
The Secret Life of the Inhibitor of Virus Replication
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Two-Hybrid Assays
2.2. Comparisons of DNA and Protein Sequences
2.3. Protein Structure Modeling by AlphaFold
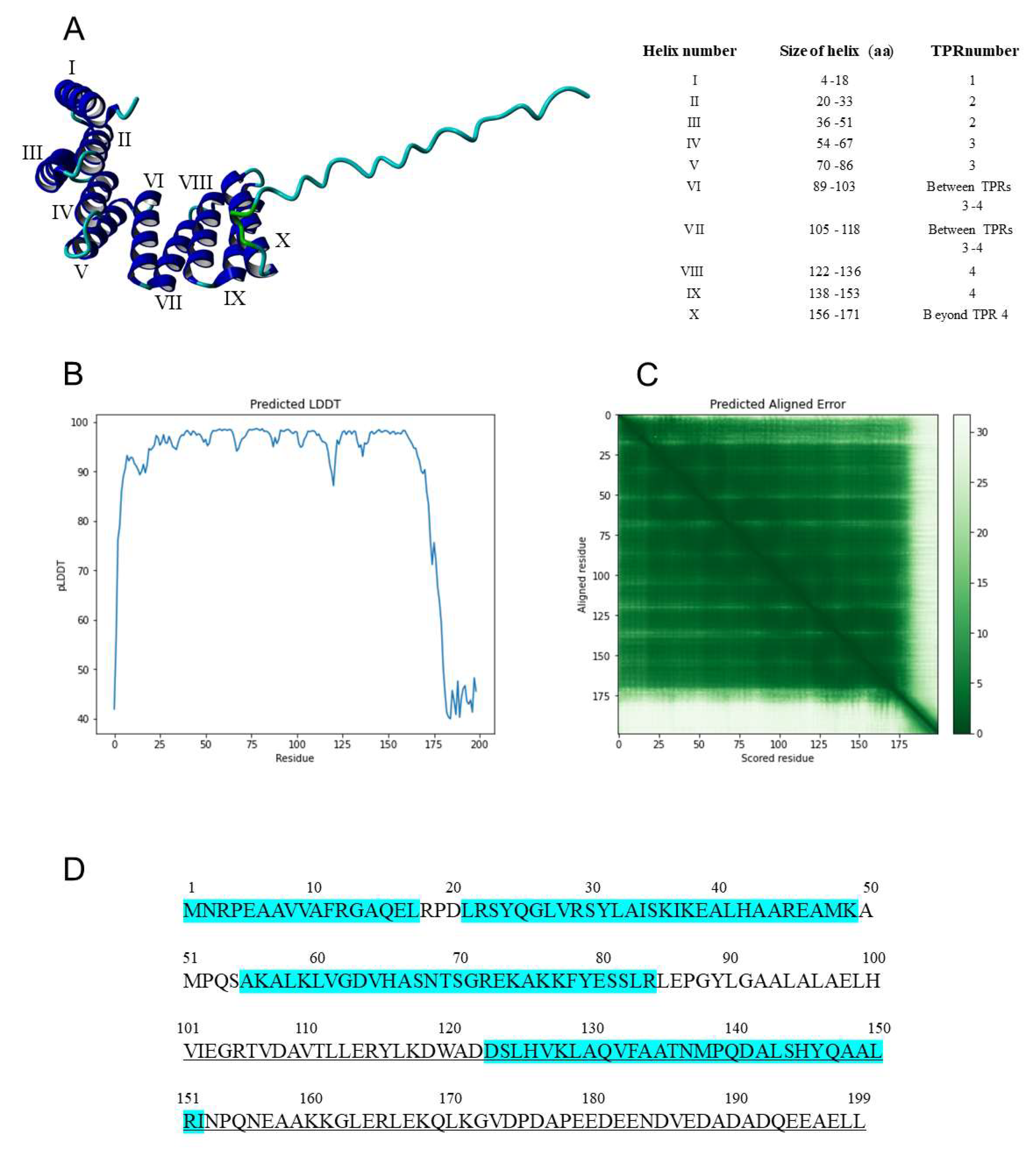
2.4. Identifying TPR Elements
3. Results
3.1. IVR1 and MYB1 Interact with the CMV 1a Protein
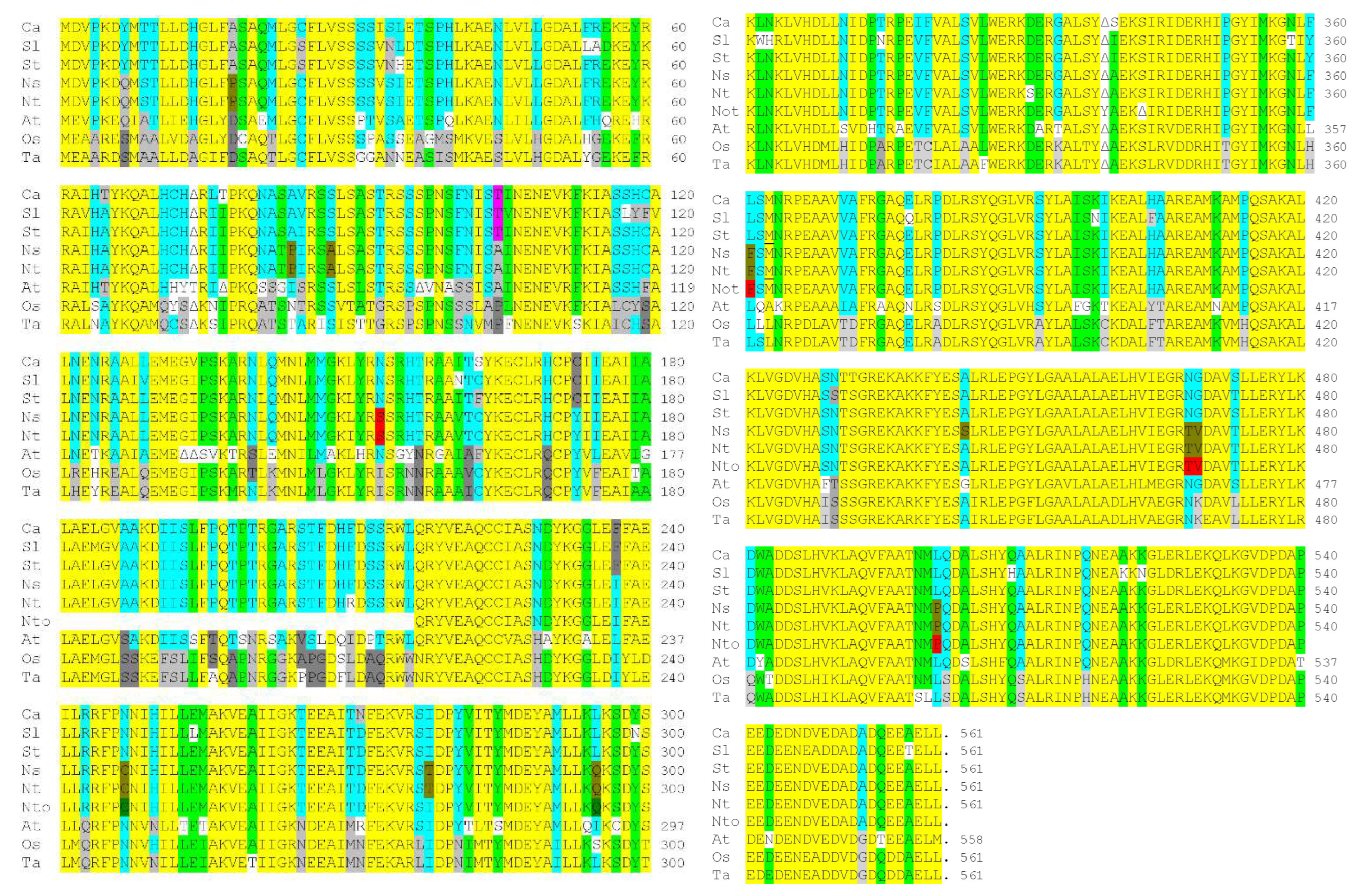
3.2. Sequence Analysis of IVR-like Proteins from Solanaceous Plants
3.3. Sequence Analysis of IVR-like Proteins from Various Plant Families
3.4. Analysis of the TPR Units in Tobacco IVRs vs. AtAPC7
3.5. Structural Analysis of the NtIVR
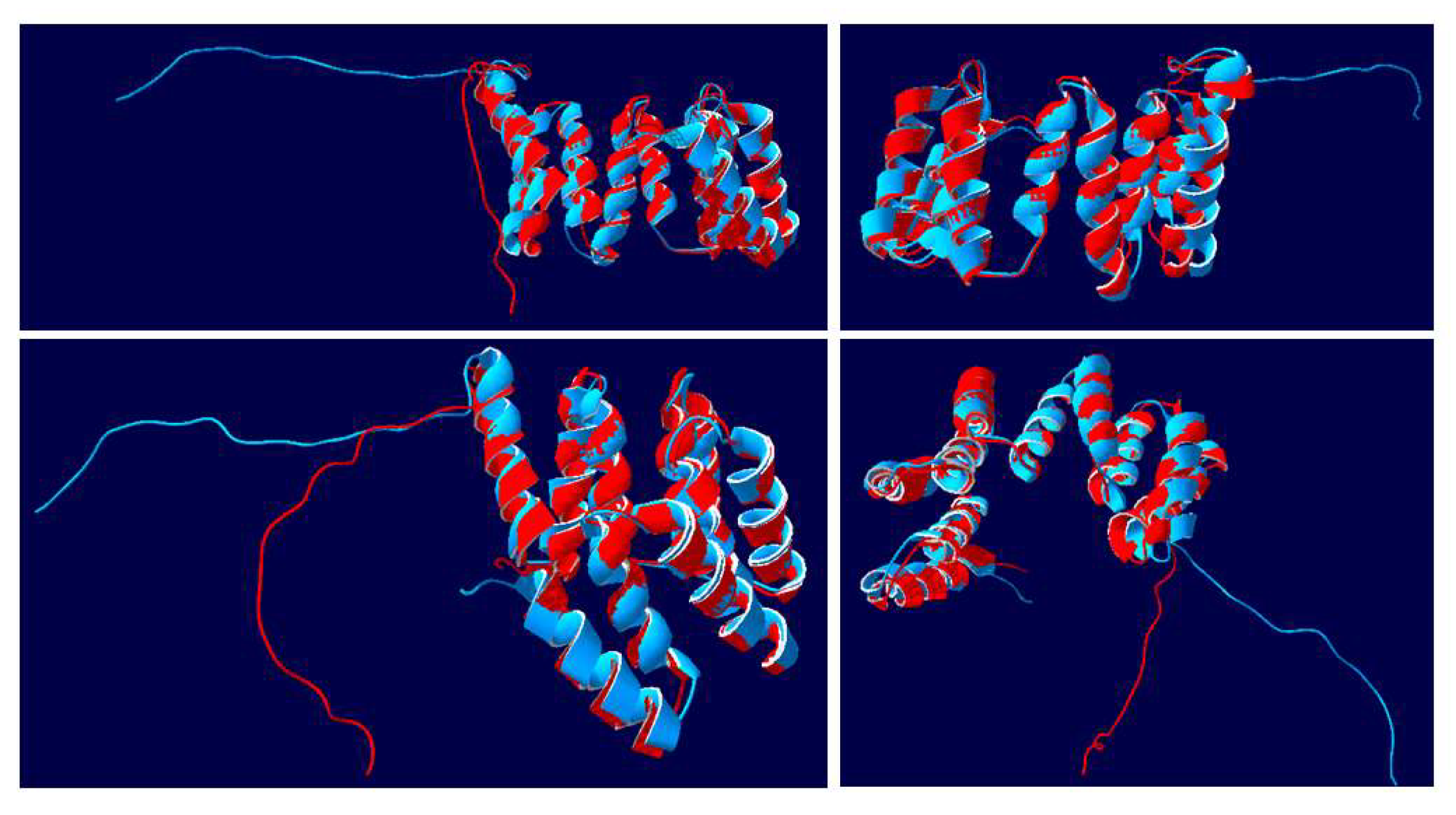
3.6. Structural Analysis of the NtIVR vs. AtAPC7
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, A. Dissecting the molecular network of virus-plant interactions: The complex roles of host factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H. Susceptibility genes to plant viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Zhang, Y. MAP kinase cascades in plant development and immune signaling. EMBO Rep. 2022, 23, e53817. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Sanfaçon, H. Plant translation factors and virus resistance. Viruses 2015, 7, 3392–3410. [Google Scholar] [CrossRef] [Green Version]
- Carr, J.P.; Lewsey, M.G.; Palukaitis, P. Signaling in induced resistance. Adv. Virus Res. 2010, 76, 57–121. [Google Scholar]
- Zvereva, A.S.; Pooggin, M.M. Silencing and innate immunity in plant defense against viral and non-viral pathogens. Viruses 2012, 4, 2578–2597. [Google Scholar] [CrossRef] [Green Version]
- Cillo, F.; Palukaitis, P. Transgenic resistance. Adv. Virus Res. 2014, 90, 35–146. [Google Scholar]
- Sela, I.; Appelbaum, S.W. Occurrence of antiviral factors in virus-infected plants. Virology 1962, 17, 543–548. [Google Scholar] [CrossRef]
- Sela, I. Plant virus interactions related to resistance and localization of viral infections. Adv. Virus Res. 1981, 26, 301–337. [Google Scholar]
- Edelbaum, O.; Ilan, N.; Grafi, G.; Sher, N.; Stram, Y.; Novick, D.; Tan, N.; Sela, I.; Rubenstein, M. Two antiviral proteins from tobacco: Purification and characterization by monoclonal antibodies to human β-interferon. Proc. Natl. Acad. Sci. USA 1990, 87, 588–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelbaum, O.; Sher, N.; Rubenstein, M.; Novick, D.; Tan, N.; Moyer, M.; Ward, E.; Ryals, J.; Sela, I. Two antiviral proteins, gp35 and gp22, correspond to β-1,3-glucanase and an isoform of PR-5. Plant Mol. Biol. 1991, 17, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Linthorst, H.J.M.; Meuwissem, R.L.J.; Kauffmann, S.; Bol, J.F. Constitutive expression of pathogenesis-related proteins PR-1, GRP, and PR-S in tobacco has no effect on virus infection. Plant Cell 1989, 1, 285–291. [Google Scholar] [PubMed]
- Reichman, M.; Devash, Y.; Suhadolnik, R.J.; Sela, I. Human leukocyte interferon and antiviral factor (AVF) from virus-infected plants stimulate plant-tissues to produce nucleotides with antiviral activity. Virology 1983, 128, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Loebenstein, G.; Gera, A. Inhibitor of virus replication released from tobacco mosaic virus-infected protoplasts of a local lesion-responding tobacco cultivar. Virology 1981, 114, 132–139. [Google Scholar] [CrossRef]
- Gera, A.; Loebenstein, G. Further studies of an inhibitor of virus replication from tobacco mosaic virus-infected protoplasts of a local lesion-responding tobacco cultivar. Phytopathology 1983, 73, 111–115. [Google Scholar] [CrossRef]
- Gera, A.; Loebenstein, G.; Shabtai, S. Enhanced tobacco mosaic virus production and supressed synthesis of a virus inhibitor in protoplasts exposed to antibiotics. Virology 1983, 127, 475–478. [Google Scholar] [CrossRef]
- Spiegel, S.; Gera, A.; Salomon, R.; Ahl, P.; Harlap, S.; Loebenstein, G. Recovery of an inhibitor of virus replication from the intercellular fluid of hypersensitive tobacco infected with tobacco mosaic virus and from uninfected induced-resistant tissue. Phytopathology 1989, 79, 258–262. [Google Scholar] [CrossRef]
- Gera, A.; Loebenstein, G. Evaluation of antisera to an inhibitor of virus replication (IVR). J. Phytopathol. 1989, 124, 366–371. [Google Scholar] [CrossRef]
- Gera, A.; Loebenstein, G.; Salamon, R.; Franck, A. Inhibitor of virus replication from protoplasts of a hypersensitive tobacco cultivar infected with tobacco mosaic virus is associated with a 23-K protein species. Phytopathology 1990, 80, 78–81. [Google Scholar] [CrossRef]
- Ahl, P.; Gianinazzi, S. b-proteins as a constitutive component in highly (TMV) resistant interspecific hybrids of Nicotiana glutinosa × Nicotiana debneyi. Plant Sci. Lett. 1982, 26, 173–181. [Google Scholar] [CrossRef]
- Gianinazzi, S.; Ahl, P. The genetic and molecular basis of b-proteins in genus Nicotiana. Neth. J. Plant Pathol. 1983, 89, 275–281. [Google Scholar] [CrossRef]
- Tahiri-Alaloui, A.; Dumas-Gaudot, E.; Gianinazzi, S.; Antoniw, J.F. Expression of the PR-b1″ gene in roots of two Nicotiana species and their amphidiploid hybrid infected with virulent and avirulent races of Chalara elegans. Plant Pathol. 1993, 42, 728–736. [Google Scholar] [CrossRef]
- Loebenstein, G.; Gera, A.; Gianinazzi, S. Constitutive production of an inhibitor of virus replication in the interspecific hybrid of Nicotiana glutinosa × Nicotiana debneyi. Physiol. Mol. Plant Pathol. 1990, 37, 145–151. [Google Scholar] [CrossRef]
- Gera, A.; Tam, Y.; Teverovsky, E.; Loebenstein, G. Enhanced tobacco mosaic virus production and suppressed synthesis of the inhibitor of virus replication in protoplasts and plants of local lesion responding cultivars exposed to 35 °C. Physiol. Mol. Plant Pathol. 1993, 43, 299–306. [Google Scholar] [CrossRef]
- Malamy, J.; Hennig, J.; Klessig, D.F. Temperature-dependent induction of salicylic acid and its conjugates during the resistance response to tobacco mosaic virus. Plant Cell 1992, 4, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Klessig, D.F. Isolation and characterization of a tobacco mosaic virus-inducible myb oncogene homolog from tobacco. Proc. Natl. Acad. Sci. USA 1996, 93, 14972–14977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akad, F.; Teverovsky, E.; David, A.; Czosnek, H.; Gidoni, D.; Gera, A.; Loebenstein, G. A cDNA from tobacco codes for an inhibitor of virus replication (IVR)-like protein. Plant Mol. Biol. 1999, 40, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Akad, F.; Teverovsky, E.; Gidoni, D.; Elad, Y.; Kirschner, B.; Ray-David, D.; Czosnek, H.; Loebenstein, G. Resistance to Tobacco mosaic virus and Botrytis cinerea in tobacco transformed with complementary DNA encoding an inhibitor of viral replication-like protein. Ann. Appl. Biol. 2005, 147, 89–100. [Google Scholar] [CrossRef]
- Fischer, U.; Dröge-Laser, W. Overexpression of NtERF5, a new member of the tobacco ethylene response transcription factor family enhances resistance to Tobacco mosaic virus. Mol. Plant-Microbe Interact. 2004, 17, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Rakhshandehroo, F.; Takeshita, M.; Squires, J.; Palukaitis, P. The influence of RNA-dependent RNA polymerase 1 on Potato virus Y infection and on other antiviral response genes. Mol. Plant-Microbe Interact. 2009, 22, 1312–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakhshandehroo, F.; Behboodi, B.S.; Mohammadi, M. Changes in peroxidase activity and transcript level of the MYB1 gene in transgenic tobacco plants silenced for the RDR-1 gene after systemic infection with Potato virus Y0. J. Phytopathol. 2012, 160, 187–194. [Google Scholar] [CrossRef]
- Yoon, J.-Y.; Baek, E.; Kim, M.; Palukaitis, P. The Virus-Induced Transcription Factor SHE1 Interacts with and Regulates the Expression of the Inhibitor of Virus Replication (IVR) in N Gene Tobacco; Department of Agricultural Convergence Technology, Jeonbuk National University: Jeonju, Republic of Korea, 2022; to be submitted. [Google Scholar]
- Yoon, J.-Y.; Palukaitis, P. Cucumber mosaic virus 1a protein interacts with the tobacco SHE1 transcription factor and partitions between the nucleus and the tonoplast membrane. Plant Pathol. J. 2021, 37, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Židek, A.; Potapenko, A. Highly accurate protein structures prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- ColabFold. Available online: https://conlab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb#scrollT0=iceGdbe_Pmt9 (accessed on 10 August 2021).
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Lim, H.; Ovchinnikov, S.; Steinegger, M. ColabFold:making protein folding accessible to all. Nature Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Mariani, V.; Biasini, M.; Barbato, A.; Schwede, T. LDDT: A local superposition-free score for comparing protein structures and models using distance difference tests. Bioinformatics 2013, 29, 2722–2728. [Google Scholar] [CrossRef] [Green Version]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Židek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef]
- Wilson, C.G.; Kajander, T.; Regan, L. The crystal structure of NlpI. A prokaryotic teratricopetide repeat protein with a globular fold. FEBS J. 2005, 272, 166–179. [Google Scholar] [CrossRef]
- Thornton, B.R.; Toczyski, D.P. Precise destruction: An emerging picture of the APC. Genes Dev. 2006, 20, 3069–3078. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Kim, K.; Kim, Y.; Kang, Y.; Lee, J.Y.; Kim, Y. Crystal structure of the N-terminal domain of anaphase-promoting complex subunit 7. J. Biol. Chem. 2009, 284, 15137–15146. [Google Scholar] [CrossRef] [Green Version]
- Ueda, H.; Yamaguchi, Y.; Sano, H. Direct interaction between the tobacco mosaic virus helicase domain and the ATP-bound resistance protein, N factor during the hypersensitive response in tobacco plants. Plant Mol. Biol. 2006, 61, 31–45. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, E.K.; Wang, Z.; French, R.; Kao, C.C. Interactions between structural domains of the RNA replication proteins of plant-infecting RNA viruses. J. Virol. 1998, 72, 7160–7169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Burch-Smith, T.; Schiff, M.; Feng, S.; Dinesh-Kumar, S.P. Molecular chaperone Hsp90 associates with resistance protein N and its signaling proteins SGT1 and Rar1 to modulate an immune response in plants. J. Biol. Chem. 2004, 279, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Involvement of MEK1 MAPKK, NTF6 MAPK, WRKY/MYB transcription factors, COI1 and CTR1 in N-mediated resistance to tobacco mosaic virus. Plant J. 2004, 38, 800–809. [Google Scholar] [CrossRef] [PubMed]
- van Verk, M.C.; Pappaioannou, D.; Neeleman, L.; Bol, J.F.; Linthorst, H.J.M. A novel WRKY transcription factor is required for induction of PR-1a gene expression by salicylic acid and bacterial elicitors. Plant Physiol. 2008, 146, 1983–1995. [Google Scholar] [CrossRef] [Green Version]
- Aoki, K.; Yano, K.; Suzuki, A.; Kawamura, S.; Sakurai, N.; Suda, K.; Kurabayashi, A.; Suzuki, T.; Tsugane, T.; Watanabe, M.; et al. Large-scale analysis of full-length cDNAs from the tomato (Solanum lycopersicum) cultivar Micro-Tom, a reference system for the Solanaceae genomics. BMC Genom. 2010, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Loebenstein, G. Local lesions and induced resistance. Adv. Virus Res. 2009, 75, 73–117. [Google Scholar]
- Visintin, R.; Prinz, S.; Amon, A. CDC20 and CDH1: A family of substrate-specific activators of APC-dependent proteolysis. Science 1997, 278, 460–463. [Google Scholar] [CrossRef]
- Vodermaier, H.C.; Gieffers, C.; Maurer-Stroh, S.; Eisenhaber, F.; Peters, J.-M. TRP subunits of the anaphase-promoting complex mediate binding to the activator protein CDH1. Curr. Biol. 2003, 13, 1459–1468. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.A.J.; Bi, Y.-M.; Darby, R.M.; Firek, S.; Draper, J. Compromising early salicylic acid accumulation delays the hypersensitive response and increases viral dispersal during lesion establishment in TMV-infected tobacco. Plant J. 1997, 12, 1113–1126. [Google Scholar] [CrossRef]
- Rizzo, T.R.; Palukaitis, P. Nucleotide sequence and evolutionary relationships of cucumber mosaic virus (CMV) strains: CMV RNA 1. J. Gen. Virol. 1989, 70, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, T.R.; Palukaitis, P. Nucleotide sequence and evolutionary relationships of cucumber mosaic virus (CMV) strains: CMV RNA 2. J. Gen. Virol. 1988, 69, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Whitham, S.; Dinesh-Kumar, S.P.; Choi, D.; Hehl, R.; Corr, C.; Baker, B. The product of the tobacco mosaic virus resistance gene N: Similarity to Toll and the Interleukin-1 receptor. Cell 1994, 78, 1101–1115. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1, and NPR/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Goelet, P.; Lomonossoff, G.P.; Butler, P.J.; Akam, M.E.; Gait, M.J.; Karn, J. Nucleotide sequence of tobacco mosaic virus RNA. Proc. Natl. Acad. Sci. USA 1982, 79, 5818–5822. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AtAPC7 | NsAPC7 | ||
|---|---|---|---|
| TPR No. | Borders (aa No.) | TPR No. | Borders (aa No.) |
| 1 | 43–76 | ||
| 2 | 138–171 | ||
| 3 | 212–245 | 1 | 219–243 |
| 4 | 246–279 | 2 | 248–278 |
| 5 | 314–346 | ||
| 6 | 348–380 | 3 | 339–379 |
| 7 | 381–413 | 4 | 383–411 |
| 8 | 414–448 | 5 | 417–446 |
| 9 | 450–482 | ||
| 10 | 483–515 | 6 | 485–514 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palukaitis, P.; Akbarimotlagh, M.; Baek, E.; Yoon, J.-Y. The Secret Life of the Inhibitor of Virus Replication. Viruses 2022, 14, 2782. https://doi.org/10.3390/v14122782
Palukaitis P, Akbarimotlagh M, Baek E, Yoon J-Y. The Secret Life of the Inhibitor of Virus Replication. Viruses. 2022; 14(12):2782. https://doi.org/10.3390/v14122782
Chicago/Turabian StylePalukaitis, Peter, Masoud Akbarimotlagh, Eseul Baek, and Ju-Yeon Yoon. 2022. "The Secret Life of the Inhibitor of Virus Replication" Viruses 14, no. 12: 2782. https://doi.org/10.3390/v14122782