol. Immunol. 2014, 58, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H.; Heiskanen, P.; Reid, I.; Buckle, K.A. Foodborne viral illness—Status in Australia. Int. J. Food Microbiol. 2000, 59, 127–136. [Google Scholar] [CrossRef]
- Mayr, C.; Strohe, G.; Contzen, M. Detection of rotavirus in food associated with a gastroenteritis outbreak in a mother and child sanatorium. Int. J. Food Microbiol. 2009, 135, 179–182. [Google Scholar] [CrossRef]
- Ansari, S.A.; Sattar, S.A.; Springthorpe, V.S.; Wells, G.A.; Tostowaryk, W. Rotavirus survival on human hands and transfer of infectious virus to animate and nonporous inanimate surfaces. J. Clin. Microbiol. 1988, 26, 1513–1518. [Google Scholar] [CrossRef] [Green Version]
- Gastañaduy, P.A.; Hall, A.J.; Parashar, U.D. Rotavirus. Foodborne Infect. Intox. 2013, 21, 303–311. [Google Scholar]
- U.S. EPA. Exposure Factors Handbook 2011 Edition (Final); EPA/600/R-09/052F; U.S. Environmental Protection Agency: Washington, DC, USA, 2011; p. 1436.
- Omatola, C.A.; Olusola, B.A.; Odaibo, G.N. Rotavirus infection among under five children presenting with gastroenteritis in Ibadan, Nigeria. Arch. Bas. Appl. Med. 2016, 3, 3–8. [Google Scholar]
- CDC (Centers for Disease Control and Prevention) (1998–2009). Foodborne Disease Outbreak Surveillance System (FDOSS), 1998–2009 Atlanta, Georgia. Available online: https://www.cdc.gov/fdoss/index.html (accessed on 16 September 2021).
- Tan, S.W.; Yap, K.L.; Lee, H.L. Mechanical Transport of Rotavirus by the Legs and Wings of Musca domestica (Diptera: Muscidae). J. Med. Entomol. 1997, 34, 527–531. [Google Scholar] [CrossRef]
- Issa, R. Musca domestica acts as transport vector hosts. Bull. Natl. Res. Cent. 2019, 43, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Collinet-Adler, S.; Babji, S.; Francis, M.; Kattula, D.; Premkumar, P.S.; Sarkar, R.; Mohan, V.R.; Ward, H.; Kang, G.; Balraj, V.; et al. Environmental Factors Associated with High Fly Densities and Diarrhea in Vellore, India. Appl. Environ. Microbiol. 2015, 81, 6053–6058. [Google Scholar] [CrossRef] [Green Version]
- Dóró, R.; Farkas, S.L.; Martella, V.; Bányai, K. Zoonotic transmission of rotavirus: Surveillance and control. Expert Rev. Anti. Infect. Ther. 2015, 13, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.; Dias, H.G.; Gonçalves, J.L.S.; Manchego, A.; Rosadio, R.; Pezo, D.; Santos, N. Genetic diversity and zoonotic potential of rotavirus A strains in the southern Andean highlands, Peru. Transbound. Emerg. Dis. 2019, 66, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.S.; Bhat, S.; Dar, P.S.; Sircar, S.; Dhama, K.; Singh, R.K. Evolving Rotaviruses, Interspecies Transmission and Zoonoses. Open Virol. J. 2020, 14, 1–6. [Google Scholar] [CrossRef]
- Omatola, C.A.; Olaniran, A.O. Epidemiological significance of the occurrence and persistence of rotaviruses in water and sewage: A critical review and proposal for routine microbiological monitoring. Environ. Sci. Processes Impacts 2022, 24, 380–399. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.R.; Davis, J.; Boehm, A.B. Mechanisms of post-supply contamination of drinking water in Bagamoyo, Tanzania. J. Water Health 2013, 11, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Yen, C.; Cortese, M.M. Rotaviruses. Principles and Practice of Pediatric Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1122–1125.e3. [Google Scholar]
- Bishop, R.F.; Masendycz, P.J.; Bugg, H.C. Epidemiological patterns of rotaviruses causing severe gastroenteritis in young children throughout Australia from 1993 to 1996. J. Clin. Microbiol. 2001, 39, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Levy, K.; Hubbard, A.E.; Elsenberg, J.N.S. Seasonality of rotavirus disease in the tropics: A systematic review and meta-analysis. Int. J. Epidemiol. 2009, 38, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Fragoso, M.; Kumar, A.; Murray, D.L. Rotavirus in nasopharyngeal secretions of children with upper respiratory tract infections. Diagn. Microbiol. Infect. Dis. 1986, 4, 87–88. [Google Scholar] [CrossRef]
- Zhen, B.J.; Chang, R.X.; Ma, G.Z.; Xie, J.M.; Liu, Q.; Liang, X.R.; Ng, M.H. Rotavirus infection of the oropharynx and respiratory tract in young children. J. Med. Virol. 1991, 34, 29–37. [Google Scholar] [CrossRef]
- Goldwater, P.; Chrystie, I.; Banatvala, J. Rotaviruses and the respiratory tract. Br. Med. J. 1979, 2, 1551. [Google Scholar] [CrossRef] [Green Version]
- Grimwood, K.; Lambert, S.B.; Milne, R.J. Rotavirus infections and vaccines: Burden of illness and potential impact of vaccination. Paediatr. Drugs 2010, 12, 235–256. [Google Scholar] [CrossRef] [PubMed]
- Ginn, O.; Rocha-Melogno, L.; Bivins, A.; Lowry, S.; Cardelino, M.; Nichols, D.; Tripathi, S.; Soria, F.; Andrade, M.; Bergin, M.; et al. Detection and quantification of enteric pathogens in aerosols near open wastewater canals in cities with poor sanitation. MedRxiv 2021, 14, 21251650. [Google Scholar] [CrossRef] [PubMed]
- Okaali, D.A.; Hofstra, N. Present and Future Human Emissions of Rotavirus and to Uganda’s Surface Waters. J. Environ. Quality. 2018, 47, 1130–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, A. Human enteric viruses in the water environment: A minireview. Int. Microbiol. 1998, 1, 191–196. [Google Scholar] [PubMed]
- Estes, M.K.; Greenberg, H.B. Rotaviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer Health/Lippincott Williams &Wilkins: Philadelphia, PA, USA, 2013; pp. 1347–1401. [Google Scholar]
- Sadiq, A.; Bostan, N.; Yinda, K.C.; Naseem, S.; Sattar, S. Rotavirus: Genetics, pathogenesis and vaccine advances. Rev. Med. Virol. 2018, 28, e2003. [Google Scholar] [CrossRef] [PubMed]
- Saxena, K.; Blutt, S.E.; Ettayebi, K.; Zeng, X.L.; Broughman, J.R.; Crawford, S.E.; Karandikar, U.C.; Sastri, N.P.; Conner, M.E.; Opekun, A.R.; et al. Human intestinal enteroids: A new model to study human rotavirus infection, host restriction, and pathophysiology. J. Virol. 2015, 90, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Jiang, X. Histo-blood group antigens: A common niche for norovirus and rotavirus. Expert Rev. Mol. Med. 2014, 16, e5. [Google Scholar] [CrossRef]
- Arias, C.F.; Silva-Ayala, D.; López, S. Rotavirus entry: A deep journey into the cell with several exits. J. Virol. 2015, 89, 890–893. [Google Scholar] [CrossRef] [Green Version]
- Ramani, S.; Hu, L.; Venkataram Prasad, B.V.; Estes, M.K. Diversity in rotavirus-host glycan interactions: A “sweet” spectrum. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Hagbom, M.; Svensson, L.; Nordgren, J. The Impact of Human Genetic Polymorphisms on Rotavirus Susceptibility, Epidemiology, and Vaccine Take. Viruses 2020, 12, 324. [Google Scholar] [CrossRef] [Green Version]
- Heggelund, J.E.; Varrot, A.; Imberty, A.; Krengel, U. Histo-blood group antigens as mediators of infections. Curr. Opin. Struct. Biol. 2017, 44, 190–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárcamo-Calvo, R.; Muñoz, C.; Buesa, J.; Rodríguez-Díaz, J.; Gozalbo-Rovira, R. The Rotavirus Vaccine Landscape, an Update. Pathogens 2021, 10, 520. [Google Scholar] [CrossRef] [PubMed]
- Imbert-Marcille, B.M.; Barbé, L.; Dupé, M.; Le Moullac-Vaidye, B.; Besse, B.; Peltier, C.; Ruvoën-Clouet, N.; Le Pendu, J. A FUT2 gene common polymorphism determines resistance to Rotavirus A of the P[8] genotype. J. Infect. Dis. 2013, 209, 1227–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Xia, M.; Tan, M.; Zhong, W.; Wei, C.; Wang, L.; Morrow, A.; Jiang, X. Spike protein VP8* of human rotavirus recognizes histo-blood group antigens in a type-specific manner. J. Virol. 2012, 86, 4833–4843. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Huang, P.; Tan, M.; Biesiada, J.; Meller, J.; Castello, A.A.; Jiang, B.; Jiang, X. Rotavirus VP8*: Phylogeny, host range, and interaction with histo-blood group antigens. J. Virol. 2012, 86, 9899–9910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Jiang, X.; Huang, P.; Jiang, B.; Tan, M.; Morrow, A.L. Poly-LacNAc as an Age-Specific Ligand for Rotavirus P[11] in Neonates and Infants. PLoS ONE 2013, 8, e78113. [Google Scholar] [CrossRef] [Green Version]
- Khambhampati, A.; Payne, D.C.; Constantini, V.; Lopman, B.A. Host genetic susceptibility to enteric viruses: A systematic review and meta-analysis. Clin. Infect. Dis. 2016, 62, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.-A.; Zhang, M.; Hou, Y.Z.; Hu, H.; Fang, L.; Tan, M.; Huang, Q.; Li, H.; Sun, L.-M.; Jiang, X.; et al. Epidemiology and HBGA-susceptibility investigation of a G9P[8] rotavirus outbreak in a school in Lechang, China. Arch. Virol. 2020, 165, 1311–1320. [Google Scholar] [CrossRef]
- Cantelli, C.P.; Velloso, A.J.; Assis, R.M.S.; de-Barros, J.J.; Mello, F.C.d.A.; Cunha, D.C.d.; Brasil, P.; Nordgren, J.; Svensson, L.; Miagostovich, M.P.; et al. Rotavirus A shedding and HBGA host genetic susceptibility in a birth community-cohort, Rio de Janeiro, Brazil, 2014–2018. Sci. Rep. 2020, 10, 6965. [Google Scholar] [CrossRef]
- Li, B.; Ding, S.; Feng, N.; Mooney, N.; Ooi, Y.S.; Ren, L.; Diep, J.; Kelly, M.R.; Yasukawa, L.L.; Patton, J.T.; et al. Drebrin restricts rotavirus entry by inhibiting dynamin-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, E3642–E3651. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, T.; Torres, R.; Salgado, E.N.; Berciu, C.; Stoddard, D.; Nicastro, D.; Jenni, S.; Harrison, S.C. Functional refolding of the penetration protein on a non-enveloped virus. Nature 2021, 590, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Kapikian, A.Z.; Chanock, R.M. Rotaviruses. In Fields Virology, 3rd ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Chanock, R.M., Monath, T.P., Melnick, J.L., Eds.; Raven Press: New York, NY, USA, 1996; Volume 2, pp. 1657–1708. [Google Scholar]
- O’Ryan, M.G. Clinical Manifestations and Diagnosis of Rotavirus Infection: An Update. Wolters Kluwers. Available online: https://www.uptodate.com/contents/clinical-manifestations-and-diagnosis-of-rotavirus-infection#H3096970958 (accessed on 12 September 2020).
- Hyams, J.S.; Krause, P.J.; Gleason, P.A. Lactose malabsorption following rotavirus infection in young children. J. Pediatr. 1981, 99, 916. [Google Scholar] [CrossRef]
- Seo, N.-N.; Carl, Q.-Y.; Utama, B.; Crawford, S.E.; Kim, K.J.; Höök, M.; Estes, M.K. Integrins α1β1 and α2β1 are receptors for the rotavirus enterotoxin. Proc. Natl. Acad. Sci. USA 2008, 105, 8811–8818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyser, J.M.; Collinson-Pautz, M.R.; Utama, B.; Estes, M.K. Rotavirus disrupts calcium homeostasis by NSP4 viroporin activity. MBio 2010, 1, e00265-10. [Google Scholar] [CrossRef] [Green Version]
- Farnworth, E.R. The evidence to support health claims for probiotics. J. Nutr. 2008, 138, 1250S–1254S. [Google Scholar] [CrossRef]
- Arya, S.C. Rotaviral Infection and Intestinal Lactase Level. J. Infect. Dis. 1984, 150, 791. [Google Scholar] [CrossRef]
- Bialowas, S.; Hagbom, M.; Nordgren, J.; Karlsson, T.; Sharma, S.; Magnusson, K.E.; Svensson, L. Rotavirus and serotonin cross-talk in diarrhoea. PLoS ONE 2016, 11, e0159660. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, O.; Peregrin, A.T.; Persson, K.; Kordasti, S.; Uhnoo, I.; Svensson, L. Role of the enteric nervous system in the fluid and electrolyte secretion of rotavirus diarrhea. Science 2000, 287, 409–411. [Google Scholar] [CrossRef]
- Bass, E.S.; Pappano, D.A.; Humiston, S.G. Rotavirus. Pediatr. Rev. 2007, 28, 183–191. [Google Scholar] [CrossRef]
- Marie, H.; Claudia, I.; David, E.; Thommie, K.; Jesus, R.; Javier, B.; John, A.T. Rotavirus stimulates release of serotonin from human enterochromaffin cells and activates brain cells involved in nausea and vomiting. PLoS Pathog. 2011, 7, 10–131. [Google Scholar]
- Ray, P.; Fenaux, M.; Sharma, S.; Malik, J.; Subodh, S.; Bhatnagar, S.; Greenberg, H.; Glass, R.I. Quantitative evaluation of rotaviral antigenemia in children with acute rotaviral diarrhea. J. Infect Dis. 2006, 194, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Rial, J.; Sánchez-Batán, S.; Rivero-Calle, I.; Pardo-Seco, J.; Martinón-Martínez, J.M.; Salas, A.; Martinón-Torres, F. Rotavirus infection beyond the gut. Infect. Drug Resist. 2019, 12, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero-Calle, I.; Gómez-Rial, J.; Martinón-Torres, F. Systemic features of rotavirus infection. J. Inf. 2016, 72, S98–S105. [Google Scholar] [CrossRef] [PubMed]
- Hemming, M.; Huhti, L.; Räsänen, S.; Salminen, M.; Vesikari, T. Rotavirus antigenemia in children is associated with more severe clinical manifestations of acute gastroenteritis. Pediatr. Infect. Dis. J. 2014, 33, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Sugata, K.; Taniguchi, K.; Yui, A.; Miyake, F.; Suga, S.; Asano, Y.; Ohashi, M.; Suzuki, K.; Nishimura, N.; Osaki, T.; et al. Analysis of rotavirus antigenemia and extraintestinal manifestations in children with rotavirus gastroenteritis. Pediatrics 2008, 122, 392–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canada Communicable Disease Report (CCDR). In Literature Review on Rotavirus: Disease and Vaccine Characteristics. An Advisory Committee Statement (ACS) by National Advisory Committee on Immunization (NACI); Public Health Agency of Canada: Ottawa, Canada, 2010; Volume 36.
- Payne, D.C.; Parashar, U.D. Rotavirus. VPD Surveillance Manual Rotavirus: Chapter 13.1. 2017; pp. 1–17. Available online: https://www.cdc.gov/vaccines/pubs/surv-manual/chpt13-rotavirus.pdf (accessed on 10 November 2021).
- Kumar, A.; Vlasova, A.N.; Deblais, L.; Huang, H.C.; Wijeratne, A.; Kandasamy, S.; Fischer, D.D.; Langel, S.N.; Paim, F.C.; Alhamo, M.A.; et al. Impact of nutrition and rotavirus infection on the infant gut microbiota in a humanized pig model. BMC Gastroenterol. 2018, 18, 93. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ochoa, G.; Flores-Mendoza, L.K.; Icedo-Garcia1, R.; Gomez-Flores, R.; Tamez-Guerra, P. Modulation of rotavirus severe gastroenteritis by the combination of probiotics and prebiotics. Arch. Microbiol. 2017, 199, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Athiyyah, A.F.; Utsumi, T.; Wahyuni, R.M.; Dinana, Z.; Yamani, L.N.; Sudarmo, S.M.; Ranuh, R.G.; Darma, A.; Juniastuti, J.; Raharjo, D.; et al. Molecular Epidemiology and Clinical Features of Rotavirus Infection among Pediatric Patients in East Java, Indonesia during 2015–2018: Dynamic Changes in Rotavirus Genotypes from Equine-Like G3 to Typical Human G1/G3. Front. Microbiol. 2019, 10, 940. [Google Scholar] [CrossRef] [Green Version]
- Senecal, M.; Brisson, M.; Lebel, M.H.; Yaremko, J.; Wong, R.; Gallant, L.A. Severity, healthcare resource use and work loss related to rotavirus gastroenteritis: A prospective study in community practice. Canadian Public Health Association. Vancouver 2006, 3, 28–31. [Google Scholar]
- Leung, A.K.; Kellner, J.D.; Davies, H.D. Rotavirus gastroenteritis. Adv. Ther. 2005, 22, 476–487. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention/National Center for Immunization and Respiratory Diseases (CDC/NCIRD). Division of Viral Diseases. Updated: 5 November 2019. Available online: https://www.cdc.gov/ (accessed on 16 May 2020).
- Glass, R.I.; Parashar, U.D.; Bresee, J.S.; Turcios, R.; Fischer, T.K.; Widdowson, M.A.; Jiang, B. Rotavirus vaccines: Current prospects and future challenges. Lancet 2006, 368, 323–332. [Google Scholar] [CrossRef]
- Sen, A.; Greenberg, H.B. Innate Immune Responses to Rotavirus Infection; Academic Press: London, UK, 2016; pp. 243–263. [Google Scholar]
- Reikine, S.; Nguyen, J.B.; Modis, Y. Pattern recognition and signaling mechanisms of RIG-I and MDA5. Front. Immunol. 2014, 5, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, M.M.; Patton, J.T. Diversity of interferon antagonist activities mediated by NSP1 proteins of different rotavirus strains. J. Virol. 2011, 85, 1970–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villena, J.; Vizoso-Pinto, M.G.; Kitazawa, H. Intestinal Innate Antiviral Immunity and Immunobiotics: Beneficial Effects against Rotavirus Infection. Front. Immunol. 2016, 7, 563. [Google Scholar] [CrossRef] [Green Version]
- Holloway, G.; Coulson, B.S. Innate cellular responses to rotavirus infection. J. Gen. Virol. 2013, 94, 1151–1160. [Google Scholar] [CrossRef]
- Uchiyama, R.; Chassaing, B.; Zhang, B.; Gewirtz, A.T. MyD88-mediated TLR signaling protects against acute rotavirus infection while inflammasome cytokines direct Ab response. Innate Immun. 2015, 21, 41628. [Google Scholar] [CrossRef]
- Sánchez-Tacuba, L.; Rojas, M.; Arias, C.F.; López, S. Rotavirus Controls Activation of the 2′-5′-Oligoadenylate Synthetase/RNase L Pathway Using at least Two Distinct Mechanisms. J. Virol. 2015, 89, 12145–12153. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Feng, N.; Sanchez-Tacuba, L.; Yasukawa, L.L.; Ren, L.; Silverman, R.H.; Ding, S.; Greenberg, H.B. Reverse Genetics Reveals a Role of Rotavirus VP3 Phosphodiesterase Activity in Inhibiting RNase L Signaling and Contributing to Intestinal Viral Replication In Vivo. J. Virol. 2020, 94, e01952-19. [Google Scholar] [CrossRef]
- Zhu, S.; Ding, S.; Wang, P.; Wei, Z.; Pan, W.; Palm, N.W.; Yang, Y.; Yu, H.; Li, H.B.; Wang, G.; et al. Nlrp9b inflammasome restricts rotavirus infection in intestinal epithelial cells. Nature 2017, 546, 667–670. [Google Scholar] [CrossRef]
- Desselberger, U.; Huppertz, H.I. Immune responses to rotavirus infection and vaccination and associated correlates of protection. J. Infect. Dis. 2011, 203, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Glass, R.I.; Jiang, B.; Santosham, M.; Lopman, B.; Parashar, U. A systematic review of anti-rotavirus serum IgA antibody titer as a potential correlate of rotavirus vaccine efficacy. J. Infect. Dis. 2013, 208, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, M.A.; Angel, J.; Greenberg, H.B. Immunity and correlates of protection for rotavirus vaccines. Vaccine 2006, 24, 2718–2731. [Google Scholar] [CrossRef]
- Green, K.Y.; Taniguchi, K.; Mackow, E.R.; Kapikian, A.Z. Homotypic and heterotypic epitope-specific antibody responses in adult and infant rotavirus vaccinees: Implications for vaccine development. J. Infect. Dis. 1990, 161, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, F.R.; Matson, D.O.; Guerrero, M.L.; Shults, J.; Calva, J.J.; Morro, A.L. Natural Immunity to Rotavirus Infection in Children. J. Infect. Dis. 2000, 182, 1602–1609. [Google Scholar] [PubMed] [Green Version]
- Matson, D.O.; O’Ryan, M.L.; Herrera, I.; Pickering, L.K.; Estes, M.K. Fecal antibody responses to symptomatic asymptomatic rotavirus infections. J. Infect. Dis. 1993, 167, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Kanungo, S.; Kim, D.R.; Manna, B.; Song, M.; Park, J.Y.; Haldar, B.; Sharma, P.; Mallick, A.H.; Kim, S.A.; et al. Antibody secreting B cells and plasma antibody response to rotavirus vaccination in infants from Kolkata India. Heliyon 2018, 4, e00519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caddy, S.L.; Vaysburd, M.; Wing, M.; Foss, S.; Andersen, J.T.; O’Connell, K. Intracellular neutralisation of rotavirus by VP6-specific IgG. PLoS Pathog 2020, 16, e1008732. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Hyöty, H.; Knip, M.; Vesikari, T.; Blazevic, V. Development of T cell immunity to norovirus and rotavirus in children under five years of age. Sci. Rep. 2019, 9, 3199. [Google Scholar] [CrossRef] [Green Version]
- Smiley, K.L.; McNeal, M.M.; Basu, M.; Choi, A.H.; Clements, J.D.; Ward, R.L. Association of gamma interferon and interleukin-17 production in intestinal CD4+ T cells with protection against rotavirus shedding in mice intranasally immunized with VP6 and the adjuvant LT(R192G). J. Virol. 2007, 81, 3740–3748. [Google Scholar] [CrossRef] [Green Version]
- Godefroy, E.; Alameddine, J.; Montassier, E.; Mathé, J.; Desfrançois-Noël, J.; Marec, N.; Bossard, C.; Jarry, A.; Bridonneau, C.; Le Roy, A.; et al. Expression of CCR6 and CXCR6 by Gut-Derived CD4+/CD8α+ T-Regulatory Cells, Which Are Decreased in Blood Samples from Patients with Inflammatory Bowel Diseases. Gastroenterology 2018, 155, 1205–1217. [Google Scholar] [CrossRef]
- Makela, M.; Marttila, J.; Simell, O.; Ilonen, J. Rotavirus-specific T-cell responses in young prospectively followed-up children. Clin. Exp. Immunol. 2004, 137, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.A.; Greenberg, H.B. Immunity to rotavirus in T cell deficient mice. Virology 1997, 238, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Sastre, A.; Biron, C.A. Type 1 interferons and the virus-host relationship: A lesson in detente. Science 2006, 312, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Dennehy, P.; Keyserling, H.; Westerman, L.E.; Wang, Y.; Holman, R.C.; Gentsch, J.R.; Glass, R.I.; Jiang, B. Serum Antibody Responses in Children with Rotavirus Diarrhea Can Serve as Proxy for Protection. Clin. Diagn. Lab. Immunol. 2005, 12, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Villamizar-Gallardo, R.A.; Osma, J.F.; Ortíz, O.O. New technique for direct fluoroimmunomagnetic detection of rotavirus in water samples. J. Water Health 2017, 15, 932–941. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Rotavirus vaccines: An update. Wkly. Epidemiol. Rec. 2009, 84, 533–537. [Google Scholar]
- El-Ageery, M.S.; Ali, R.; El-Khier, N.T.A.; Rakha, S.A.; Zeid, M.S. Comparison of enzyme immunoassay, latex agglutination and polyacrylamide gel electrophoresis for diagnosis of rotavirus in children. Egypt. J. Basic Appl. Sci. 2020, 7, 47–52. [Google Scholar] [CrossRef]
- Carossino, M.; Barrandeguy, M.E.; Erol, E.; Li, Y.; Balasuriya, U. Development and evaluation of a one-step multiplex real-time TaqMan® RT-qPCR assay for the detection and genotyping of equine G3 and G14 rotaviruses in fecal samples. Virol. J. 2019, 16, 49. [Google Scholar] [CrossRef]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D. WHO-coordinated Global Rotavirus Surveillance Network. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Badura, S.; Öztürka, S.; Pereira, P.; AbdelGhanyc, M.; Khalafd, M.; Lagoubie, Y.; Hanifg, O.K.; Saha, D. Systematic review of the rotavirus infection burden in the WHO-EMRO region. Hum. Vaccines Immunother. 2019, 15, 2754–2768. [Google Scholar] [CrossRef] [PubMed]
- Parashar, U.D.; Burton, A.; Lanata, C.; Boschi-Pinto, C.; Shibuya, K.; Steele, D.; Birmingham, M.; Glass, R.I. Global mortality associated with rotavirus disease among children in 2004. J. Infect. Dis. 2009, 200, S9–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez-Domingo, J.; Garces-Sanchez, M.; Gimenez-Sanchez, F.; Colomina-Rodriguez, J.; Martinon-Torres, F. What have we learnt about rotavirus in Spain in the last 10 years? An. Pediatr. 2019, 91, 166–179. [Google Scholar] [CrossRef]
- Rota Council. Global Introduction Status. 2017. Available online: http://rotacouncil.org/vaccine-introduction/globalintroduction-status/ (accessed on 5 December 2017).
- Ardura-Garcia, C.; Kreis, C.; Rakic, M.; Jaboyedoff, M.; Mallet, M.C.; Low, N.; Kuehni, C.E. Rotavirus disease and health care utilisation among children under 5 years of age in highly developed countries: A systematic review and meta-analysis. Vaccine 2021, 39, 2917–2928. [Google Scholar] [CrossRef]
- Anderson, E.J.; Weber, S.G. Rotavirus infection in adults. Lancet Infect. Dis. 2004, 4, 91–99. [Google Scholar] [CrossRef]
- Linhares, A.C.; Pinheiro, F.P.; Freitas, R.B.; Gabbay, Y.B.; Shirley, J.A.; Beards, G.M. An outbreak of rotavirus diarrhea among a non-immune, isolated South American Indian community. Am. J. Epidemiol. 1981, 113, 703–710. [Google Scholar] [CrossRef]
- Bucardo, F.; Karlsson, B.; Nordgren, J.; Paniagua, M.; González, A.; Amador, J.J.; Espinoza, F.; Svensson, L. Mutated G4P[8] rotavirus associated with a nationwide outbreak of gastroenteritis in Nicaragua in 2005. J. Clin. Microbiol. 2007, 45, 990–997. [Google Scholar] [CrossRef] [Green Version]
- WHO. Africa. WHO Supports Botswana to Respond to an Outbreak of Diarrhoea in Children below Five Years of Age. 29 October 2018. Available online: https://www.afro.who.int/news/who-supports-botswana-respond-outbreak-diarrhoea-children-below-five-years-age (accessed on 24 February 2022).
- Lahon, A.; Maniya, N.H.; Tambe, G.U.; Chinchole, P.R.; Purwar, S.; Jacob, G.; Chitambar, S.D. Group B rotavirus infection in patients with acute gastroenteritis from India: 1994–1995 and 2004–2010. Epidemiol. Infect. 2013, 141, 969–975. [Google Scholar] [CrossRef]
- Omatola, C.A.; Ogunsakin, R.E.; Olaniran, A.O. Prevalence, Pattern and Genetic Diversity of Rotaviruses among Children under 5 Years of Age with Acute Gastroenteritis in South Africa: A Systematic Review and Meta-Analysis. Viruses 2021, 13, 1905. [Google Scholar] [CrossRef]
- Li, K.; Lin, X.D.; Huang, K.Y.; Zhang, B.; Shi, M.; Guo, W.P.; Wang, M.R.; Wang, W.; Xing, J.G.; Li, M.H.; et al. Identification of novel and diverse rotaviruses in rodents and insectivores, and evidence of cross-species transmission into humans. Virology 2016, 494, 168–177. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Rotavirus Information and Surveillance Bulletin. Reporting Period: January through December 2010; World Health Organization: Geneva, Switzerland, 2011; Volume 4, p. 2011.
- Seheri, L.M.; Magagula, N.B.; Peenze, I.; Rakau, K.; Ndadza, A.; Mwenda, J.M.; Weldegebriel, G.; Steele, A.D.; Mphahlele, M.J. Rotavirus strain diversity in Eastern and Southern African countries before and after vaccine introduction. Vaccine 2018, 36, 7222–7230. [Google Scholar] [CrossRef] [PubMed]
- Rakau, K.G.; Nyaga, M.M.; Gededzha, M.P.; Mwenda, J.M.; Mphahlele, M.J.; Seheri, L.M.; Steele, A.D. Genetic characterization of G12P[6] and G12P[8] rotavirus strains collected in six African countries between 2010 and 2014. BMC Infect. Dis. 2021, 21, 107. [Google Scholar] [CrossRef] [PubMed]
- Azemi, M.; Berisha, M.; Ismaili-Jaha, V.; Kolgeci, S.; Avdiu, M.; Jakupi, X.; Hoxha, R.; Hoxha-Kamberi, T. Socio-demographic, clinical and laboratory features of rotavirus gastroenteritis in children treated in pediatric clinic. Mat. Soc. Med. 2013, 25, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wobudeya, E.; Bachou, H.; Karamagi, C.K.; Kalyango, J.N.; Mutebi, E.; Wamani, H. Breastfeeding and the risk of rotavirus diarrhea in hospitalized infants in Uganda: A matched case control study. BMC Pediatr. 2011, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Waure, C.; Sarnari, L.; Chiavarini, M.; Ianiro, G.; Monini, M.; Alunno, A.; Camilloni, B. 10-Year Rotavirus Infection Surveillance: Epidemiological Trends in the Pediatric Population of Perugia Province. Int. J. Environ. Res. Pub. Health 2020, 17, 1008. [Google Scholar] [CrossRef] [Green Version]
- Lestari, F.B.; Vongpunsawad, S.; Wanlapakorn, N.; Poovorawan, Y. Rotavirus infection in children in Southeast Asia 2008–2018: Disease burden, genotype distribution, seasonality, and vaccination. J. Biomed. Sci. 2020, 27, 66. [Google Scholar] [CrossRef]
- Patel, M.; Pedreira, C.; De Oliveira, L.H.; Umaña, J.; Tate, J.; Lopman, B.; Sanchez, E.; Reyes, M.; Mercado, J.; Gonzalez, A. Duration of protection of pentavalent rotavirus vaccination in Nicaragua. Pediatrics 2012, 130, e365–e372. [Google Scholar] [CrossRef]
- Center for Disease Prevention and Control/National Center for Immunization and Respiratory Diseases, Division of Viral Diseases (CDC/NCIRD). Rotavirus Transmission. Page Last Reviewed: 26 March 2021. Available online: https://www.cdc.gov/rotavirus/about/transmission.html (accessed on 19 May 2021).
- Pitzer, V.E.; Viboud, C.; Lopman, B.A.; Patel, M.M.; Parashar, U.D.; Grenfell, B.T. Influence of birth rates and transmission rates on the global seasonality of rotavirus incidence. J. R. Soc. Interface 2011, 8, 1584–1593. [Google Scholar] [CrossRef]
- Pitzer, V.E.; Viboud, C.; Simonsen, L.; Steiner, C.; Panozzo, C.A.; Alonso, W.J.; Miller, M.A.; Glass, R.I.; Glasser, J.W.; Parashar, U.D.; et al. Demographic variability, vaccination, and the spatiotemporal dynamics of rotavirus epidemics. Science 2009, 325, 290–294. [Google Scholar] [CrossRef] [Green Version]
- Johansen, K.; Hedlund, K.-O.; Zweygberg-Wirgart, B.; Bennet, R. Complications attributable to rotavirus-induced diarrhoea in a Swedish paediatric population: Report from an 11-year surveillance. Scand. J. Infect. Dis. 2008, 40, 958–964. [Google Scholar] [CrossRef]
- Stefkovicov, M.; Simurka, P.; Jurackov, L.; Hudeckov, H.; Mad’ar, R. Nosocomial rotaviral gastroenteritis in paediatric departments. Cent. Eur. J. Public Health 2008, 16, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, A.N.; Husberg, M.; Bennet, R.; Brytting, M.; Carlsson, P.; Eriksson, M.; Storsaeter, J.; Österlin, B.; Johansen, K. Impact on affected families and society of severe rotavirus infections in Swedish children assessed in a prospective cohort study. Infect. Dis. 2018, 50, 361–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, R.M.; Tate, J.E.; Kirkwood, C.D.; Steele, A.D.; Parashar, U.D. Current and new rotavirus vaccines. Curr. Opin. Infect. Dis. 2019, 32, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.; Gardner, R.C.; Debrus, S.; Benninghoff, B.; Pereira, P. Established and new rotavirus vaccines: A comprehensive review for healthcare professionals [published online ahead of print, 19 February 2021]. Hum. Vaccin. Immunother. 2021, 18, 1–17. [Google Scholar]
- Soares-Weiser, K.; Bergman, H.; Henschke, N.; Pitan, F.; Cunliffe, N. Vaccines for preventing rotavirus diarrhoea: Vaccines in use. Cochrane Database of Systematic Reviews. Cochrane Database Syst. Rev. 2019, 3, CD008521. [Google Scholar]
- Sengupta, P. Rotavirus: The challenges ahead. Indian J Community Med. 2009, 34, 279–282. [Google Scholar] [CrossRef]
- Dang, D.A.; Nguyen, V.T.; Vu, D.T.; Nguyen, T.H.; Nguyen, D.M.; Yuhuan, W.; Baoming, J.; Nguyen, D.H.; Le, T.L.; Rotavin-M1 Vaccine Trial Group. A dose-escalation safety and immunogenicity study of a new live attenuated human rotavirus vaccine (Rotavin-M1) in Vietnamese children. Vaccine 2012, 30, A114–A121. [Google Scholar]
- Skansberg, A.; Sauer, M.; Tan, M.; Santosham, M.; Jennings, M.C. Product review of the rotavirus vaccines ROTASIIL, ROTAVAC, and Rotavin-M1. Hum. Vaccine. Immunother. 2021, 17, 1223–1234. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Yang, Y.; Liang, Z.; Tian, Y.; Liu, B.; Gao, Z.; Jia, L.; Chen, L.; Wang, Q. Effectiveness of Lanzhou lamb rotavirus vaccine in preventing gastroenteritis among children younger than 5 years of age. Vaccine 2019, 37, 3611–3616. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention; National Center for Emerging and Zoonotic Infectious Diseases (NCEZID); Division of Foodborne, Waterborne, and Environmental Diseases (DFWED). Rotavirus and Drinking Water from Private Wells. Page Last Reviewed: 1 July 2019. Available online: https://www.cdc.gov/healthywater/drinking/private/wells/disease/rotavirus.html (accessed on 20 April 2020).
- Pindyck, T.; Tate, J.E.; Parashar, U.D. A decade of experience with rotavirus vaccination in the United States—vaccine uptake, effectiveness, and impact. Expert Rev. Vaccines 2018, 17, 593–606. [Google Scholar] [CrossRef]
- Mphahlele, M.J.; Groome, M.J.; Page, N.A.; Bhagwandin, N.; Mwenda, J.M.; Steele, A.D. A decade of rotavirus vaccination in Africa–Saving lives and changing the face of diarrhoeal diseases: Report of the 12th African Rotavirus Symposium. Vaccine 2021, 39, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Uprety, T.; Wang, D.; Li, F. Recent advances in rotavirus reverse genetics and its utilization in basic research and vaccine development. Arch. Virol. 2021, 166, 2369–2386. [Google Scholar] [CrossRef] [PubMed]
- Philip, A.A.; Patton, J.T. Expression of Separate Heterologous Proteins from the Rotavirus NSP3 Genome Segment Using a Translational 2A Stop-Restart Element. J. Virol. 2020, 94, e00959-20. [Google Scholar] [CrossRef] [PubMed]
- Philip, A.A.; Patton, J.T. Rotavirus as an expression platform of domains of the SARS-CoV-2 spike protein. Vaccines 2021, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Vesikari, T.; Karvonen, A.; Prymula, R.; Schuster, V.; Tejedor, J.C.; Cohen, R.; Meurice, F.; Han, H.H.; Damaso, S.; Bouckenooghe, A. Efficacy of human rotavirus vaccine against rotavirus gastroenteritis during the first 2 years of life in European infants: Randomised, double-blind controlled study. Lancet 2007, 370, 1757–1763. [Google Scholar] [CrossRef]
- Ruiz-Palacios, G.M.; Pérez-Schael, I.; Velázquez, F.R.; Abate, H.; Breuer, T.; Clemens, S.C.; Cheuvart, B.; Espinoza, F.; Gillard, P.; Innis, B.L.; et al. Safety and Efficacy of an Attenuated Vaccine against Severe Rotavirus Gastroenteritis. N. Engl. J. Med. 2006, 354, 11–22. [Google Scholar] [CrossRef]
- Madhi, S.A.; Cunliffe, N.A.; Steele, D.; Witte, D.; Kirsten, M.; Louw, C.; Ngwira, B.; Victor, J.C.; Gillard, P.H.; Cheuvart, B.B.; et al. Effect of human rotavirus vaccine on severe diarrhea in African infants. N. Engl. J. Med. 2010, 362, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Vesikari, T.; Matson, D.O.; Dennehy, P.; Van Damme, P.; Santosham, M.; Rodriguez, Z.; Dallas, M.J.; Heyse, J.F.; Goveia, M.G.; Black, S.B.; et al. Safety and Efficacy of a Pentavalent Human–Bovine (WC3) Reassortant Rotavirus Vaccine. N. Engl. J. Med. 2006, 354, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Armah, G.E.; Sow, S.O.; Breiman, R.F.; Dallas, M.J.; Tapia, M.D.; Feikin, D.R.; Binka, F.N.; Steele, A.D.; Laserson, K.F.; Ansah, N.A.; et al. Efficacy of pentavalent rotavirus vaccine against severe rotavirus gastroenteritis in infants in developing countries in sub-Saharan Africa: A randomised, double-blind, placebo-controlled trial. Lancet 2010, 376, 606–614. [Google Scholar] [CrossRef]
- Zaman, K.; Dang, D.A.; Victor, J.C.; Shin, S.; Yunus, M.; Dallas, M.J.; Podder, G.; Vu, D.T.; Le, T.P.; Luby, S.P.; et al. Efficacy of pentavalent rotavirus vaccine against severe rotavirus gastroenteritis in infants in developing countries in Asia: A randomised, double-blind, placebo-controlled trial. Lancet 2010, 376, 615–623. [Google Scholar] [CrossRef]
- Bhandari, N.; Rongsen-Chandola, T.; Bavdekar, A.; John, J.; Antony, K.; Taneja, S.; Goyal, N.; Kawade, A.; Kang, G.; Rathore, S.S.; et al. Efficacy of a monovalent human-bovine (116E) rotavirus vaccine in Indian children in the second year of life. Vaccine 2014, 32, A110–A116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isanaka, S.; Guindo, O.; Langendorf, C.; Seck, M.A.; Plikaytis, B.D.; Sayinzoga-Makombe, N.; McNeal, M.M.; Meyer, N.; Adehossi, E.; Djibo, A.; et al. Efficacy of a Low-Cost, Heat-Stable Oral Rotavirus Vaccine in Niger. N. Engl. J. Med. 2017, 376, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.S.; Desai, S.; Tewari, T.; Kawade, A.; Goyal, N.; Garg, B.S.; Kanungo, S.; Kamat, V.; Kang, G.; SII BRV-PV author group (2017); et al. A randomized Phase III clinical trial to assess the efficacy of a bovine-human reassortant pentavalent rotavirus vaccine in Indian infants. Vaccine 2017, 35, 6228–6237. [Google Scholar] [CrossRef]
- Church, J.A.; Rukobo, S.; Govha, M.; Lee, B.; Carmolli, M.P.; Chasekwa, B.; Prendergast, A.J. The Impact of Improved Water, Sanitation, and Hygiene on Oral Rotavirus Vaccine Immunogenicity in Zimbabwean Infants: Substudy of a Cluster-randomized Trial. Clin. Infect. Dis. 2019, 69, 2074–2081. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Takanashi, S.; Miyazaki, A.; Rajashekara, G.; Saif, L.J. How the gut microbiome regulates host immune responses to viral vaccines. Curr. Opin. Virol. 2019, 37, 16–25. [Google Scholar] [CrossRef]
- Srivastava, V.; Deblais, L.; Huang, H.C.; Miyazaki, A.; Kandasamy, S.; Langel, S.N.; Paim, F.C.; Chepngeno, J.; Kathayat, D.; Vlasova, A.N.; et al. Reduced rotavirus vaccine efficacy in protein malnourished human-faecal-microbiota-transplanted gnotobiotic pig model is in part attributed to the gut microbiota. Benef. Microbes 2020, 11, 733–751. [Google Scholar] [CrossRef]
- WHO. Malnutrition. Updated: 9 June 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/malnutrition (accessed on 27 July 2021).
- Kandasamy, S.; Chattha, K.S.; Vlasova, A.N.; Saif, L.J. Prenatal vitamin A deficiency impairs adaptive immune responses to pentavalent rotavirus vaccine (RotaTeq®) in a neonatal gnotobiotic pig model. Vaccine 2014, 32, 816–824. [Google Scholar] [CrossRef]
- Mora, J.R.; Iwata, M.; Eksteen, B.; Song, S.Y.; Junt, T.; Senman, B.; Otipoby, K.L.; Yokota, A.; Takeuchi, H.; Ricciardi-Castagnoli, P.; et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science 2006, 314, 1157–1160. [Google Scholar] [CrossRef] [Green Version]
- Perez-Schael, I.; Salinas, B.; Tomat, M.; Linhares, A.C.; Guerrero, M.L.; Ruiz-Palacios, G.M. Efficacy of the human rotavirus vaccine RIX4414 in malnourished children. J. Infect. Dis. 2007, 196, 537–540. [Google Scholar] [CrossRef]
- Linhares, A.C.; Carmo, K.B.; Oliveira, K.K.; Oliveira, C.S.; Freitas, R.B.; Bellesi, N. Nutritional status in relation to the efficacy of the rhesus-human reassortant, tetravalent rotavirus vaccine (RRV-TV) in infants from Belem, para state, Brazil. Rev. Inst. Med. Trop. São Paulo 2002, 44, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Michael, H.; Langel, S.N.; Miyazaki, A.; Paim, F.C.; Chepngeno, J.; Alhamo, M.A.; Fischer, D.D.; Srivastava, V.; Kathayat, D.; Deblais, L.; et al. Malnutrition Decreases Antibody Secreting Cell Numbers Induced by an Oral Attenuated Human Rotavirus Vaccine in a Human Infant Fecal Microbiota Transplanted Gnotobiotic Pig Model. Front. Immunol. 2020, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, A.; Kandasamy, S.; Michael, H.; Langel, S.N.; Paim, F.C.; Chepngeno, J. Protein deficiency reduces efficacy of oral attenuated human rotavirus vaccine in a human infant fecal microbiota transplanted gnotobiotic pig model. Vaccine 2018, 36, 6270–6281. [Google Scholar] [CrossRef] [PubMed]
- Appaiahgari, M.B.; Glass, R.; Singh, S.; Taneja, S.; Rongsen-Chandola, T.; Bhandari, N.; Mishra, S.; Vrati, S. Transplacental rotavirus IgG interferes with immune response to live oral rotavirus vaccine ORV-116E in Indian infants. Vaccine 2014, 32, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.S.; Tate, J.E.; Ray, P.; Dennehy, P.H.; Archary, D.; Coutsoudis, A.; Bland, R.; Newell, M.L.; Glass, R.I.; Parashar, U.; et al. Differential profiles and inhibitory effect on rotavirus vaccines of nonantibody components in breast milk from mothers in developing and developed countries. Pediatr. Infect. Dis. J. 2013, 32, 863–870. [Google Scholar] [CrossRef] [Green Version]
- Becker-Dreps, S.; Vilchez, S.; Velasquez, D.; Moon, S.S.; Hudgens, M.G.; Zambrana, L.E.; Jiang, B. Rotavirus-specific IgG antibodies from mothers’ serum may inhibit infant immune responses to the pentavalent rotavirus vaccine. Pediatr. Infect. Dis. J. 2015, 34, 115–116. [Google Scholar] [CrossRef] [Green Version]
- Rongsen-Chandola, T.; Strand, T.A.; Goyal, N. Effect of withholding breastfeeding on the immune response to a live oral rotavirus vaccine in North Indian infants. Vaccine 2014, 32, A134–A139. [Google Scholar] [CrossRef] [Green Version]
- Groome, M.J.; Moon, S.S.; Velasquez, D.; Jones, S.; Koen, A.; van Niekerk, N.; Jiang, B.; Parashar, U.D.; Madhi, S.A. Effect of breastfeeding on immunogenicity of oral live-attenuated human rotavirus vaccine: A randomized trial in HIV-uninfected infants in Soweto, South Africa. Bull. World Health Organ 2014, 92, 238–245. [Google Scholar] [CrossRef]
- Ali, A.; Kazi, A.M.; Cortese, M.M.; Fleming, J.A.; Moon, S.; Parashar, U.D.; Jiang, B.; McNeal, M.M.; Steele, D.; Bhutta, Z.; et al. Impact of withholding breastfeeding at the time of vaccination on the immunogenicity of oral rotavirus vaccine—A randomized trial. PLoS ONE 2015, 10, e0127622. [Google Scholar]
- Mwila, K.; Chilengi, R.; Simuyandi, M.; Permar, S.R.; Becker-Dreps, S. Contribution of Maternal Immunity to Decreased Rotavirus Vaccine Performance in Low- and Middle-Income Countries. Clin. Vaccine Immunol. 2017, 24, e00405–e00416. [Google Scholar] [CrossRef] [Green Version]
- Emperador, D.M.; Velasquez, D.E.; Estivariz, C.F.; Lopman, B.; Jiang, B.; Parashar, U.; Anand, A.; Zaman, K. Interference of monovalent, bivalent, and trivalent oral poliovirus vaccines on monovalent rotavirus vaccine immunogenicity in rural Bangladesh. Clin. Infect. Dis. 2016, 62, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Steele, A.D.; De Vos, B.; Tumbo, J.; Reynders, J.; Scholtz, F.; Bos, P.; de Beer, M.C.; Van der Merwe, C.F.; Delem, A. Co-administration study in South African infants of a live-attenuated oral human rotavirus vaccine (RIX4414) and poliovirus vaccines. Vaccine 2010, 28, 6542–6548. [Google Scholar] [CrossRef] [PubMed]
- Kaslow, D.C. Force of infection: A determinant of vaccine efficacy? NPJ Vaccines 2021, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.; Nagelkerke, N.; Heinsbroek, E.; Premkumar, P.S.; Wnęk, M.; Kang, G.; French, N.; Cunliffe, N.A.; Bar-Zeev, N.; Lopman, B.; et al. Estimating the incidence of rotavirus infection in children from India and Malawi from serial anti-rotavirus IgA titres. PLoS ONE 2017, 12, e0190256. [Google Scholar] [CrossRef] [PubMed]
- Armah, G.; Lewis, K.D.; Cortese, M.M.; Parashar, U.D.; Ansah, A.; Gazley, L.; Victor, J.C.; McNeal, M.M.; Binka, F.; Steele, A.D. A Randomized, Controlled Trial of the Impact of Alternative Dosing Schedules on the Immune Response to Human Rotavirus Vaccine in Rural Ghanaian Infants. J. Infect. Dis. 2016, 213, 1678–1685. [Google Scholar] [CrossRef] [Green Version]
- Bines, J.E.; Thobari., J.; Satria, C.D.; Handley, A.; Watts, E.; Cowley, D.; Nirwati, H.; Ackland, J.; Standish, J.; Justice, F.; et al. Human Neonatal Rotavirus Vaccine (RV3-BB) to Target Rotavirus from Birth. N. Engl. J. Med. 2018, 22, 719–730. [Google Scholar] [CrossRef]
- Mengelle, C.; Mansuy, J.M.; Prere, M.F.; Grouteau, E.; Claudet, I.; Kamar, N.; Huynh, A.; Plat, G.; Benard, M.; Marty, N.; et al. Simultaneous detection of gastrointestinal pathogens with a multiplex Luminex-based molecular assay in stool samples from diarrhoeic patients. Clin. Microbiol. Infect. 2013, 19, E458–E465. [Google Scholar] [CrossRef] [Green Version]
- Eibach, D.; Krumkamp, R.; Hahn, A.; Sarpong, N.; Adu-Sarkodie, Y.; Leva, A.; Käsmaier, J.; Panning, M.; May, J.; Tannich, E. Application of a multiplex PCR assay for the detection of gastrointestinal pathogens in a rural African setting. BMC Infect. Dis. 2016, 16, 150. [Google Scholar] [CrossRef] [Green Version]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef] [Green Version]
- Mokomane, M.; Tate, J.E.; Steenhoff, A.P.; Esona, M.D.; Bowen, M.D.; Lechiile, K.; Pernica, J.M.; Kasvosve, I.; Parashar, U.D.; Goldfarb, D.M. Evaluation of the Influence of Gastrointestinal Coinfections on Rotavirus Vaccine Effectiveness in Botswana. Pediatr. Infect. Dis. J. 2018, 37, e58–e62. [Google Scholar] [CrossRef] [Green Version]
- Praharaj, I.; Platts-Mills, J.A.; Taneja, S.; Antony, K.; Yuhas, K.; Flores, J.; Cho, I.; Bhandari, N.; Revathy, R.; Bavdekar, A.; et al. Diarrheal Etiology and Impact of Coinfections on Rotavirus Vaccine Efficacy Estimates in a Clinical Trial of a Monovalent Human-Bovine (116E) Oral Rotavirus Vaccine, Rotavac, India. Clin. Infect. Dis. 2019, 69, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Wallace, T.C.; Guarner, F.; Madsen, K.; Cabana, M.D.; Gibson, G.; Hentges, E.; Sanders, M.E. Human gut microbiota and its relationship to health and disease. Nutr. Rev. 2011, 69, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, J.L.; Backhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Huq, S.; Yatsunenko, T.; Haque, R.; Mahfuz, M.; Alam, M.A.; Benezra, A.; DeStefano, J.; Meier, M.F.; Muegge, B.D.; et al. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature 2014, 510, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Iturriza-Gómara, M.; Cunliffe, N.A. The Gut Microbiome as Possible Key to Understanding and Improving Rotavirus Vaccine Performance in High–Disease Burden Settings. J. Infect. Dis. 2017, 215, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Naylor, C.; Lu, M.; Haque, R.; Mondal, D.; Buonomo, E.; Nayak, U.; Mychaleckyj, J.C.; Kirkpatrick, B.; Colgate, R.; Carmolli, M.; et al. Environmental Enteropathy, Oral Vaccine Failure and Growth Faltering in Infants in Bangladesh. EBioMedicine 2015, 2, 1759–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, V.C.; Haak, B.W.; Handley, S.A.; Jiang, B.; Velasquez, D.E.; Hykes, B.L., Jr.; Droit, L.; Berbers, G.A.M.; Kemper, E.M.; van Leeuwen, E.M.M.; et al. Effect of Antibiotic-Mediated Microbiome Modulation on Rotavirus Vaccine Immunogenicity: A Human, Randomized-Control Proof-of-Concept Trial. Cell Host Microbe 2018, 24, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Center for Health Protection (CHP). Health Topics on Rotavirus Infection by the Department of Health, the Government of the Hong Kong Special Administrative Region. 5 July 2019. Available online: https://www.chp.gov.hk/en/healthtopics/content/24/38.html (accessed on 18 September 2020).
- UNICEF. Diarrhoea: Why Children Are Still Dying and What Can Be Done; UNICEF/WHO: New York, NY, USA, 2009. Available online: https://www.who.int/publications/i/item/9789241598415 (accessed on 18 September 2020).
- Shumetie, G.; Gedefaw, M.; Kebede, A.; Derso, T. Exclusive breastfeeding and rotavirus vaccination are associated with decreased diarrheal morbidity among under-five children in Bahir Dar, northwest Ethiopia. Public Health Rev. 2018, 39, 28. [Google Scholar] [CrossRef]
- Krawczyk, A.; Lewis, M.G.; Venkatesh, B.T.; Nair, S.N. Effect of Exclusive Breastfeeding on Rotavirus Infection among Children. Indian J. Pediatr. 2016, 83, 220–225. [Google Scholar] [CrossRef]
- Das, S.; Sahoo, G.C.; Das, P.; Singh, U.K.; Jaiswal, A.K.; Singh, P.; Kumar, R.; Kumar, R. Evaluating the Impact of Breastfeeding on Rotavirus Antigenemia and Disease Severity in Indian Children. PLoS ONE 2016, 11, e0146243. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Zhang, B.; Zhu, S.; Chen, J. No direct correlation between rotavirus diarrhea and breast feeding: A meta-analysis. Pediatrics Neonatol. 2018, 59, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Salim, H.; Karyana, I.P.; Sanjaya-Putra, I.G.; Budiarsa, S.; Soenarto, Y. Risk factors of rotavirus diarrhea in hospitalized children in Sanglah Hospital, Denpasar: A prospective cohort study. BMC Gastroenterol. 2014, 14, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Pei, J.; Qin, Z.; Wei, L. Efficacy of probiotics to prevent and/or alleviate childhood rotavirus infections. J. Funct. Foods 2019, 52, 90–99. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Makizaki, Y.; Oikawa, Y.; Tanaka, Y.; Maeda, A.; Shimakawa, M.; Komoto, S.; Moriguchi, K.; Ohno, H.; Taniguchi, K. Oral administration of Bifidobacterium bifidum G9-1 alleviates rotavirus gastroenteritis through regulation of intestinal homeostasis by inducing mucosal protective factors. PLoS ONE 2017, 12, e0173979. [Google Scholar] [CrossRef] [Green Version]
- Vlasova, A.N.; Kandasamy, S.; Chattha, K.S.; Rajashekara, G.; Saif, L.J. Comparison of probiotic lactobacilli and bifidobacteria effects, immune responses and rotavirus vaccines and infection in different host species. Vet. Immunol. Immunopathol. 2016, 172, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Lazarus, R.P.; John, J.; Shanmugasundaram, E.; Rajan, A.K.; Thiagarajan, S.; Giri, S.; Babji, S.; Sarkar; Kaliappan, P.S.; Venugopal, S. The effect of probiotics and zinc supplementation on the immune response to oral rotavirus vaccine: A randomized, factorial design, placebo-controlled study among Indian infants. Vaccine 2018, 36, 273–279. [Google Scholar] [CrossRef]
- Zeuthen, L.H.; Fink, L.N.; Frøkiaer, H. Toll-like receptor 2 and nucleotide-binding to ligomerization domain-2 play divergent roles in the recognition of gut-derived lactobacilli and bifidobacteria in dendritic cells. Immunology 2008, 124, 489–502. [Google Scholar] [CrossRef]
- Oksaharju, A.; Kankainen, M.; Kekkonen, R.A.; Lindstedt, K.A.; Kovanen, P.T.; Korpela, R. Probiotic Lactobacillus rhamnosus downregulates FCER1 and HRH4 expression in human mast cells. World J. Gastroenterol. 2011, 17, 750–759. [Google Scholar] [CrossRef]
- Ahmadi, E.; Alizadeh-Navaei, R.; Rezai, M.S. Efficacy of probiotic use in acute rotavirus diarrhea in children: A systematic review and meta-analysis. Casp. J. Int. Med. 2015, 6, 187–195. [Google Scholar]
- Park, M.S.; Kwon, B.; Ku, S.; Ji, G.E. The Efficacy of Bifidobacterium longum BORI and Lactobacillus acidophilus AD031 Probiotic Treatment in Infants with Rotavirus Infection. Nutrients 2017, 9, 887. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wanga, Y.; Lia, P.; Yina, Y.; Bijveldsa, M.J.; de Jongea, H.R.; Peppelenboscha, M.P.; Kainovb, D.E.; Pan, Q. Drug screening identifies gemcitabine inhibiting rotavirus through alteration of pyrimidine nucleotide synthesis pathway. Antivir. Res. 2020, 180, 104823. [Google Scholar] [CrossRef] [PubMed]
- Dycke, J.V.; Arnoldi, F.; Papa, G.; Vandepoele, J.; Burrone, O.R.; Mastrangelo, E.; Tarantino, D.; Heylen, E.; Neyts, J.; Rocha-Pereira, J. A Single Nucleoside Viral Polymerase Inhibitor against Norovirus, Rotavirus, and Sapovirus-Induced Diarrhea. J. Infect. Dis. 2018, 218, 1753–1758. [Google Scholar] [CrossRef] [PubMed]
- Knorr, J.I.E.; Erro, I.O.; Andueza, M.M.C.; Blecua, M.C.T.; Martinez, A.E.; Iparraguirre, A.I. Systematic review of the efficacy of racecadotril in the treatment of acute diarrhoea. An. Pediatr. 2008, 69, 432–438. [Google Scholar] [CrossRef] [Green Version]
- La-Frazia, S.; Ciucci, A.; Arnoldi, F.; Coira, M.; Gianferretti, P.; Angelini, M.; Belardo, G.; Burrone, O.R.; Rossignol, J.F.; Santoro, M.G. Thiazolides, a New Class of Antiviral Agents Effective against Rotavirus Infection, Target Viral Morphogenesis, Inhibiting Viroplasm Formation. J. Virol. 2013, 87, 11096–11106. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liu, L.; Chen, W.; Qin, F.; Zhou, F.; Yang, H. Ziyuglycoside II Inhibits Rotavirus Induced Diarrhea Possibly via TLR4/NF-κB Pathways. Biol. Pharm. Bull. 2020, 43, 932–937. [Google Scholar] [CrossRef]
- Siddique, I.; Khan, I. Mechanism of Regulation of Na-H Exchanger in Inflammatory Bowel Disease: Role of TLR-4 Signaling Mechanism. Dig. Dis. Sci. 2011, 56, 1656–1662. [Google Scholar] [CrossRef]
- Chen, S.; Ding, S.; Yin, Y.; Xu, L.; Li, P.; Peppelenbosch, M.P.; Pan, Q.; Wang, W. Suppression of pyrimidine biosynthesis by targeting DHODH enzyme robustly inhibits rotavirus replication. Antivir. Res. 2019, 167, 35–44. [Google Scholar] [CrossRef]
- Huang, H.; Liao, D.; Zhou, G.; Zhu, Z.; Cui, Y.; Pu, R. Antiviral activities of resveratrol against rotavirus in vitro and in vivo. Phytomedicine 2020, 77, 153230. [Google Scholar] [CrossRef]
- Gordon, M.; Akobeng, A. Racecadotril for acute diarrhoea in children: Systematic review and meta-analyses. Arch. Dis. Child. 2016, 101, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Mahapatro, S.; Mahilary, N.; Satapathy, A.K.; Das, R.R. Nitazoxanide in Acute Rotavirus Diarrhea: A Randomized Control Trial from a Developing Country. J. Trop. Med. 2017, 2017, 7942515. [Google Scholar] [CrossRef] [Green Version]
- Eichwald, C.; De Lorenzo, G.; Schraner, E.M.; Papa, G.; Bollati, M.; Swuec, P.; de Rosa, M.; Milani, M.; Mastrangelo, E.; Ackermann, M.; et al. Identification of a Small Molecule That Compromises the Structural Integrity of Viroplasms and Rotavirus Double-Layered Particles. J. Virol. 2018, 92, e01943-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Kim, K.; Kim, W. Genipin inhibits rotavirus-induced diarrhea by suppressing viral replication and regulating inflammatory responses. Sci. Rep. 2020, 10, 15836. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention (CDC). Rotavirus. Last Reviewed 23 April 2018. Available online: https://www.cdc.gov/rotavirus/index.html (accessed on 30 March 2020).




{kind=link}
{kind=link}
{kind=link}
| Genome Segment | Size (bp) | Number of Genotype | Genotype Denotation | Protein Product | Type of Protein/Location in Virion | Function |
|---|---|---|---|---|---|---|
| 1 | 3302 | 22 | R | VP1 | Structural, inner capsid | -RdRp -ss-RNA binding |
| 2 | 2687 | 20 | C | VP2 | Structural, core | -Houses RNA genome |
| 3 | 2592 | 20 | M | VP3 | Structrural, inner capsid | -guanyltransferase -methyltransferase -ss RNA binding |
| 4 | 2362 | 51 | P | VP4 | Structural, outer capsid | -receptor binding protein -infectivity enhancement through trysin cleavage |
| 5 | 1356 | 26 | I | VP6 | Structural, middle capsid | -Serological grouping and subgrouping antigen |
| 6 | 1062 | 36 | G | VP7 | Structural, outer capsid | -Neutralization antigen -Bases of binary classification |
| 7 | 1581 | 31 | A | NSP1 | Non-structural | -host interferon antagonist -anti-apoptosis |
| 8 | 1059 | 22 | N | NSP2 | Non-structural | -helicase -NTPase -NDPK -RBP |
| 9 | 1074 | 22 | T | NSP3 | Non-structural | -competition with host PABP for elf-4G1 binding -Translation enhancer |
| 10 | 751 | 27 | E | NSP4 | Non-structural | -enterotoxin -Transmembrane gp |
| 11 | 666 | 22 | H | NSP5 | Non-structural | -phosphoprotein |
| NSP6 | Non-structural | -ssRNA and dsRNA binding |
| Name | Licensing | Date of WHO Prequalification | Vaccine Antigens | Formulation | Storage Conditions | No. of Doses | Schedule | References |
|---|---|---|---|---|---|---|---|---|
| Rotarix (RV1; GSK) | Globally | March, 2009 | Live-attenuated, human wild-type G1P[8] strain [R1X4414] | Liquid | 2–8 °C for 36 months | 2 | 2 months and 4 months | [60,171] |
| RotaTeq (RV5; Merck) | Globally | October, 2008 | Live-attenuated, human-bovine rotavirus reassortant G1, G2, G3, G4 and P[8] | Liquid | 2–8 °C for 36 months | 3 | 2 months, 4 months and 6 months | [170] |
| Rotavac (Bharat) | Globally | January, 2018 | Live-attenuated wild-type reassortant G9P[11] strain [116E] | Liquid frozen | 2–8 °C for 7 months, −20 °C (long-term) | 3 | 6 weeks, 10 weeks and 14 weeks | [78] |
| Rotasiil (Serum institute) | Globally | September, 2018 | Live-attenuated human-bovine rotavirus reassortant G1, G2, G3, G4, and G9 | Lyophilized, Thermostable lyophilized & Liquid | <40 °C for 18 months <25 °C for 30 months 14 weeks | 3 | 6 weeks, 10 weeks and 14 weeks | [4,22] |
| Rotavin-M1 (POLYVAC) | Nationally | Not yet | Live-attenuated human rotavirus strain G1P[8] | Liquid frozen | 2–8 °C for 2 months −20 °C for 24 months | 2 | Minimum at 6 weeks, for 4 weeks apart | [4,22] |
| Lanzhou lamb (Lanzhou institute) | Nationally | Not yet | Live-attenuated lamb G10P[15] rotavirus strain | Liquid | 2–8 °C for 12 months | 4 | I dose annually for children aged 2–36 months | [4,22,60] |
| Name | Region | Efficacy (95% CI) | Reference |
|---|---|---|---|
| Rotarix | Europe | 96% (90–99%) | [182] |
| Latin America | 85% (72–92%) | [183] | |
| Africa | 62% (44–73%) | [184] | |
| Rotateq | Europe | 98% (88–100%) | [185] |
| Africa | 64% (40–79%) | [186] | |
| Asia | 51% (13–73%) | [187] | |
| Rotavac | Asia | 54% (37–70%) | [188] |
| Rotasiil | Africa | 67% (50–78%) | [189] |
| Rotasiil | Asia | 36% (12–54%) | [190] |
| Name | Mechanism | Reference |
|---|---|---|
| Gemcitabine | Pyrimidine nucleotide inhibitor | [242] |
| 2′-C-methyl nucleosides | Viral polymerase inhibitor | [243] |
| Racecadotril | Intestinal encephalinase inhibitor | [244] |
| Nitazoxanide | Inhibitor of viroplasm formation | [245] |
| Resveratrol | Inhibitor of viral protein synthesis | [249] |
| Ziyuglycoside II | Inhibitor of TLR4/NF-κB pathway | [246] |
| Brequinar | Pyrimidine biosynthesis inhibitor | [248] |
| ML-60218 | RNA polymerase III inhibitor | [252] |
| Genipin | Entry inhibitor | [253] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omatola, C.A.; Olaniran, A.O. Rotaviruses: From Pathogenesis to Disease Control—A Critical Review. Viruses 2022, 14, 875. https://doi.org/10.3390/v14050875
Omatola CA, Olaniran AO. Rotaviruses: From Pathogenesis to Disease Control—A Critical Review. Viruses. 2022; 14(5):875. https://doi.org/10.3390/v14050875
Chicago/Turabian StyleOmatola, Cornelius A., and Ademola O. Olaniran. 2022. "Rotaviruses: From Pathogenesis to Disease Control—A Critical Review" Viruses 14, no. 5: 875. https://doi.org/10.3390/v14050875