
High Level of Pre-Treatment HIV-1 Drug Resistance and Its Association with HLA Class I-Mediated Restriction in the Pumwani Sex Worker Cohort

Abstract
:1. Introduction
2. Methods
2.1. Study Participants and Samples
2.2. Amplification and Sequencing of HIV Genome
2.3. HLA Genotyping
2.4. Sequence Analysis
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Study Participants
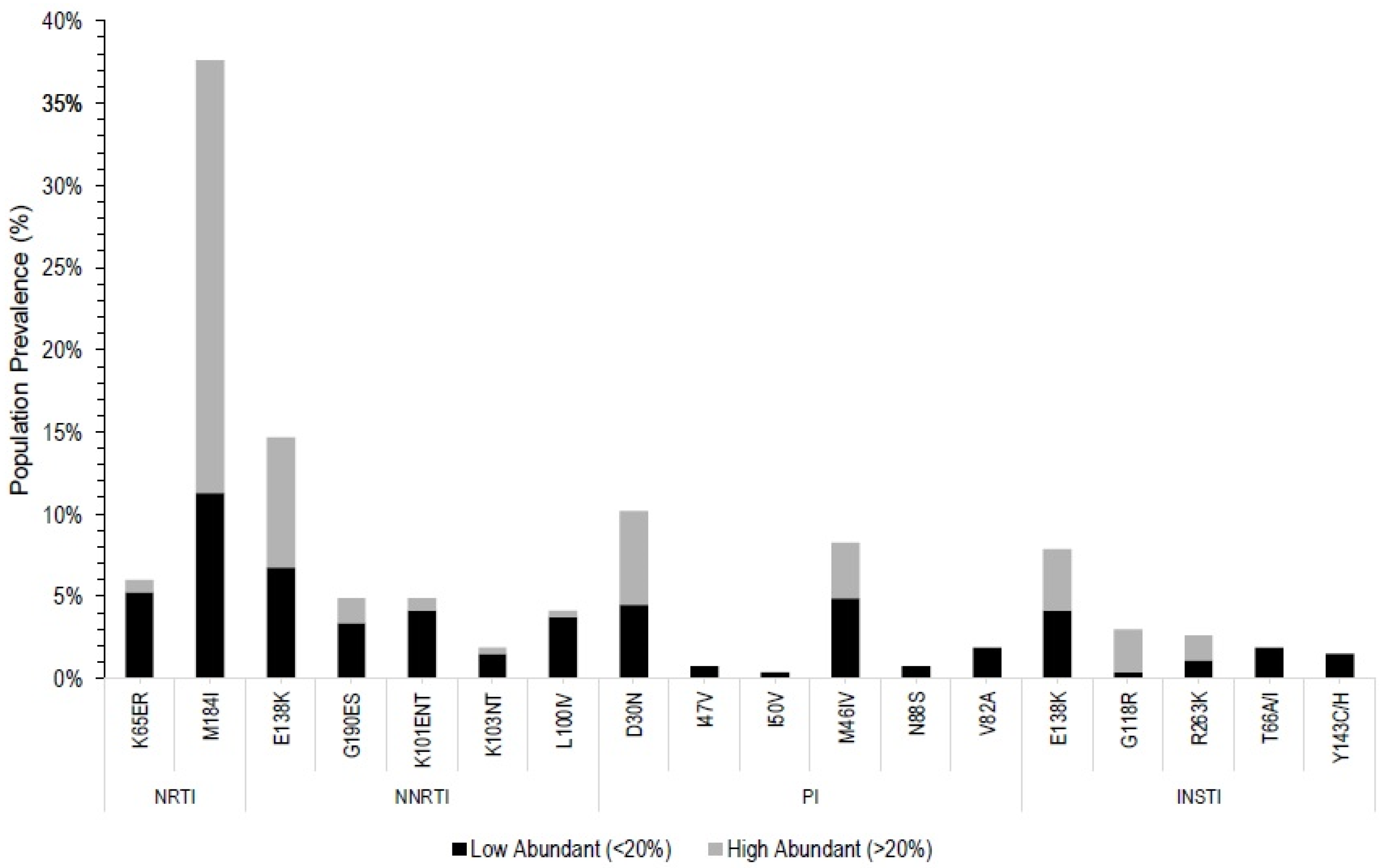
3.2. Pre-ART Drug Resistance Mutations
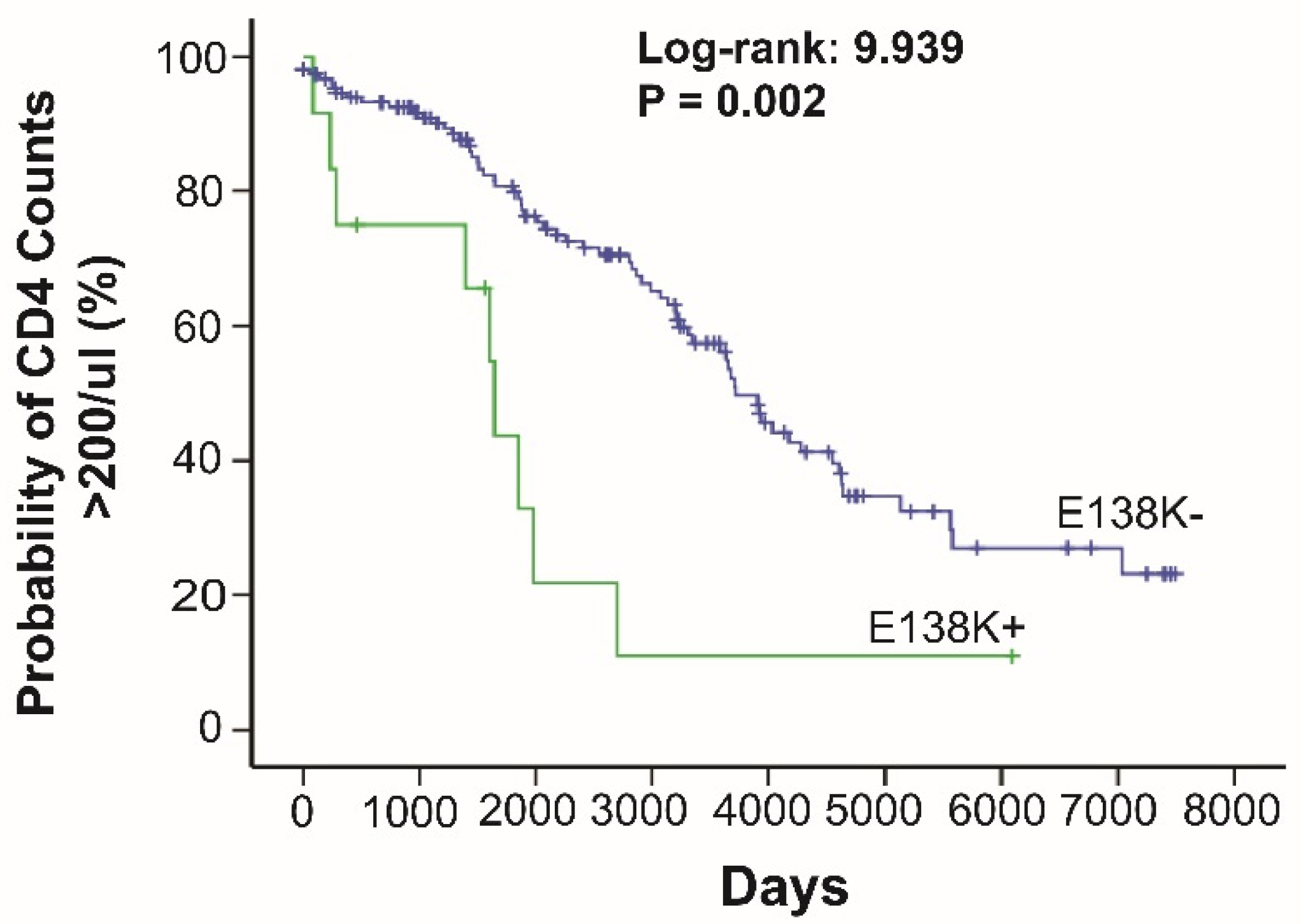
3.3. Pre-ART Drug Resistance Associated with HLA Class I Alleles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Global HIV/AIDS Epidemic, HIV.gov. 2021. Available online: https://www.hiv.gov/hiv-basics/overview/data-and-trends/global-statistics (accessed on 29 June 2021).
- UNAIDS, Update. 2020. Switzerland. Available online: Https://www.unaids.org/en/resources/documents/2020/global-aids-report (accessed on 29 June 2021).
- AIDS and the Sustainable Development Goals, UNAIDS. Switzerland. 2021. Available online: Https://www.unaids.org/en/AIDS_SDGs (accessed on 29 June 2021).
- Fast-Track–Strategy to End the AIDS Epidemic by 2030, UNAIDS. 2014. Switerland. Available online: Https://www.unaids.org/en/resources/documents/2014/JC2686_WAD2014report (accessed on 29 June 2021).
- Eisinger, R.W.; Dieffenbach, C.W.; Fauci, A.S. HIV viral load and transmissibility of HIV infection. JAMA 2019, 321, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Chen, Y.Q.; McCauley, M.; Gamble, T.; Hosseinipour, M.C.; Kumarasamy, N.; Hakim, J.G.; Kumwenda, J.; Grinsztejn, B.; Pilotto, J.H.; et al. Prevention of HIV-1 infection with early antiretroviral therapy. N. Engl. J. Med. 2011, 365, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Rodger, A.J.; Cambiano, V.; Bruun, T.; Vernazza, P.; Collins, S.; Van Lunzen, J.; Corbelli, G.M.; Estrada, V.; Geretti, A.M.; Beloukas, A.; et al. Sexual activity without condoms and risk of HIV transmission in serodifferent couples when the HIV-positive partner is using suppressive antiretroviral therapy. JAMA 2016, 316, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Bavinton, B.R.; Pinto, A.N.; Phanuphak, N.; Grinsztejn, B.; Prestage, G.; Zablotska-Manos, I.B.; Jin, F.; Fairley, C.K.; Moore, R.; Roth, N.; et al. Viral suppression and HIV transmission in serodiscordant male couples: An international, prospective, observational, cohort study. Lancet HIV 2018, 5, e438–e447. [Google Scholar] [CrossRef]
- Clutter, D.S.; Jordan, M.R.; Bertagnolio, S.; Shafer, R.W. HIV-1 drug resistance and resistance testing. Infect. Genet. Evol. 2016, 46, 292–307. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. HIV Drug Resistance. 2021. Switzerland. Available online: Https://www.who.int/publications/i/item/9789240039608 (accessed on 29 June 2021).
- Fanales-Belasio, E.; Raimondo, M.; Suligoi, B.; Butto, S. HIV virology and pathogenetic mechanisms of action: A brief overview. Ann. Ist. Super. Sanità 2010, 46, 5–14. [Google Scholar] [CrossRef]
- Brumme, Z.L.; Kinloch, N.N.; Sanche, S.; Wong, A.; Martin, E.; Cobarrubias, K.D.; Sandstrom, P.; Levett, P.N.; Harrigan, P.R.; Joy, J.B. Extensive host immune adaptation in a concentrated North American HIV epidemic. AIDS 2018, 32, 1927–1938. [Google Scholar] [CrossRef]
- Gatanaga, H.; Murakoshi, H.; Hachiya, A.; Hayashida, T.; Chikata, T.; Ode, H.; Tsuchiya, K.; Sugiura, W.; Takiguchi, M.; Oka, S. Naturally selected rilpivirine-resistant HIV-1 variants by host cellular immunity. Clin. Infect. Dis. 2013, 57, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.; Muenchhoff, M.; Mann, J.; Roberts, H.E.; Matthews, P.; Adland, E.; Hempenstall, A.; Huang, K.-H.; Brockman, M.; Brumme, Z.; et al. Impact of HLA-driven HIV adaptation on virulence in populations of high HIV seroprevalence. Proc. Natl. Acad. Sci. USA 2014, 111, E5393–E5400. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.B.; John, M.; James, I.R.; Christiansen, F.T.; Witt, C.S.; Mallal, S.A. Evidence of HIV-1 adaptation to HLA-restricted immune responses at a population level. Science 2002, 296, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- Brumme, Z.; John, M.; Carlson, J.M.; Brumme, C.; Chan, D.; Brockman, M.; Swenson, L.; Tao, I.; Szeto, S.; Rosato, P.; et al. HLA-Associated Immune Escape Pathways in HIV-1 Subtype B Gag, Pol and Nef Proteins. PLoS ONE 2009, 4, e6687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.K.; Gregson, J.; Parkin, N.; Haile-Selassie, H.; Tanuri, A.; Forero, L.A.; Kaleebu, P.; Watera, C.; Aghokeng, A.; Mutenda, N.; et al. HIV-1 drug resistance before initiation or re-initiation of first-line antiretroviral therapy in low-income and middle-income countries: A systematic review and meta-regression analysis. Lancet Infect. Dis. 2018, 18, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Murray, M.; Sampathkumar, R.; Luo, M. HIV-1 genotypic drug resistance testing and Next-generation sequencing. Front. Clin. Drug Res.–HIV 2020, 5, 1–58. [Google Scholar]
- WHO. Update of Recommendations on First- and Second-Line Antiretroviral Regimens; World Health Organization: Geneva, Switzerland, 2019; p. 3. [Google Scholar]
- Mbunkah, H.A.; Bertagnolio, S.; Hamers, R.L.; Hunt, G.; Inzaule, S.; De Wit, T.F.R.; Paredes, R.; Parkin, N.T.; Jordan, M.R.; Metzner, K.J.; et al. Low-Abundance Drug-Resistant HIV-1 Variants in Antiretroviral Drug-Naive Individuals: A Systematic Review of Detection Methods, Prevalence, and Clinical Impact. J. Infect. Dis. 2020, 221, 1584–1597. [Google Scholar] [CrossRef] [PubMed]
- Land, A.M.; Luo, M.; Pilon, R.; Sandstrom, P.; Embree, J.; Wachihi, C.; Kimani, J.; Plummer, F.A.; Ball, T.B. High Prevalence of Genetically Similar HIV-1 Recombinants among Infected Sex Workers in Nairobi, Kenya. AIDS Res. Hum. Retrovir. 2008, 24, 1455–1460. [Google Scholar] [CrossRef]
- Luo, M.; Blanchard, J.; Brunham, K.; Pan, Y.; Shen, C.X.; Lu, H.; Brunham, R.C. Two-Step High Resolution Sequence-Based HLA-DRB Typing of Exon 2 DNA With Taxonomy-Based Sequence Analysis Allele Assignment. Hum. Immunol. 2001, 62, 1294–1310. [Google Scholar] [CrossRef]
- Land, A.M.; Ball, T.B.; Luo, M.; Rutherford, J.; Sarna, C.; Wachihi, C.; Kimani, J.; Plummer, F.A. Full-length HIV type 1 Proviral Sequencing of 10 Highly Exposed Women from Nairobi, Kenya Reveals a High Proportion of Intersubtype Recombinants. AIDS Res. Hum. Retrovir. 2008, 24, 865–872. [Google Scholar] [CrossRef]
- Luo, M.; Blanchard, J.; Pan, Y.; Brunham, K.; Brunham, R.C. High-resolution sequence typing of HLA-DQA1 and -DQB1 exon 2 DNA with taxonomy-based sequence analysis (TBSA) allele assignment. Tissue Antigens 1999, 54, 69–82. [Google Scholar] [CrossRef]
- Liu, T.F.; Shafer, R.W. Web Resources for HIV Type 1 Genotypic-Resistance Test Interpretation. Clin. Infect. Dis. 2006, 42, 1608–1618. [Google Scholar] [CrossRef] [Green Version]
- Wensing, A.M.; Calvez, V.; Ceccherini-Silberstein, F.; Charpentier, C.; Günthard, H.F.; Paredes, R.; Shafer, R.W.; Richman, D.D. 2019 update of the drug resistance mutations in HIV-1. Top. Antivir. Med. 2019, 27, 111–121. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Macintosh; Version 27; IBM Corp.: Armonk, NY, USA, 2020. [Google Scholar]
- National AIDS and STI Control Programme. Guidelines on Use of Antiretroviral Drugs for Treating and Preventing HIV Infection in Kenya; Nascop: Nairobi, Kenya, 2018. [Google Scholar]
- Hassan, A.S.; Esbjörnsson, J.; Wahome, E.; Thiong’o, A.; Makau, G.N.; Price, M.A.; Sanders, E.J. HIV-1 subtype diversity, transmission networks and transmitted drug resistance amongst acute and early infected MSM populations from Coastal Kenya. PLoS ONE 2018, 13, e0206177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatanaga, H.; Brumme, Z.L.; Adland, E.; Reyes-Terán, G.; Avila-Rios, S.; Mejía-Villatoro, C.R.; Hayashida, T.; Chikata, T.; van Tran, G.; van Nguyen, K.; et al. Potential for immune-driven viral polymorphisms to compromise antiretroviral-based pre-exposure prophylaxis for prevention of HIV-1 infection. AIDS 2009, 31, 1935–1943. [Google Scholar] [CrossRef] [PubMed]
- Crowell, T.A.; Danboise, B.; Parikh, A.; Esber, A.; Dear, N.; Coakley, P.; Kasembeli, A.; Maswai, J.; Khamadi, S.; Bahemana, E.; et al. Pretreatment and Acquired Antiretroviral Drug Resistance Among Persons Living With HIV in Four African Countries. Clin. Infect. Dis. 2020, 73, e2311–e2322. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.S.; Bibby, D.F.; Mwaringa, S.M.; Agutu, C.A.; Ndirangu, K.K.; Sanders, E.J.; Cane, P.A.; Mbisa, J.; Berkley, J.A. Presence, persistence and effects of pre-treatment HIV-1 drug resistance variants detected using next generation sequencing: A Retrospective longitudinal study from rural coastal Kenya. PLoS ONE 2019, 14, E0210559. [Google Scholar] [CrossRef]
- Crowell, T.A.; Kijak, G.H.; Sanders-Buell, E.; O’sullivan, A.M.; Kokogho, A.; Parker, Z.F.; Lawlor, J.; Polyak, C.S.; Adebajo, S.; Nowak, R.G.; et al. Transmitted, pre-treatment and acquired antiretroviral drug resistance among men who have sex with men and transgender women living with HIV in Nigeria. Antivir. Ther. 2019, 24, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Paredes, R.; Ribaudo, H.J.; Svarovskaia, E.S.; Metzner, K.J.; Kozal, M.J.; Hullsiek, K.H.; Balduin, M.; Jakobsen, M.R.; Geretti, A.M.; et al. Low-frequency HIV-1 drug resistance mutations and risk of NNRTI-based antiretroviral treatment failure: A systematic review and pooled analysis. JAMA-J. Am. Med. Assoc. 2011, 305, 1327–1335. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Z.; Paredes, R.; Ribaudo, H.J.; Svarovskaia, E.S.; Kozal, M.; Hullsiek, K.H.; Miller, M.D.; Bangsberg, D.R.; Kuritzkes, D.R. Relationship between minority nonnucleoside reverse transcriptase inhibitor resistance mutations, adherence, and the risk of virologic failure. AIDS 2012, 26, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Beck, I.A.; Levine, M.; McGrath, C.J.; Bii, S.; Milne, R.S.; Kingoo, J.M.; So, I.; Andersen, N.; Dross, S.; Coombs, R.W.; et al. Pre-treatment HIV-drug resistance associated with virologic outcome of first-line NNRTI-antiretroviral therapy: A cohort study in Kenya. EClinicalMedicine 2020, 18, 100239. [Google Scholar] [CrossRef]
- Onywera, H.; Maman, D.; Inzaule, S.; Auma, E.; Were, K.; Fredrick, H.; Owiti, P.; Opollo, V.; Etard, J.-F.; Mukui, I.; et al. Surveillance of HIV-1 pol transmitted drug resistance in acutely and recently infected antiretroviral drug-naïve persons in rural western Kenya. PLoS ONE 2017, 12, e0171124. [Google Scholar] [CrossRef] [Green Version]
- Makwaga, O.; Adhiambo, M.; Mulama, D.H.; Muoma, J.; Adungo, F.; Wanjiku, H.; Ongaya, A.; Maitha, G.M.; Mwau, M. Prevalence of human immunodeficiency virus-1 drug-resistant mutations among adults on first- and second-line antiretroviral therapy in a resource-limited health facility in Busia County, Kenya. Pan Afr. Med. J. 2020, 37, 311. [Google Scholar] [CrossRef]
- Kasang, C.; Kalluvya, S.; Majinge, C.; Stich, A.; Bodem, J.; Kongola, G.; Jacobs, G.B.; Mlewa, M.; Mildner, M.; Hensel, I.; et al. HIV drug resistance (HIVDR) in antiretroviral therapy-naïve patients in Tanzania not eligible for WHO threshold HIVDR survey is dramatically high. PLoS ONE 2011, 6, e23091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, R.; Babaoglu, K.; Lansdon, E.B.; Rimsky, L.; van Eygen, V.; Picchio, G.; Svarovskaia, E.; Miller, M.D.; White, K.L. The HIV-1 Reverse Transcriptase M184I Mutation Enhances the E138K-Associated Resistance to Rilpivirine and Decreases Viral Fitness. J. Acquir. Immune Defic. Syndr. 2012, 59, 47–54. Available online: https://www.jaids.com (accessed on 17 July 2021). [CrossRef] [PubMed] [Green Version]
- Carlson, J.M.; Du, V.Y.; Pfeifer, N.; Bansal, A.; Tan, V.Y.; Power, K.; Brumme, C.; Kreimer, A.; DeZiel, C.E.; Fusi, N.; et al. Impact of pre-Adapted HIV transmission. Nat. Med. 2017, 22, 606–613. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, S.M.; Kamelian, K.; Musinguzi, N.; Kigozi, S.; Boum, Y.; Bwana, M.B.; Muzoora, C.; Brumme, Z.L.; Carrington, M.; Carlson, J.; et al. Pre-treatment integrase inhibitor resistance is uncommon in antiretroviral therapy-naive individuals with HIV-1 subtype A1 and D infections in Uganda. AIDS 2021, 35, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.M.; Spriewald, B.M.; Bergmann, S.; Eismann, K.; Leykauf, M.; Korn, K.; Walter, H.; Schmidt, B.; Arnold, M.-L.; Harrer, E.G.; et al. Influence of Major HIV-1 Protease Inhibitor Resistance Mutations on CTL Recognition. Available online: https://journals.lww.com/jaids (accessed on 20 July 2021).



{kind=link}
{kind=link}
| Parameters | Median ± IQR * | n | % |
|---|---|---|---|
| Age (yrs) | 35 ± 10 | 252 | 94.7 |
| 18–29 | 59 | 22.2 | |
| 30–39 | 127 | 47.7 | |
| 40+ | 66 | 24.8 | |
| Unknown | 14 | 5.3 | |
| Country of origin | 266 | 100 | |
| Kenya | 126 | 47.4 | |
| Tanzania | 123 | 46.2 | |
| Uganda | 14 | 5.3 | |
| unknown | 3 | 1.1 | |
| HIV Subtype | 266 | 100 | |
| A1 | 150 | 56.4 | |
| B | 1 | 0.4 | |
| C | 14 | 5.3 | |
| D | 25 | 9.4 | |
| G | 1 | 0.4 | |
| AD | 35 | 13.2 | |
| Other CRF | 35 | 13.2 | |
| unknown | 5 | 1.9 | |
| CD4+ T-cell count, cells/μL | 259 ± 218.75 | 65 | 24.4 |
| Duration of HIV-1 Positive (yrs) | 7.67 ± 8.93 | 260 | 97.7 |
| Level of ARV Drug Resistance Conferred by Mutation | DRM Represented in Databases | ||||||
|---|---|---|---|---|---|---|---|
| Drug Class | Mutations (% T a, D b, L c) | High-Level Resistance | Intermediate Level Resistance | Low-Level Resistance | IAS-USA | Stanford HIV Db | WHO Surveillance Mutation |
| NRTI | M184I * (37, 26, 11) | FTC, 3TC | ✓ | ✓ | ✓ | ||
| K65ER * (6, 1, 5) | TDF | ABC, FTC, 3TC | ✓ | ✓ | ✓ | ||
| NNRTI | E138AK (15, 8, 7) | RPV | ✓ | ✓ | |||
| K101EN (5, 1,4) | NVP, RPV | DOR, EFV, ETR | ✓ | ✓ | ✓ | ||
| G190ES (5, 2, 3) | DOR, EFV, NVP, RPV | ETR, DOR | RPV | ✓ | ✓ | ✓ | |
| L100IV (4, 0.4, 3.6) | EFV, NVP | RPV | ✓ | ✓ | ✓ | ||
| K103NT (2, 0.5, 1.5) | EFV, NVP | ✓ | ✓ | ✓ | |||
| PI | D30N (10, 6, 4) | NFV ‡ | ✓ | ✓ | ✓ | ||
| M46I (8, 3, 5) | ATV/R, LPV/r | ✓ | ✓ | ✓ | |||
| V82A (2, 0, 2) | LPV/r | ATV/r | ✓ | ✓ | ✓ | ||
| N88S (1, 0, 1) | ATV/r | ✓ | ✓ | ✓ | |||
| I47V (1, 0, 1) | LPV/r, DRV/r, ATV/r | ✓ | ✓ | ✓ | |||
| I50V (0.5, 0, 0.5) | FPV | LPV/R | DRV/r | ✓ | ✓ | ✓ | |
| INSTI | E138K † (8, 4, 4) | EVG, RAL, DTG, CAB, BIC | ✓ | ✓ | |||
| G118R (3, 2.6, 0.4) | CAB, EVG, RAL | DTG, BIC | ✓ | ✓ | |||
| R263K (3, 1, 2) | DTG, EVG, CAB, BIC | RAL | ✓ | ✓ | |||
| T66IA (2, 0, 2) | EVG | RAL | ✓ | ✓ | |||
| Y143CH (1.5, 0, 1.5) | RAL | ✓ | ✓ | ||||
| DRM ^ (POPULATION PREVALENCE (% T A, D B, L C) | HLA & ALLELE (HLA−DRM-, HLA+ DRM-, HLA- DRM+, HLA+ DRM+) | p-Value | Adjusted p-Value # | Allele Frequency (>10%) | Allele Frequency (5–10%) |
|---|---|---|---|---|---|
| RT % E138K (11, 6, 5) RT G190ES (5, 2, 3) | A*66:01 (196,19,214,25) | 0.038 | 1.000 | Kenya, Uganda | |
| B*45:01 (202,15,18,7) | 0.003 | 0.928 | Rwanda, Uganda | Zimbabwe, Kenya, Cameroon, South African Black | |
| A*68:02 (201,29,3,4) | 0.008 | 1.000 | Kenya | Zimbabwe Ghana, South Africa, Uganda | |
| PI § M46I (8, 3, 5) | C*04:05 (201,14,15,4) | 0.046 | 1.000 | ||
| C*04:07 (208,7,16,3) | 0.038 | 1.000 | |||
| C*17:01 (171,44,10,9) | 0.018 | 1.000 | Ghana, Kenya, South Africa Black | Zimbabwe, Uganda, South Africa | |
| IN $ T97A (12, 9, 3) | A*66:01 (200,13,14,12) | 6.20 × 10−7 | 0.001 | Kenya, Uganda | |
| IN E138K (8, 4, 4) | A*68:01 (220,4,13,2) | 0.048 | 1.000 | Senegal | |
| A*68:02 (194,28,10,5) | 0.042 | 1.000 | Kenya | Zimbabwe Ghana, South Africa, Uganda | |
| B*15:17 (224,3,13,2) | 0.032 | 1.000 | |||
| B*35:02 (223,4,13,2) | 0.043 | 1.000 | |||
| C*17:01 (173,46,8,7) | 0.021 | 1.000 | Ghana, Kenya, South African Black | Kenya, Zimbabwe, Uganda, South Africa | |
| IN R263K (3, 1, 2) | A*30:04 (225,8,4,2) | 0.022 | 1.000 | ||
| B*15:03 (200,36,2,4) | 0.011 | 1.000 | Kenya, Uganda, Zimbabwe, Senegal, South Africa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willim, R.; Shadabi, E.; Sampathkumar, R.; Li, L.; Balshaw, R.; Kimani, J.; Plummer, F.A.; Luo, M.; Liang, B. High Level of Pre-Treatment HIV-1 Drug Resistance and Its Association with HLA Class I-Mediated Restriction in the Pumwani Sex Worker Cohort. Viruses 2022, 14, 273. https://doi.org/10.3390/v14020273
Willim R, Shadabi E, Sampathkumar R, Li L, Balshaw R, Kimani J, Plummer FA, Luo M, Liang B. High Level of Pre-Treatment HIV-1 Drug Resistance and Its Association with HLA Class I-Mediated Restriction in the Pumwani Sex Worker Cohort. Viruses. 2022; 14(2):273. https://doi.org/10.3390/v14020273
Chicago/Turabian StyleWillim, Rachel, Elnaz Shadabi, Raghavan Sampathkumar, Lin Li, Robert Balshaw, Joshua Kimani, Francis A. Plummer, Ma Luo, and Binhua Liang. 2022. "High Level of Pre-Treatment HIV-1 Drug Resistance and Its Association with HLA Class I-Mediated Restriction in the Pumwani Sex Worker Cohort" Viruses 14, no. 2: 273. https://doi.org/10.3390/v14020273