
Molecular Analysis of Caprine Enterovirus Circulating in China during 2016–2021: Evolutionary Significance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Viral Isolation
2.2. Viral Characterization
2.3. Immunoperoxidase Monolayer Assay (IPMA)
2.4. RNA Extraction, cDNA Synthesis, and PCR Amplification
2.5. Bioinformatics Analysis
3. Results
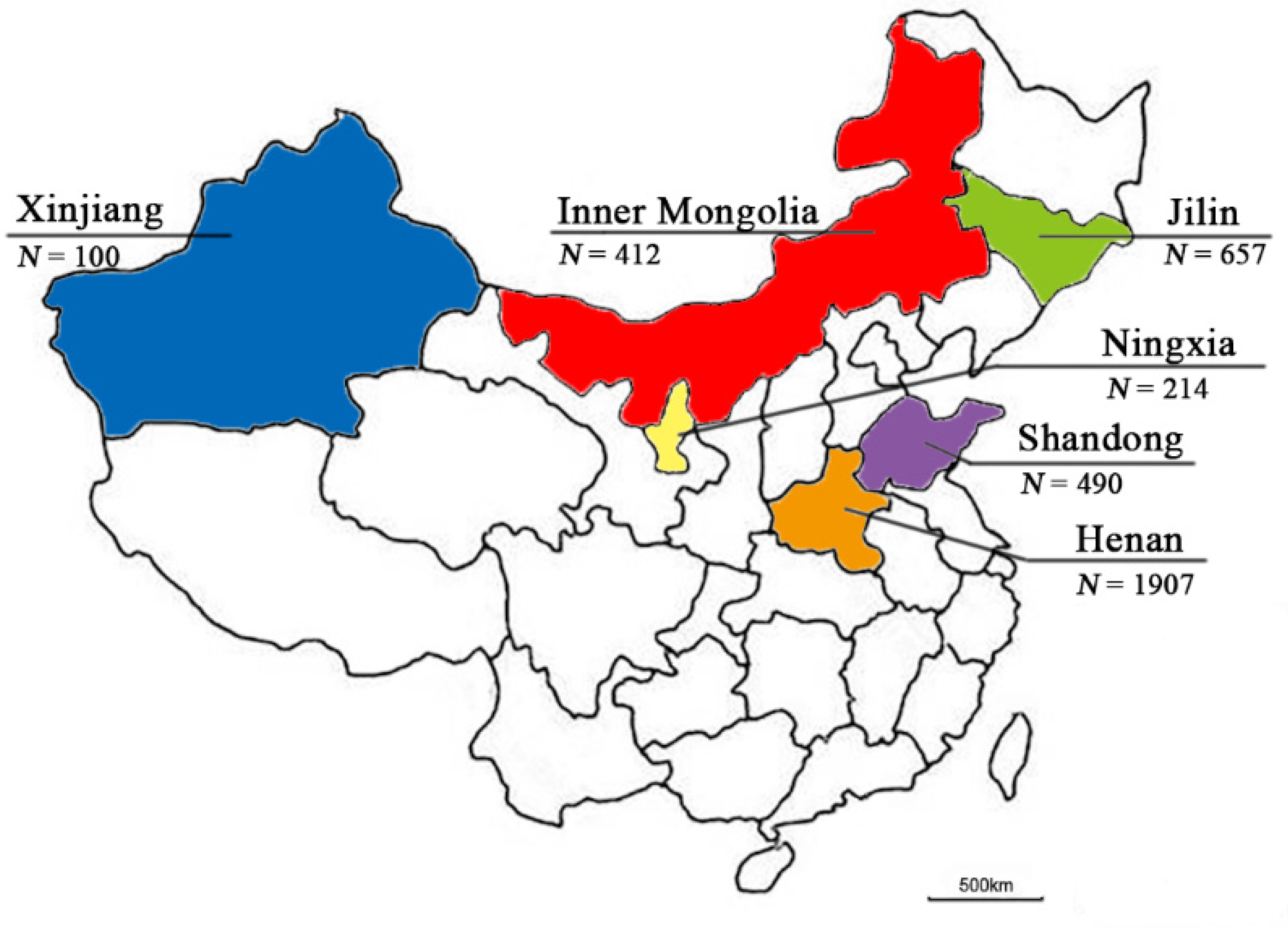
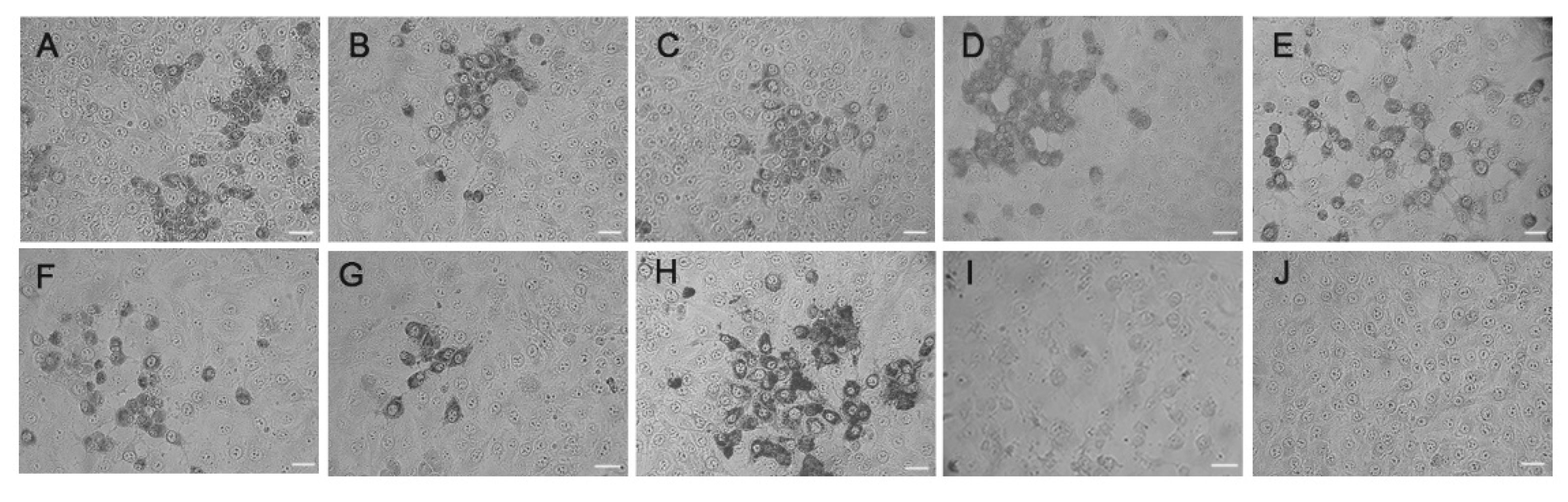
3.1. Isolation and Characterization of the Enterovirus Strains from Goat/Sheep Herds
3.2. Immune Cross-Reaction of the Novel Caprine Enterovirus Strains to CEV-JL14
3.3. Unveiling of the Complete Genome Sequences of Caprine/Ovine Enterovirus
3.4. High Sequence Identity Shared by These New Caprine/Ovine Enterovirus Strains with CEV-JL14
3.5. Novel Caprine/Ovine Enterovirus Strains Phylogenetically Similar to EV-G, and Three New EV-G Types Revealed
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Moll, T.; Ulrich, M.I. Biologic characteristics of certain bovine enteric viruses. Am. J. Vet. Res. 1963, 24, 545–550. [Google Scholar] [PubMed]
- Straub, O.C. Isolation of a bovine enterovirus from the genitalia. Dtsch. Tierarztl. Wochenschr. 1965, 72, 54–55. [Google Scholar] [PubMed]
- Blas-Machado, U.; Saliki, J.T.; Sanchez, S.; Brown, C.C.; Zhang, J.; Keys, D.; Woolums, A.; Harvey, S.B. Pathogenesis of a bovine enterovirus-1 isolate in experimentally infected calves. Vet. Pathol. 2011, 48, 1075–1084. [Google Scholar] [CrossRef]
- Li, Y.L.; Chang, J.T.; Wang, Q.; Yu, L. Isolation of two Chinese bovine enteroviruses and sequence analysis of their complete genomes. Arch. Virol. 2012, 157, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Blas-Machado, U.; Saliki, J.T.; Boileau, M.J.; Goens, S.D.; Caseltine, S.L.; Duffy, J.C.; Welsh, R.D. Fatal ulcerative and hemorrhagic typhlocolitis in a pregnant heifer associated with natural bovine enterovirus type-1 infection. Vet. Pathol. 2007, 44, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Xing, Z.; Gai, X.; Li, S.; San, Z.; Wang, X. Identification of a novel enterovirus E isolates HY12 from cattle with severe respiratory and enteric diseases. PLoS ONE 2014, 9, e97730. [Google Scholar] [CrossRef] [Green Version]
- Anbalagan, S.; Hesse, R.A.; Hause, B.M. First identification and characterization of porcine enterovirus G in the United States. PLoS ONE 2014, 9, e97517. [Google Scholar] [CrossRef]
- Donin, D.G.; de Arruda Leme, R.; Alfieri, A.F.; Alberton, G.C.; Alfieri, A.A. First report of Porcine teschovirus (PTV), Porcine sapelovirus (PSV) and Enterovirus G (EV-G) in pig herds of Brazil. Trop. Anim. Health Prod. 2014, 46, 523–528. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Y.; Shen, Q.; Zhang, W.; Hua, X. Prevalence of porcine enterovirus 9 in pigs in middle and eastern China. Virol. J. 2013, 10, 99. [Google Scholar] [CrossRef] [Green Version]
- Bunke, J.; Receveur, K.; Oeser, A.C.; Fickenscher, H.; Zell, R.; Krumbholz, A. High genetic diversity of porcine enterovirus G in Schleswig-Holstein, Germany. Arch. Virol. 2018, 163, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Boros, A.; Pankovics, P.; Knowles, N.J.; Reuter, G. Natural interspecies recombinant bovine/porcine enterovirus in sheep. J. Gen. Virol. 2012, 93, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, T.; Tsuchiaka, S.; Hirata, T.; Shiroma, Y.; Okazaki, S.; Katayama, Y.; Oba, M.; Nishiura, N.; Sassa, Y.; Furuya, T.; et al. Detection of enterovirus genome sequence from diarrheal feces of goat. Virus Genes 2014, 48, 550–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; He, J.; Lu, H.; Liu, Y.; Deng, Y.; Zhu, L.; Guo, C.; Tu, C.; Wang, X. A novel enterovirus species identified from severe diarrheal goats. PLoS ONE 2017, 12, e0174600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Income, N.; Kosoltanapiwat, N.; Taksinoros, S.; Leaungwutiwong, P.; Maneekan, P.; Chavez, I.F. Molecular Identification of Enteroviruses from Cattle and Goat Feces and Environment in Thailand. Appl. Environ. Microbiol. 2019, 85, e02420-18. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Lin, Q.; Hu, J.; Wang, X.; Li, X.; Cai, M.; Wang, W.; Zhang, Z.; Wang, X. Discovery of a virus of the species Enterovirus F in goats. Arch. Virol. 2019, 164, 2551–2558. [Google Scholar] [CrossRef]
- Pallansch, M.A.; Oberste, M.S.; Whitton, J.L. Enteroviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 490–530. [Google Scholar]
- Zell, R.; Sidigi, K.; Henke, A.; Schmidt-Brauns, J.; Hoey, E.; Martin, S.; Stelzner, A. Functional features of the bovine enterovirus 5’-non-translated region. J. Gen. Virol. 1999, 80, 2299–2309. [Google Scholar] [CrossRef]
- Zell, R.; Krumbholz, A.; Dauber, M.; Hoey, E.; Wutzler, P. Molecular-based reclassification of the bovine enteroviruses. J. Gen. Virol. 2006, 87, 375–385. [Google Scholar] [CrossRef]
- Tuthill, T.J.; Groppelli, E.; Hogle, J.M.; Rowlands, D.J. Picornaviruses. Curr. Top. Microbiol. Immunol. 2010, 343, 43–89. [Google Scholar] [CrossRef]
- Plevka, P.; Perera, R.; Cardosa, J.; Kuhn, R.J.; Rossmann, M.G. Crystal structure of human enterovirus 71. Science 2012, 336, 1274. [Google Scholar] [CrossRef] [Green Version]
- Cordey, S.; Petty, T.J.; Schibler, M.; Martinez, Y.; Gerlach, D.; van Belle, S.; Turin, L.; Zdobnov, E.; Kaiser, L.; Tapparel, C. Identification of site-specific adaptations conferring increased neural cell tropism during human enterovirus 71 infection. PLoS Pathog. 2012, 8, e1002826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmonds, P.; Gorbalenya, A.E.; Harvala, H.; Hovi, T.; Knowles, N.J.; Lindberg, A.M.; Oberste, M.S.; Palmenberg, A.C.; Reuter, G.; Skern, T.; et al. Recommendations for the nomenclature of enteroviruses and rhinoviruses. Arch. Virol. 2020, 165, 793–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Liu, D.; Liu, C.; Hu, J.; Hang, L.; Li, X.; Wei, Y.; Zhu, H.; Zhang, Q.; Wang, X. Construction and evaluation of HA-epitope-tag introduction onto the VP1 structural protein of a novel HY12 enterovirus. Virology 2018, 525, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Hu, J.; Zhang, Q.; Wang, Y.; Guo, C.; Zhu, L.; Tu, C.; Wang, X. Development of a sandwich ELISA for detection of CEV antigen and epidemiological investigation. Chin. J. Vet. Sci. 2017, 37, 653–657. [Google Scholar] [CrossRef]
- Earle, J.A.; Skuce, R.A.; Fleming, C.S.; Hoey, E.M.; Martin, S.J. The complete nucleotide sequence of a bovine enterovirus. J. Gen. Virol. 1988, 69, 253–263. [Google Scholar] [CrossRef]
- Oberste, M.S.; Maher, K.; Kilpatrick, D.R.; Flemister, M.R.; Brown, B.A.; Pallansch, M.A. Typing of human enteroviruses by partial sequencing of VP1. J. Clin. Microbiol. 1999, 37, 1288–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaukat, S.; Angez, M.; Alam, M.M.; Sharif, S.; Khurshid, A.; Malik, F.; Rana, M.S.; Mahmood, T.; Zaidi, S.S. Molecular identification and characterization of a new type of bovine enterovirus. Appl. Environ. Microbiol. 2012, 78, 4497–4500. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | Virus Strains | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 76.9 | 75.3 | 87.8 | 81.8 | 81.2 | 81.3 | 80.6 | 79.2 | 79.8 | 64.4 | 1 | CEV-JL14 | |
| 75.1 | 76.7 | 76.2 | 76.3 | 76.4 | 75.0 | 75.5 | 75.6 | 63.9 | 2 | TB4-OEV | ||
| 75.5 | 75.0 | 75.4 | 75.4 | 78.6 | 78.2 | 77.5 | 63.3 | 3 | 990/UK-NI | |||
| 79.9 | 79.2 | 79.3 | 78.3 | 78.3 | 77.7 | 64.5 | 4 | JL-LS174 | ||||
| 91.9 | 92.0 | 81.0 | 79.0 | 79.4 | 64.5 | 5 | JL-LS127 | |||||
| 98.8 | 80.3 | 78.3 | 78.7 | 64.4 | 6 | JL-LS165 | ||||||
| 80.4 | 78.3 | 78.7 | 64.3 | 7 | JL-LS34 | |||||||
| 83.5 | 82.5 | 64.6 | 8 | NMG-F37 | ||||||||
| 86.0 | 64.8 | 9 | NX-DR26 | |||||||||
| 64.5 | 10 | SD-S68 | ||||||||||
| 11 | SD-S67 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | Virus Strains | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 68.7 | 61.7 | 95.8 | 67.3 | 67.3 | 66.9 | 64.2 | 66.3 | 65.6 | 52.0 | 1 | CEV-JL14 | |
| 61.0 | 69.7 | 74.3 | 73.6 | 73.9 | 62.8 | 66.3 | 65.2 | 52.7 | 2 | TB4-OEV | ||
| 60.6 | 59.9 | 59.9 | 59.3 | 77.4 | 75.3 | 74.9 | 50.9 | 3 | 990/UK-NI | |||
| 66.9 | 66.9 | 66.5 | 64.2 | 67.0 | 66.3 | 51.6 | 4 | JL-LS174 | ||||
| 99.3 | 98.6 | 61.3 | 62.1 | 61.7 | 49.1 | 5 | JL-LS127 | |||||
| 97.6 | 61.3 | 62.1 | 61.7 | 48.7 | 6 | JL-LS165 | ||||||
| 60.6 | 61.3 | 61.0 | 48.7 | 7 | JL-LS34 | |||||||
| 78.8 | 79.2 | 52.0 | 8 | NMG-F37 | ||||||||
| 95.1 | 53.5 | 9 | NX-DR26 | |||||||||
| 52.0 | 10 | SD-S68 | ||||||||||
| 11 | SD-S67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, X.; Lin, Q.; Zhang, Q.; Hu, J.; Tursun, G.; Deng, Y.; Guo, C.; Wang, X. Molecular Analysis of Caprine Enterovirus Circulating in China during 2016–2021: Evolutionary Significance. Viruses 2022, 14, 1051. https://doi.org/10.3390/v14051051
Chang X, Lin Q, Zhang Q, Hu J, Tursun G, Deng Y, Guo C, Wang X. Molecular Analysis of Caprine Enterovirus Circulating in China during 2016–2021: Evolutionary Significance. Viruses. 2022; 14(5):1051. https://doi.org/10.3390/v14051051
Chicago/Turabian StyleChang, Xiaoran, Qian Lin, Qun Zhang, Junying Hu, Gulbahar Tursun, Yingrui Deng, Chunguang Guo, and Xinping Wang. 2022. "Molecular Analysis of Caprine Enterovirus Circulating in China during 2016–2021: Evolutionary Significance" Viruses 14, no. 5: 1051. https://doi.org/10.3390/v14051051