
Exosomes Released by Influenza-Virus-Infected Cells Carry Factors Capable of Suppressing Immune Defense Genes in Naïve Cells

 , ,
, ,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Viruses, and Antibodies
2.2. Exosome Production
2.3. Next Generation Sequencing
2.4. Isolation of Exosomes and Influenza A Viral Particles
2.5. Dot Blotting
2.6. Transmission Electron Microscopy
2.7. Nano-Tracking Analysis
2.8. Determination of Infectious Activity
2.9. Stimulation of A549 Cells by Exosomes
2.10. Real-Time Quantitative PCR
2.11. Infection of Cells with Influenza A Virus in the Presence of Exosomes
3. Results and Discussion
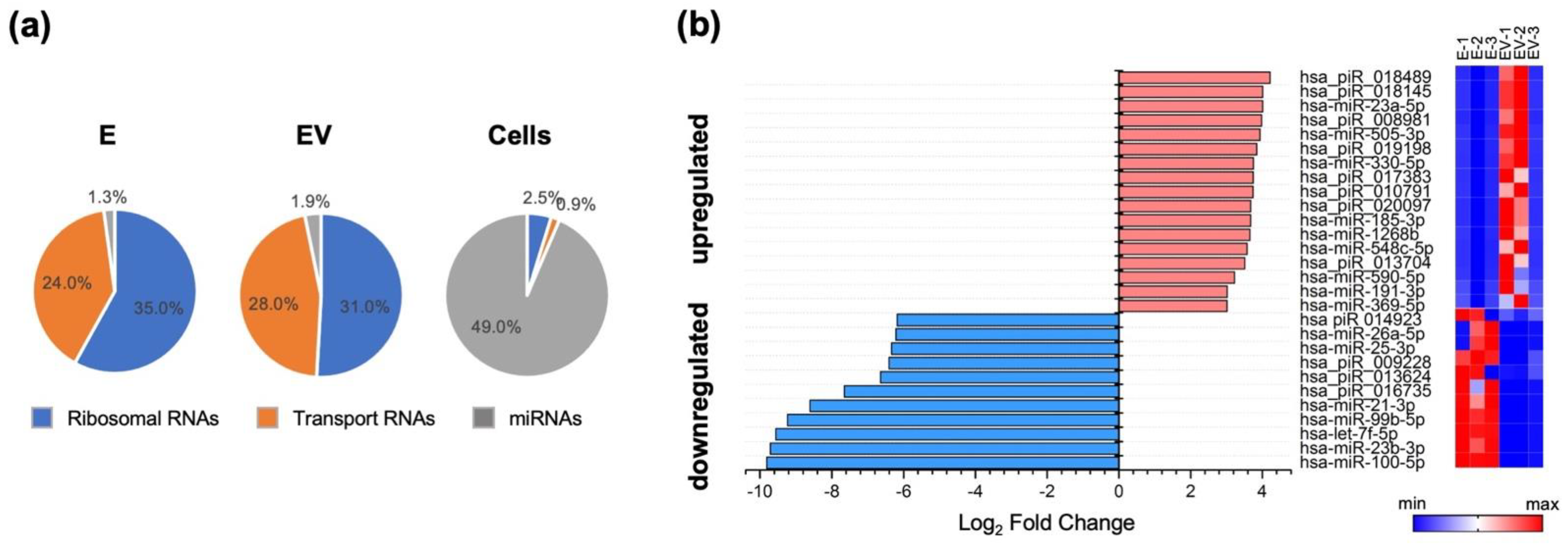
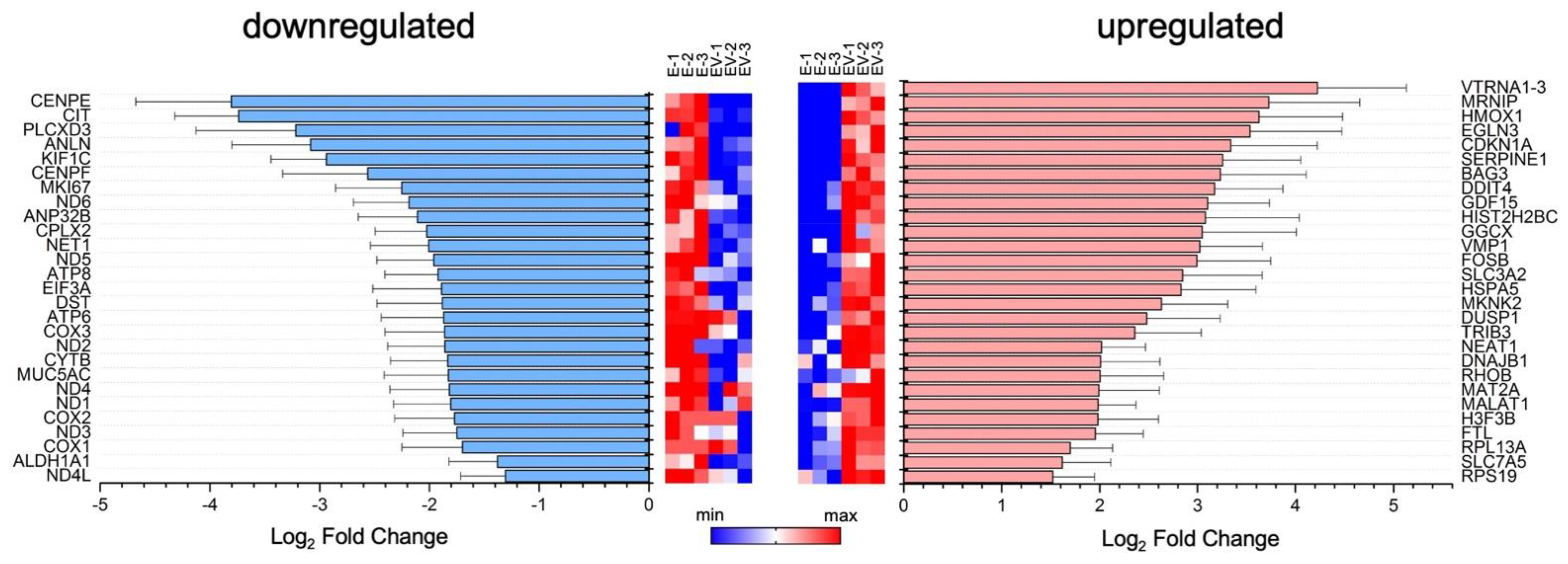
3.1. Exosomal miRNA and mRNA Profile Changes with Influenza Virus Infection
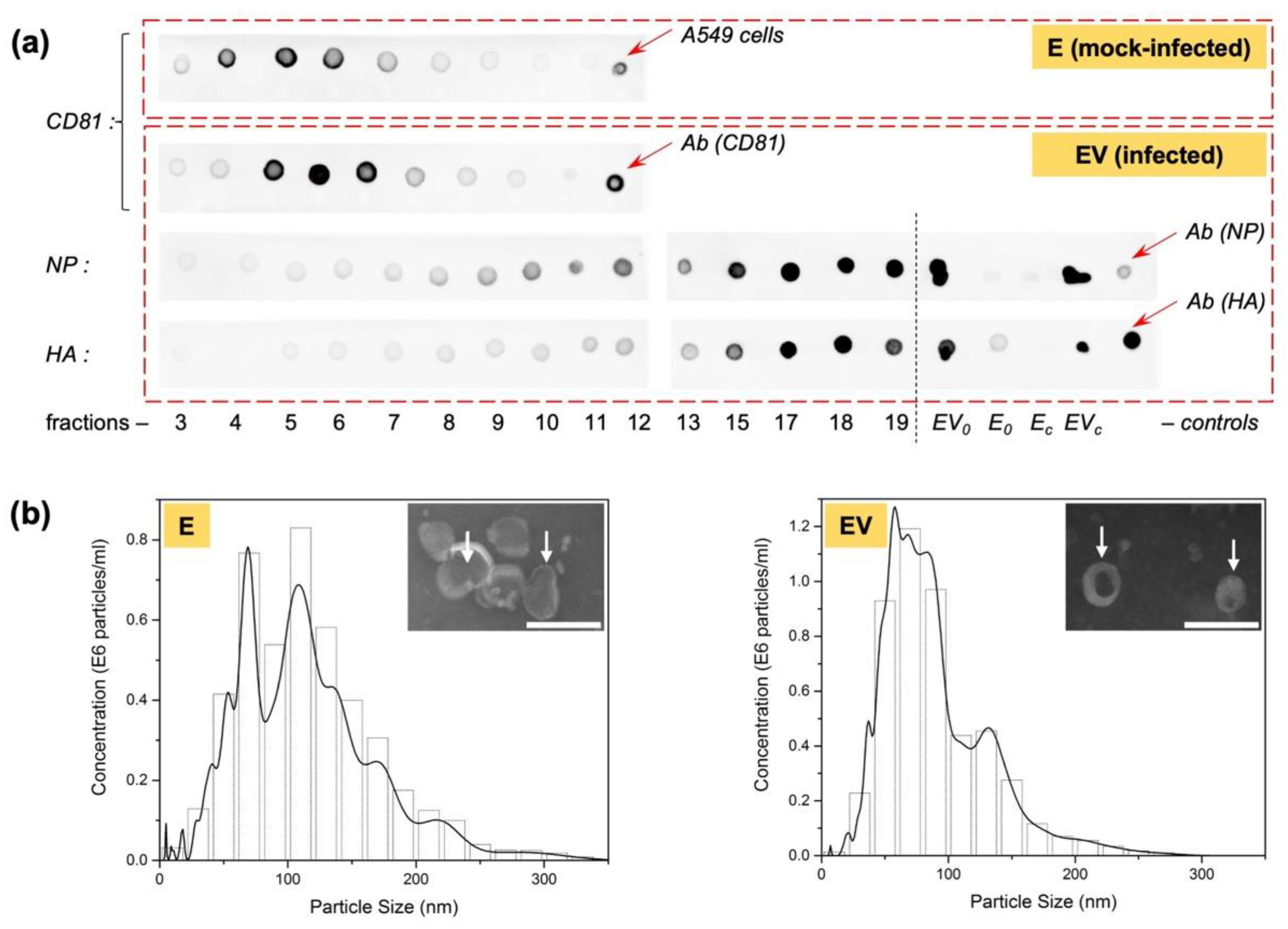
3.2. Isolation of Exosomes Secreted by IAV-Infected A549 Cells, Free of Viral Particles
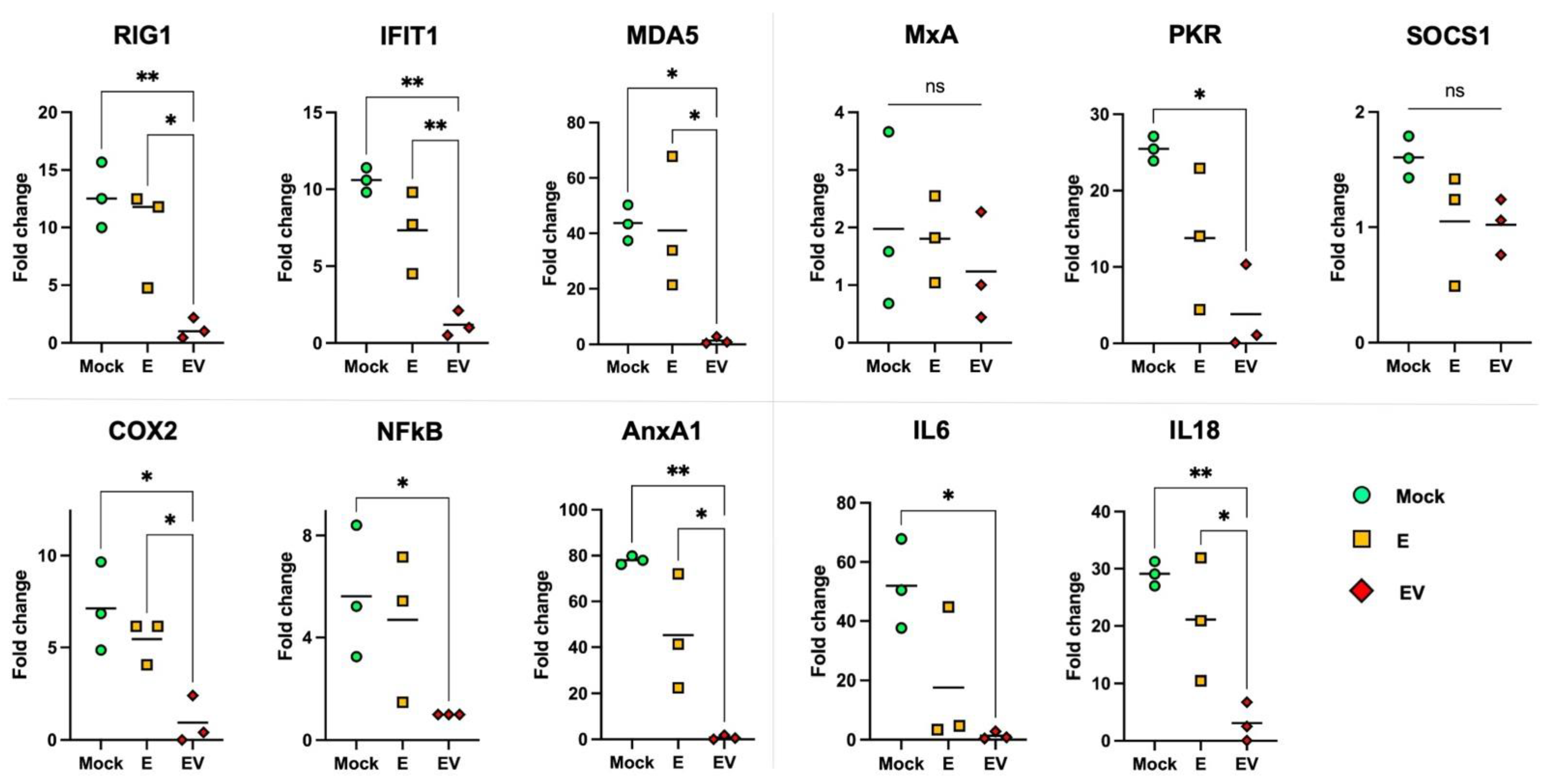
3.3. The Effect of E and EV on Intact A549 Cells
3.4. Identification of Viral RNA in Exosomes
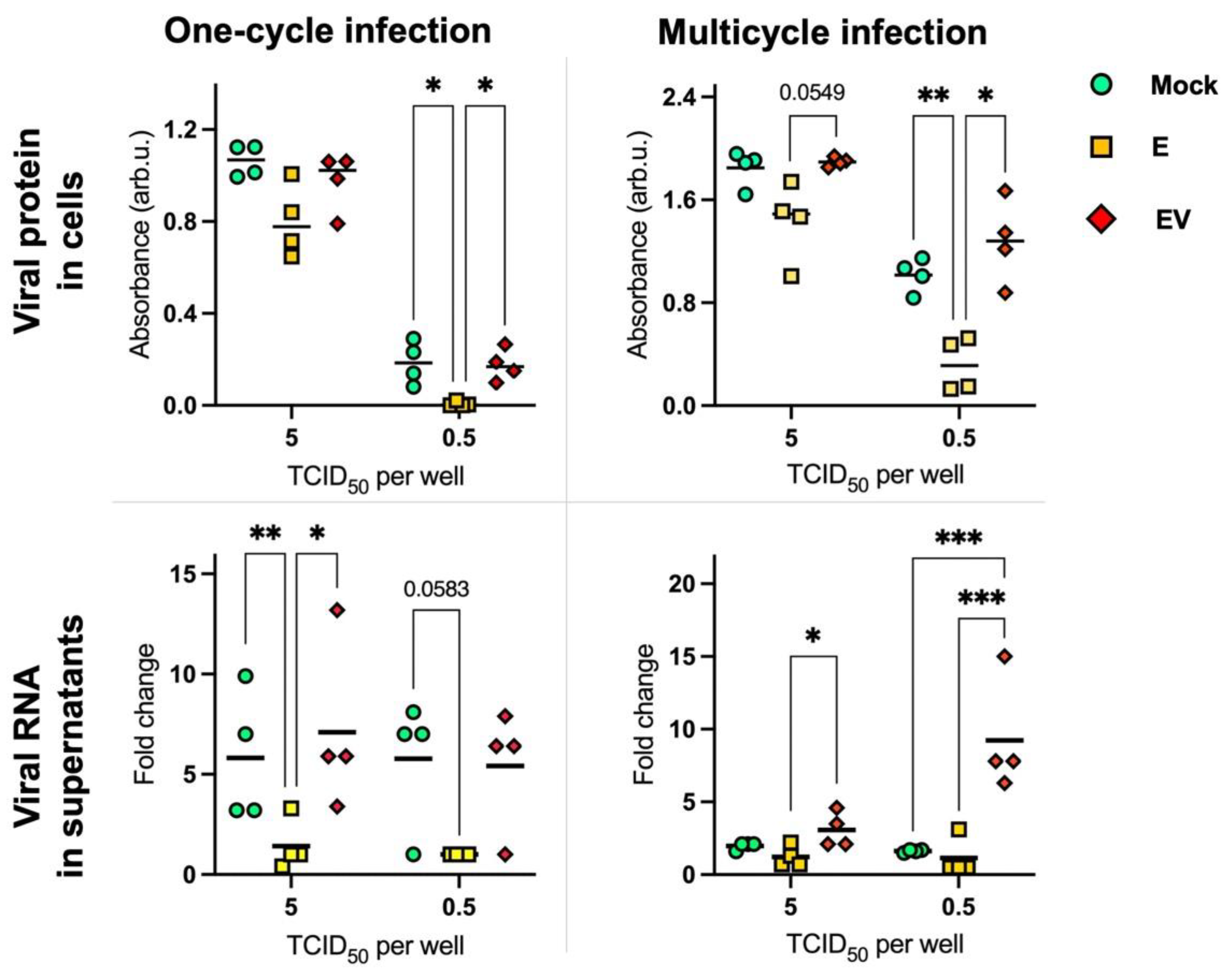
3.5. Effect of Exosomes on Influenza A Virus Replication
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Conde-Vancells, J.; Rodriguez-Suarez, E.; Gonzalez, E.; Berisa, A.; Gil, D.; Embade, N.; Valle, M.; Luka, Z.; Elortza, F.; Wagner, C.; et al. Candidate biomarkers in exosome-like vesicles purified from rat and mouse urine samples. Proteom. Clin. Appl. 2010, 4, 416–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, A.; Bajracharya, S.; Yuen, P.; Zhou, H.; Star, R.; Illei, G.; Alevizos, I. Exosomes from human saliva as a source of microRNA biomarkers. Oral Dis. 2010, 16, 34–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caby, M.-P.; Lankar, D.; Vincendeau-Scherrer, C.; Raposo, G.; Bonnerot, C. Exosomal-like vesicles are present in human blood plasma. Int. Immunol. 2005, 17, 879–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filén, J.-J.; Lahesmaa, R.; Norman, M.; Neve, E.P.A.; Scheynius, A.; Gabrielsson, S. Exosomes with Immune Modulatory Features Are Present in Human Breast Milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.; Rupp, C.; Stoeck, A.; Runz, S.; Fogel, M.; Lugert, S.; Hager, H.-D.; Abdel-Bakky, M.; Gutwein, P.; Altevogt, P. CD24 is a marker of exosomes secreted into urine and amniotic fluid. Kidney Int. 2007, 72, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Prado, N.; Marazuela, E.G.; Segura, E.; Fernández-García, H.; Villalba, M.; Théry, C.; Rodríguez, R.; Batanero, E. Exosomes from Bronchoalveolar Fluid of Tolerized Mice Prevent Allergic Reaction. J. Immunol. 2008, 181, 1519–1525. [Google Scholar] [CrossRef] [Green Version]
- Chahar, H.S.; Corsello, T.; Kudlicki, A.S.; Komaravelli, N.; Casola, A. Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells. Sci. Rep. 2018, 8, 387. [Google Scholar] [CrossRef] [Green Version]
- Familtseva, A.; Jeremic, N.; Tyagi, S.C. Exosomes: Cell-created drug delivery systems. Mol. Cell. Biochem. 2019, 459, 1–6. [Google Scholar] [CrossRef]
- Kharkwal, H.; Smith, C.G.; Wilson, D.W. Herpes Simplex Virus Capsid Localization to ESCRT-VPS4 Complexes in the Presence and Absence of the Large Tegument Protein UL36p. J. Virol. 2016, 90, 7257–7267. [Google Scholar] [CrossRef] [Green Version]
- Votteler, J.; Sundquist, W.I. Virus Budding and the ESCRT Pathway. Cell Host Microbe 2013, 14, 232–241. [Google Scholar] [CrossRef]
- Flanagan, J.; Middeldorp, J.; Sculley, T. Localization of the Epstein–Barr virus protein LMP 1 to exosomes. J. Gen. Virol. 2003, 84, 1871–1879. [Google Scholar] [CrossRef]
- Jiang, W.; Ma, P.; Deng, L.; Liu, Z.; Wang, X.; Liu, X.; Long, G. Hepatitis A virus structural protein pX interacts with ALIX and promotes the secretion of virions and foreign proteins through exosome-like vesicles. J. Extracell. Vesicles 2020, 9, 1716513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Randall, G.; Higginbottom, A.; Monk, P.; Rice, C.M.; McKeating, J.A. CD81 Is Required for Hepatitis C Virus Glycoprotein-Mediated Viral Infection. J. Virol. 2004, 78, 1448–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-M.; Tseng, C.-H.; Chen, Y.-C.; Yu, W.-Y.; Ho, M.-Y.; Ho, C.-Y.; Lai, M.M.C.; Su, W.-C. Exosome-delivered and Y RNA-derived small RNA suppresses influenza virus replication. J. Biomed. Sci. 2019, 26, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maemura, T.; Fukuyama, S.; Sugita, Y.; Lopes, T.J.S.; Nakao, T.; Noda, T.; Kawaoka, Y. Lung-Derived Exosomal miR-483-3p Regulates the Innate Immune Response to Influenza Virus Infection. J. Infect. Dis. 2018, 217, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Maemura, T.; Fukuyama, S.; Kawaoka, Y. High Levels of miR-483-3p Are Present in Serum Exosomes Upon Infection of Mice With Highly Pathogenic Avian Influenza Virus. Front. Microbiol. 2020, 11, 144. [Google Scholar] [CrossRef] [PubMed]
- Tambyah, P.A.; Sepramaniam, S.; Mohamed Ali, J.; Chai, S.C.; Swaminathan, P.; Armugam, A.; Jeyaseelan, K. microRNAs in Circulation Are Altered in Response to Influenza A Virus Infection in Humans. PLoS ONE 2013, 8, e76811. [Google Scholar] [CrossRef] [Green Version]
- Scheller, N.; Herold, S.; Kellner, R.; Bertrams, W.; Jung, A.L.; Janga, H.; Greulich, T.; Schulte, L.N.; Vogelmeier, C.F.; Lohmeyer, J.; et al. Proviral MicroRNAs Detected in Extracellular Vesicles From Bronchoalveolar Lavage Fluid of Patients With Influenza Virus–Induced Acute Respiratory Distress Syndrome. J. Infect. Dis. 2018, 219, 540–543. [Google Scholar] [CrossRef]
- Cypryk, W.; Lorey, M.; Puustinen, A.; Nyman, T.A.; Matikainen, S. Proteomic and Bioinformatic Characterization of Extracellular Vesicles Released from Human Macrophages upon Influenza A Virus Infection. J. Proteome Res. 2016, 16, 217–227. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartley, S.W.; Mullikin, J.C. QoRTs: A comprehensive toolset for quality control and data processing of RNA-Seq experiments. BMC Bioinform. 2015, 16, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankish, A.; Diekhans, M.; Jungreis, I.; Lagarde, J.; Loveland, J.E.; Mudge, J.M.; Sisu, C.; Wright, J.C.; Armstrong, J.; Barnes, I.; et al. GENCODE 2021. Nucleic Acids Res. 2020, 49, D916–D923. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. Feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Zenkova, D.; Kamenev, V.; Sablina, R.; Artyomov, M.; Sergushichev, A. Phantasus: Visual and Interactive Gene Expression Analysis. 2018. Available online: https://bioconductor.org/packages/release/bioc/html/phantasus.html (accessed on 8 October 2022).
- Croft, D.; O’Kelly, G.; Wu, G.; Haw, R.; Gillespie, M.; Matthews, L.; Caudy, M.; Garapati, P.; Gopinath, G.; Jassal, B.; et al. Reactome: A database of reactions, pathways and biological processes. Nucleic Acids Res. 2011, 39, D691–D697. [Google Scholar] [CrossRef] [Green Version]
- Carbon, S.; Mungall, C. Gene Ontology Data Archive. Zenodo. 2020. Available online: https://zenodo.org (accessed on 1 December 2020).
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Griss, J.; Viteri, G.; Sidiropoulos, K.; Nguyen, V.; Fabregat, A.; Hermjakob, H. ReactomeGSA—Efficient Multi-Omics Comparative Pathway Analysis. Mol. Cell. Proteom. 2020, 19, 2115–2125. [Google Scholar] [CrossRef]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2021, 060012. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. In Current Protocols in Cell Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Lu, J.; Wang, R.; Yang, Z.; Zhou, X.; Huang, P. miR-21-3p Regulates Influenza A Virus Replication by Targeting Histone Deacetylase-8. Front. Cell. Infect. Microbiol. 2018, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Li, J.; Song, L.; Wu, J.; Huang, W. Influenza A virus-induced downregulation of miR-26a contributes to reduced IFNα/β production. Virol. Sin. 2017, 32, 261–270. [Google Scholar] [CrossRef]
- Gu, X.; Gao, Y.; Mu, D.-G.; Fu, E.-Q. MiR-23a-5p modulates mycobacterial survival and autophagy during mycobacterium tuberculosis infection through TLR2/MyD88/NF-κB pathway by targeting TLR2. Exp. Cell Res. 2017, 354, 71–77. [Google Scholar] [CrossRef]
- Li, Y.; Xie, J.; Xu, X.; Wang, J.; Ao, F.; Wan, Y.; Zhu, Y. MicroRNA-548 down-regulates host antiviral response via direct targeting of IFN-λ1. Protein Cell 2012, 4, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Zhou, J.; Wang, H. Host microRNAs and exosomes that modulate influenza virus infection. Virus Res. 2020, 279, 197885. [Google Scholar] [CrossRef]
- Cho, H.-Y.; Park, S.; Miller, L.; Lee, H.-C.; Langenbach, R.; Kleeberger, S.R. Role for Mucin-5AC in Upper and Lower Airway Pathogenesis in Mice. Toxicol. Pathol. 2021, 49, 1077–1099. [Google Scholar] [CrossRef]
- Nguyen, N.N.T.; Lim, Y.-S.; Nguyen, L.P.; Tran, S.C.; Luong, T.T.D.; Nguyen, T.T.T.; Pham, H.T.; Mai, H.N.; Choi, J.-W.; Han, S.-S.; et al. Hepatitis C Virus Modulates Solute carrier family 3 member 2 for Viral Propagation. Sci. Rep. 2018, 8, 15486. [Google Scholar] [CrossRef]
- Arif, A.; Chatterjee, P.; Moodt, R.A.; Fox, P.L. Heterotrimeric GAIT Complex Drives Transcript-Selective Translation Inhibition in Murine Macrophages. Mol. Cell. Biol. 2012, 32, 5046–5055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buxadé, M.; Parra, J.L.; Rousseau, S.; Shpiro, N.; Marquez, R.; Morrice, N.; Bain, J.; Espel, E.; Proud, C.G. The Mnks Are Novel Components in the Control of TNFα Biosynthesis and Phosphorylate and Regulate hnRNP A1. Immunity 2005, 23, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, S.; Sharma, B.; Kaur, S.; Majchrzak, B.; Ueda, T.; Fukunaga, R.; Verma, A.K.; Fish, E.N.; Platanias, L.C. Essential Role for Mnk Kinases in Type II Interferon (IFNγ) Signaling and Its Suppressive Effects on Normal Hematopoiesis. J. Biol. Chem. 2011, 286, 6017–6026. [Google Scholar] [CrossRef] [Green Version]
- Ord, T.; Ord, T. Mammalian Pseudokinase TRIB3 in Normal Physiology and Disease: Charting the Progress in Old and New Avenues. Curr. Protein Pept. Sci. 2017, 18, 819–842. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xu, L.G.; Zhai, Z.; Shu, H.B. SINK Is a p65-interacting Negative Regulator of NF-κB-dependent Transcription. J. Biol. Chem. 2003, 278, 27072–27079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzymski, T.; Paantjens, A.; Bod, J.; Harris, A.L. Multiple pathways are involved in the anoxia response of SKIP3 including HuR-regulated RNA stability, NF-κB and ATF4. Oncogene 2008, 27, 4532–4543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Moran, A.P.; Duggan, S.P.; Ahmed, S.E.; Mohamed, A.S.; Windle, H.J.; O’Neill, L.A.; Kelleher, D.P. Tribbles 3: A Novel Regulator of TLR2-Mediated Signaling in Response to Helicobacter Pylori Lipopolysaccharide. J. Immunol. 2011, 186, 2462–2471. [Google Scholar] [CrossRef] [Green Version]
- Wurzer, W.J.; Ehrhardt, C.; Pleschka, S.; Berberich-Siebelt, F.; Wolff, T.; Walczak, H.; Planz, O.; Ludwig, S. NF-κB-dependent Induction of Tumor Necrosis Factor-related Apoptosis-inducing Ligand (TRAIL) and Fas/FasL Is Crucial for Efficient Influenza Virus Propagation. J. Biol. Chem. 2004, 279, 30931–30937. [Google Scholar] [CrossRef] [Green Version]
- Nimmerjahn, F.; Dudziak, D.; Dirmeier, U.; Hobom, G.; Riedel, A.; Schlee, M.; Staudt, L.M.; Rosenwald, A.; Behrends, U.; Bornkamm, G.W.; et al. Active NF-κB signalling is a prerequisite for influenza virus infection. J. Gen. Virol. 2004, 85, 2347–2356. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Ekström, K.; Valadi, H.; Sjöstrand, M.; Malmhäll, C.; Bossios, A.; Eldh, M.; Lötvall, J. Characterization of mRNA and microRNA in human mast cell-derived exosomes and their transfer to other mast cells and blood CD34 progenitor cells. J. Extracell. Vesicles 2012, 1, 18389. [Google Scholar] [CrossRef] [PubMed]
- Skog, J.; Würdinger, T.; Van Rijn, S.; Meijer, D.H.; Gainche, L.; Curry, W.T., Jr.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Cantin, R.; Diou, J.; Bélanger, D.; Tremblay, A.M.; Gilbert, C. Discrimination between exosomes and HIV-1: Purification of both vesicles from cell-free supernatants. J. Immunol. Methods 2008, 338, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, E.C.; Charles, P.D.; Hester, S.S.; Thomas, B.; Trudgian, D.; Martínez-Alonso, M.; Fodor, E. Conserved and host-specific features of influenza virion architecture. Nat. Commun. 2014, 5, 4816. [Google Scholar] [CrossRef] [Green Version]
- Barry, R.D.; Davies, P. The Sedimentation of Influenza Virus and its RNA in Sucrose Density Gradients. J. Gen. Virol. 1968, 2, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.Y.; Cheung, C.Y.; Nicholls, J.M.; Hui, K.P.Y.; Leung, C.Y.H.; Uiprasertkul, M.; Tipoe, G.L.; Lau, Y.L.; Poon, L.L.M.; Ip, N.Y.; et al. Hyperinduction of Cyclooxygenase-2–Mediated Proinflammatory Cascade: A Mechanism for the Pathogenesis of Avian Influenza H5N1 Infection. J. Infect. Dis. 2008, 198, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Li, Y.; Pan, R.; Zou, X. Characterizing and controlling the inflammatory network during influenza A virus infection. Sci. Rep. 2014, 4, 3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owaki, T.; Asakawa, M.; Morishima, N.; Mizoguchi, I.; Fukai, F.; Takeda, K.; Mizuguchi, J.; Yoshimoto, T. STAT3 Is Indispensable to IL-27-Mediated Cell Proliferation but Not to IL-27-Induced Th1 Differentiation and Suppression of Proinflammatory Cytokine Production. J. Immunol. 2008, 180, 2903–2911. [Google Scholar] [CrossRef] [Green Version]
- Sander, W.J.; O’Neill, H.G.; Pohl, C.H. Prostaglandin E2 As a Modulator of Viral Infections. Front. Physiol. 2017, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, S.; Chen, Q.; Chen, Y.; Chi, X.; Zhang, L.; Huang, S.; Gao, G.F.; Chen, J.-L. Suppression of Interferon Lambda Signaling by SOCS-1 Results in Their Excessive Production during Influenza Virus Infection. PLoS Pathog. 2014, 10, e1003845, Correction in 2016, 12, e1005402. [Google Scholar] [CrossRef] [PubMed]
- Lozhkov, A.A.; Plotnikova, M.A.; Egorova, M.A.; Baranovskaya, I.L.; Elpaeva, E.A.; Klotchenko, S.A.; Vasin, A.V. Simultaneous Detection of RIG-1, MDA5, and IFIT-1 Expression Is a Convenient Tool for Evaluation of the Interferon-Mediated Response. Viruses 2022, 14, 2090. [Google Scholar] [CrossRef] [PubMed]
- Yap, G.L.R.; Sachaphibulkij, K.; Foo, S.L.; Cui, J.; Fairhurst, A.-M.; Lim, L.H.K. Annexin-A1 promotes RIG-I-dependent signaling and apoptosis via regulation of the IRF3–IFNAR–STAT1–IFIT1 pathway in A549 lung epithelial cells. Cell Death Dis. 2020, 11, 463. [Google Scholar] [CrossRef]
- Ge, Y.; Liu, K.; Chi, Y.; Zhu, X.; Wu, T.; Zhao, K.; Qiao, Q.; Wu, B.; Zhu, F.; Cui, L. Exosomal microRNA expression profiles derived from A549 human lung cells in response to influenza A/H1N1pdm09 infection. Virology 2022, 574, 9–17. [Google Scholar] [CrossRef]
- Chaimayo, C.; Dunagan, M.; Hayashi, T.; Santoso, N.; Takimoto, T. Specificity and functional interplay between influenza virus PA-X and NS1 shutoff activity. PLOS Pathog. 2018, 14, e1007465. [Google Scholar] [CrossRef]
- Hayashi, T.; Chaimayo, C.; McGuinness, J.; Takimoto, T. Critical Role of the PA-X C-Terminal Domain of Influenza A Virus in Its Subcellular Localization and Shutoff Activity. J. Virol. 2016, 90, 7131–7141. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Wang, X.; Liu, X. NS1: A Key Protein in the “Game” Between Influenza A Virus and Host in Innate Immunity. Front. Cell. Infect. Microbiol. 2021, 11, 670177. [Google Scholar] [CrossRef]
- Shaldzhyan, A.A.; Zabrodskaya, Y.A.; Baranovskaya, I.L.; Sergeeva, M.V.; Gorshkov, A.N.; Savin, I.I.; Shishlyannikov, S.M.; Ramsay, E.S.; Protasov, A.V.; Kukhareva, A.P.; et al. Old dog, new tricks: Influenza A virus NS1 and in vitro fibrillogenesis. Biochimie 2021, 190, 50–56. [Google Scholar] [CrossRef]
- Bedford, J.G.; Infusini, G.; Dagley, L.F.; Villalon-Letelier, F.; Zheng, M.Z.M.; Bennett-Wood, V.; Reading, P.C.; Wakim, L.M. Airway Exosomes Released During Influenza Virus Infection Serve as a Key Component of the Antiviral Innate Immune Response. Front. Immunol. 2020, 11, 887. [Google Scholar] [CrossRef]
- Kesimer, M.; Scull, M.; Brighton, B.; DeMaria, G.; Burns, K.; O’Neal, W.; Pickles, R.J.; Sheehan, J.K. Characterization of exosome-like vesicles released from human tracheobronchial ciliated epithelium: A possible role in innate defense. FASEB J. 2009, 23, 1858–1868. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Probe |
|---|---|---|---|
| IFIT1 | AAACTTCGGAGAAAGGCATTAGAT | TGAAATGAAATGTGAAAGTGGCTG | (HEX)-CCTGAGACTGGCTGCTGACTTTGAGAAC-(BHQ1) |
| MxA | GAGACAATCGTGAAACAGCAAATCA | TATCGAAACTCTGTGAAAGCAAGC | (HEX)-CACTGGAAGAGCCGGCTGGGATATG-(BHQ1) |
| RIG1 | GAGCACTTGTGGACGCTTTA | ATACACTTCTGTGCCGGGAG | (ROX)-CCTGGCATATTGACTGGACGTGGC-(BHQ2) |
| MDA5 | AAACCCATGACACAGAATGAACA | TGTGAGCAACCAGGACGTAG | (Cy5.5)-CACAGTGGCAGAAGAAGGTCTGGA-(BHQ3) |
| SOCS1 | CCTGGTTGTTGTAGCAGCTTA | CCTGGTTTGTGCAAAGATACTG | (ROX)-CCTGGTTGTTGTAGCAGCTTA-(BHQ2) |
| PKR | GAAAGCGAACAAGGAGTAAGGGA | CCATCCCGTAGGTCTGTGAAA | Cy5-AGCCCCAAAGCGTAGAGGTCCACTTCC-BHQ3 |
| NFκB | GCTCAGTGAGCCCATGGAAT | TGATGCTCTTGAAGGTCTCATATGTC | (FAM)-TCACCGGATTGAGGAGAAAC-(BHQ1) |
| COX2 | CTGATGATTGCCCGACTCCC | GGCGCAGTTTACGCTGTCTA | (ROX)-GGGCTGGGCCATGGGGTGGA-(BHQ2) |
| AnxA1 | ACCACCAGAAGCTATCCACAA | CGAGTTCCAACACCTTTCATGG | (Cy5.5)-AAGTGCGCCACAAGCAAACCAGC-(BHQ3) |
| GAPDH | CAGTCAGCCGCATCTTCTTTTGCGTCG | CAGAGTTAAAAGCAGCCCTGGTGACCAG | (FAM)-TGGGGAAGGTGAAGGTCGGAGTCAACGGATTTGGTC-(BHQ1) |
| IL6 | CCACTCACCTCTTCAGAACG | CATCTTTGGAAGGTTCAGGTTG | (HEX)-AAATTCGGTACATCCTCGACGGCATC-(BHQ1) |
| IL18 | AAACTATTTGTCGCAGGAATAAAGAT | GCTTGCCAAAGTAATCTGATTCC | (ROX)-TGCAATTGTCTTCTACTGGTTCAGCAGC-(BHQ2) |
| NS | TACCTAACTGACATGACTCTTGAG | TCGCCTGGTCCATTCTGATAC | - |
| M2 | GACCRATCCTGTCACCTCTGAC | AGGGCATTYTGGACAAAKCGTCTA | (FAM)-TGCAGTCCTCGCTCACTGGGCACG-(BHQ1) |
| PA | TTCAGGCACTTAGGGACAA | AGGAAGGAGTTGAACCAAGA | (HEX)-TGCCTGATTAATGATCCCTGGGTTTTGC-(BHQ1) |
| Sample | RT Primers | GAPDH | IAV Genes | ||
|---|---|---|---|---|---|
| M2 | NS | PA | |||
| EV | dT16 | 37.7 | 38.28 | 38.43 | N/A |
| Random9 | 38.55 | 32.33 | 30.83 | 36.29 | |
| E | dT16 | 38.76 | N/A | N/A | N/A |
| Random9 | 37.4 | N/A | N/A | N/A | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zabrodskaya, Y.; Plotnikova, M.; Gavrilova, N.; Lozhkov, A.; Klotchenko, S.; Kiselev, A.; Burdakov, V.; Ramsay, E.; Purvinsh, L.; Egorova, M.; et al. Exosomes Released by Influenza-Virus-Infected Cells Carry Factors Capable of Suppressing Immune Defense Genes in Naïve Cells. Viruses 2022, 14, 2690. https://doi.org/10.3390/v14122690
Zabrodskaya Y, Plotnikova M, Gavrilova N, Lozhkov A, Klotchenko S, Kiselev A, Burdakov V, Ramsay E, Purvinsh L, Egorova M, et al. Exosomes Released by Influenza-Virus-Infected Cells Carry Factors Capable of Suppressing Immune Defense Genes in Naïve Cells. Viruses. 2022; 14(12):2690. https://doi.org/10.3390/v14122690
Chicago/Turabian StyleZabrodskaya, Yana, Marina Plotnikova, Nina Gavrilova, Alexey Lozhkov, Sergey Klotchenko, Artem Kiselev, Vladimir Burdakov, Edward Ramsay, Lada Purvinsh, Marja Egorova, and et al. 2022. "Exosomes Released by Influenza-Virus-Infected Cells Carry Factors Capable of Suppressing Immune Defense Genes in Naïve Cells" Viruses 14, no. 12: 2690. https://doi.org/10.3390/v14122690