
Utility of Alternative Promoters for Foreign Gene Expression Using the Baculovirus Expression Vector System

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Recombinant Baculovirus Generation, Amplification, and Quantification
2.3. Infections
2.4. Flow Cytometry and Analysis
2.5. SEAP Activity Assays
2.6. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.7. Bioinformatics
3. Results
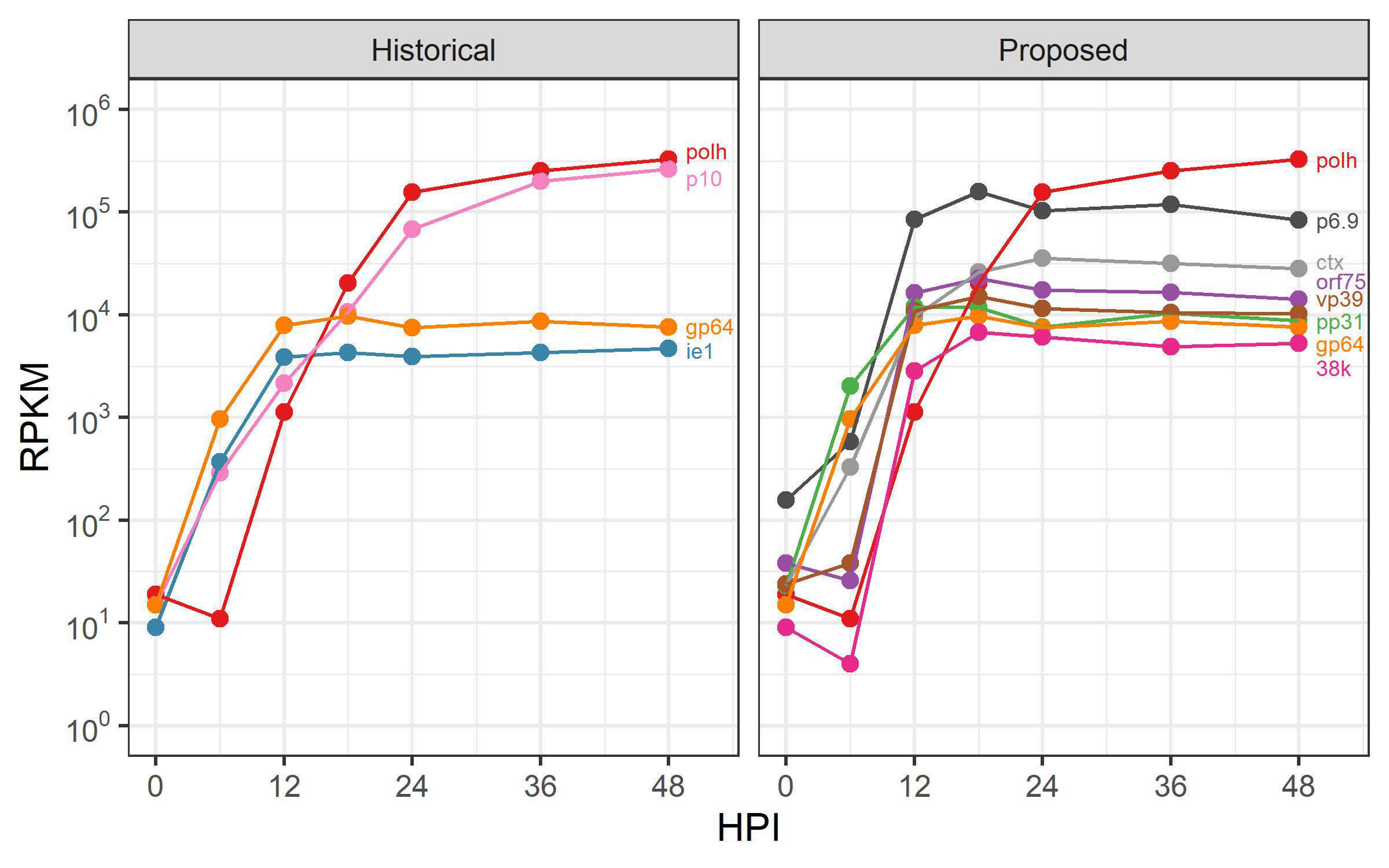
3.1. Selection of AcMNPV Promoters
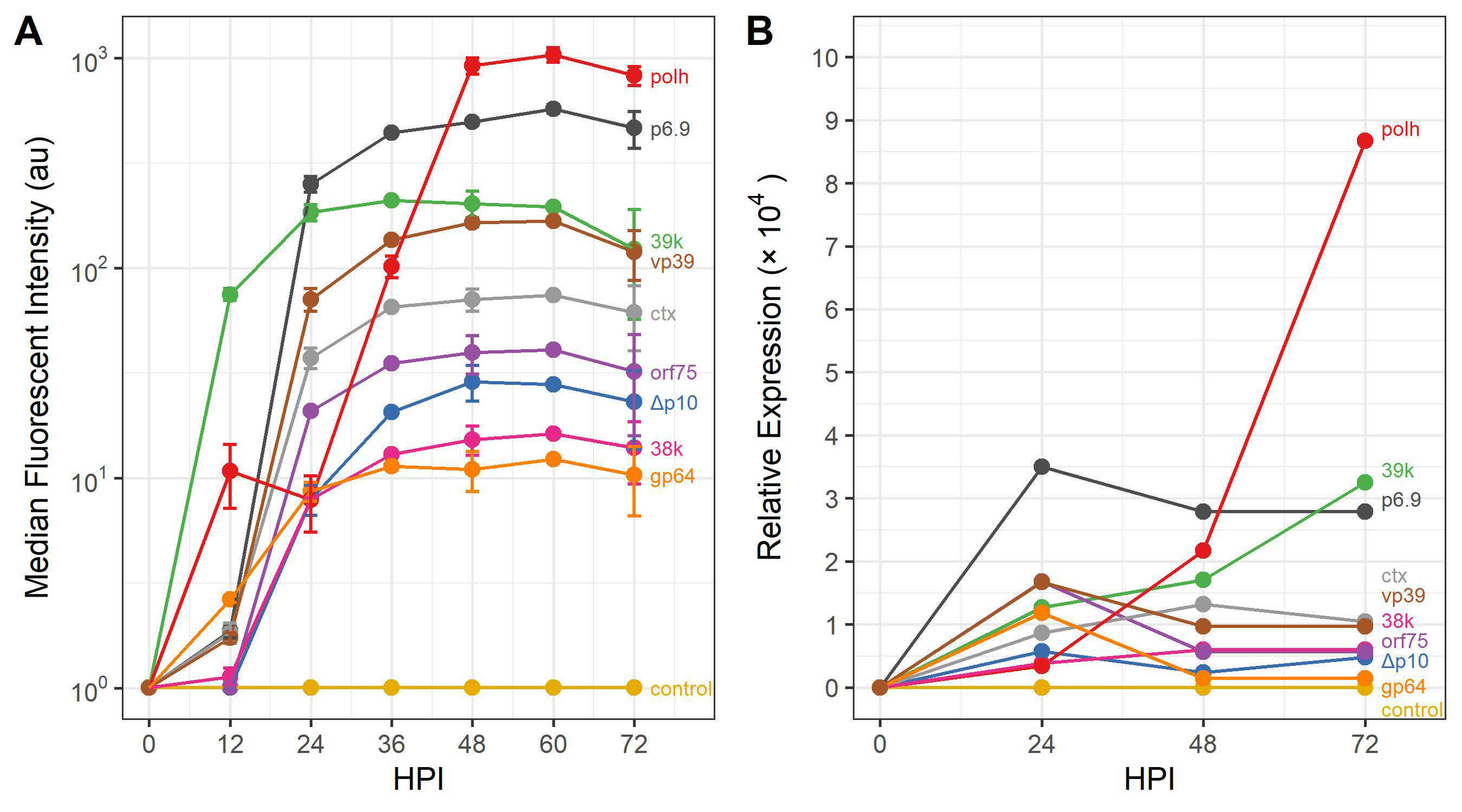
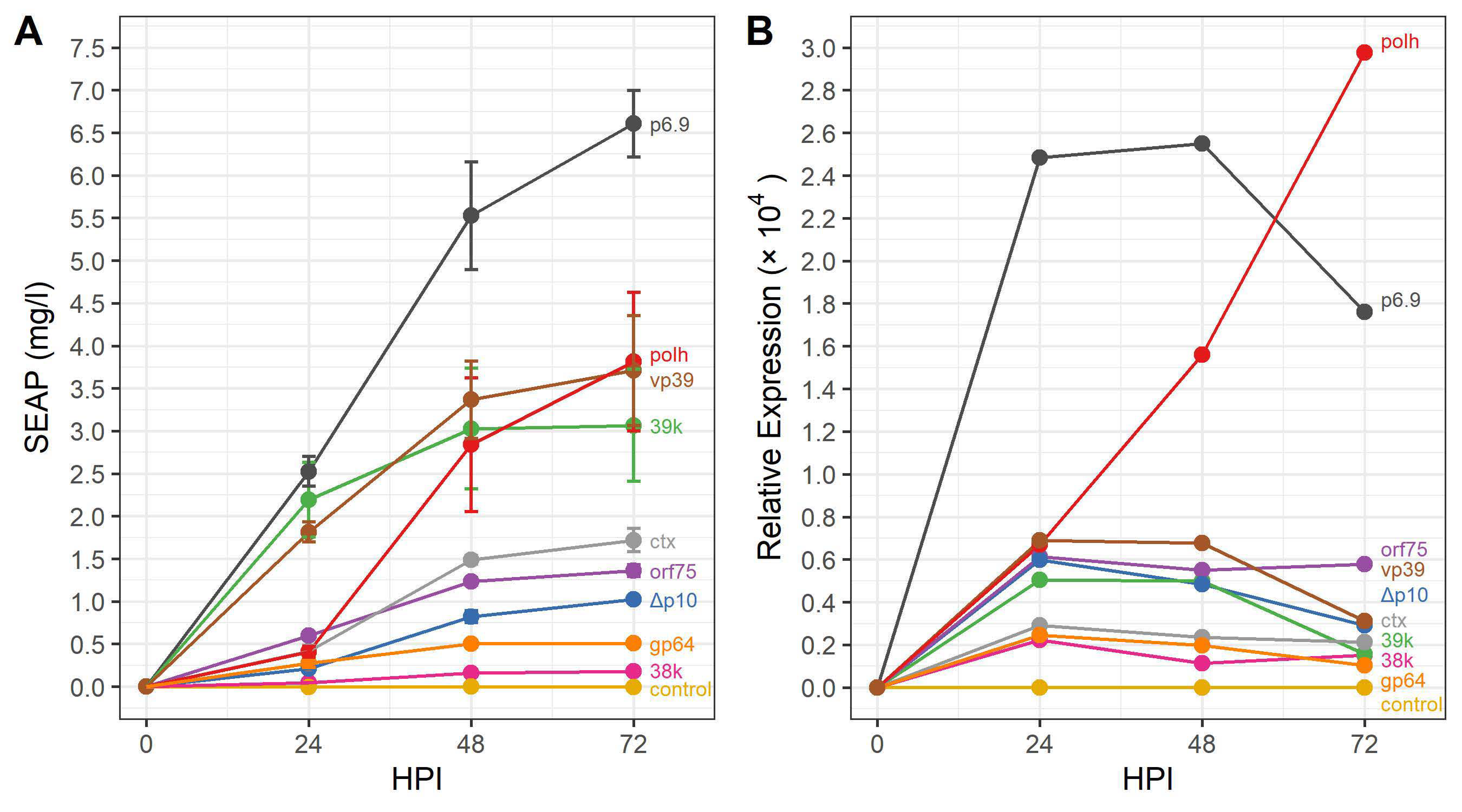
3.2. Evaluating the Expression Profile of AcMNPV Promoters
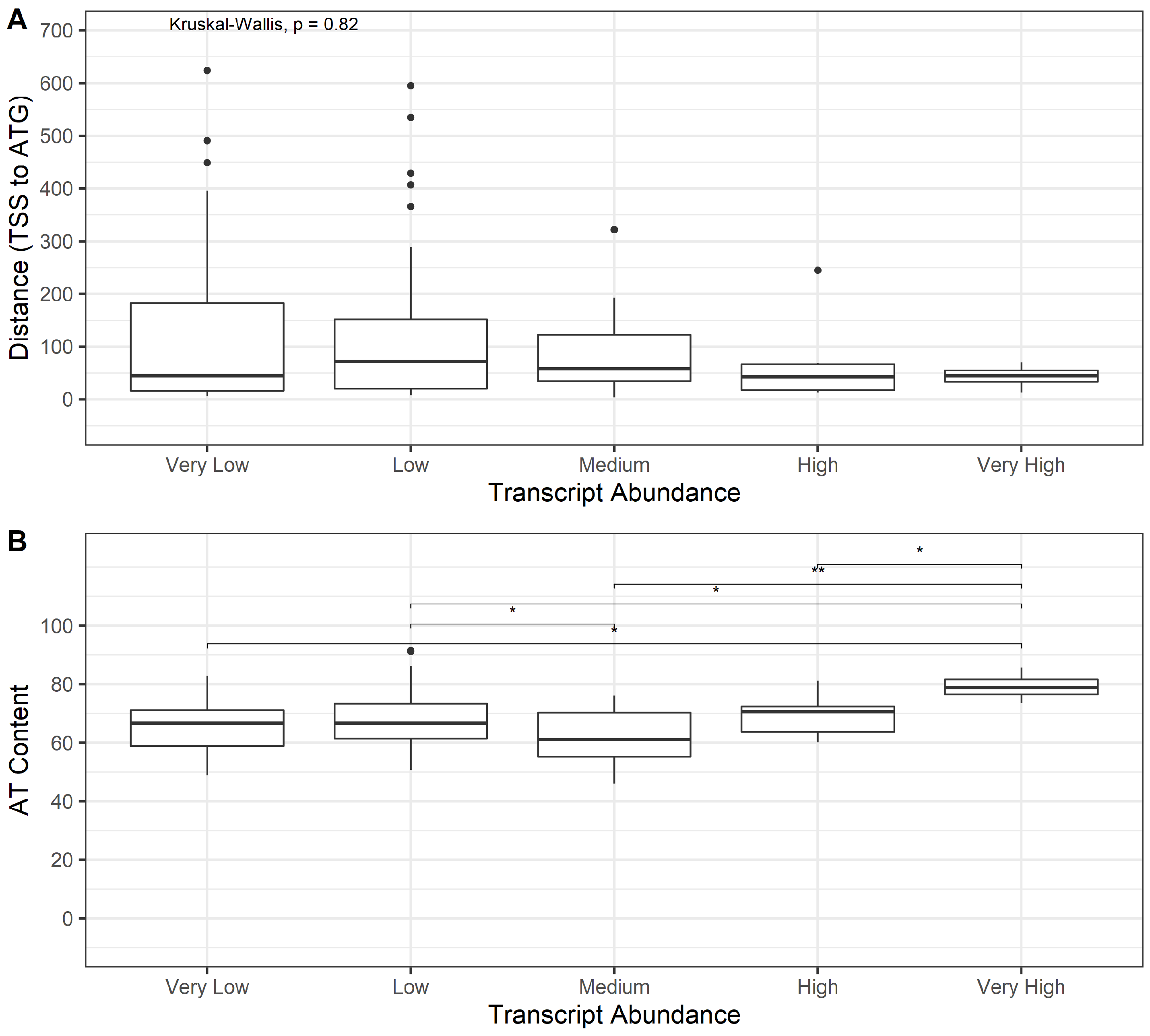
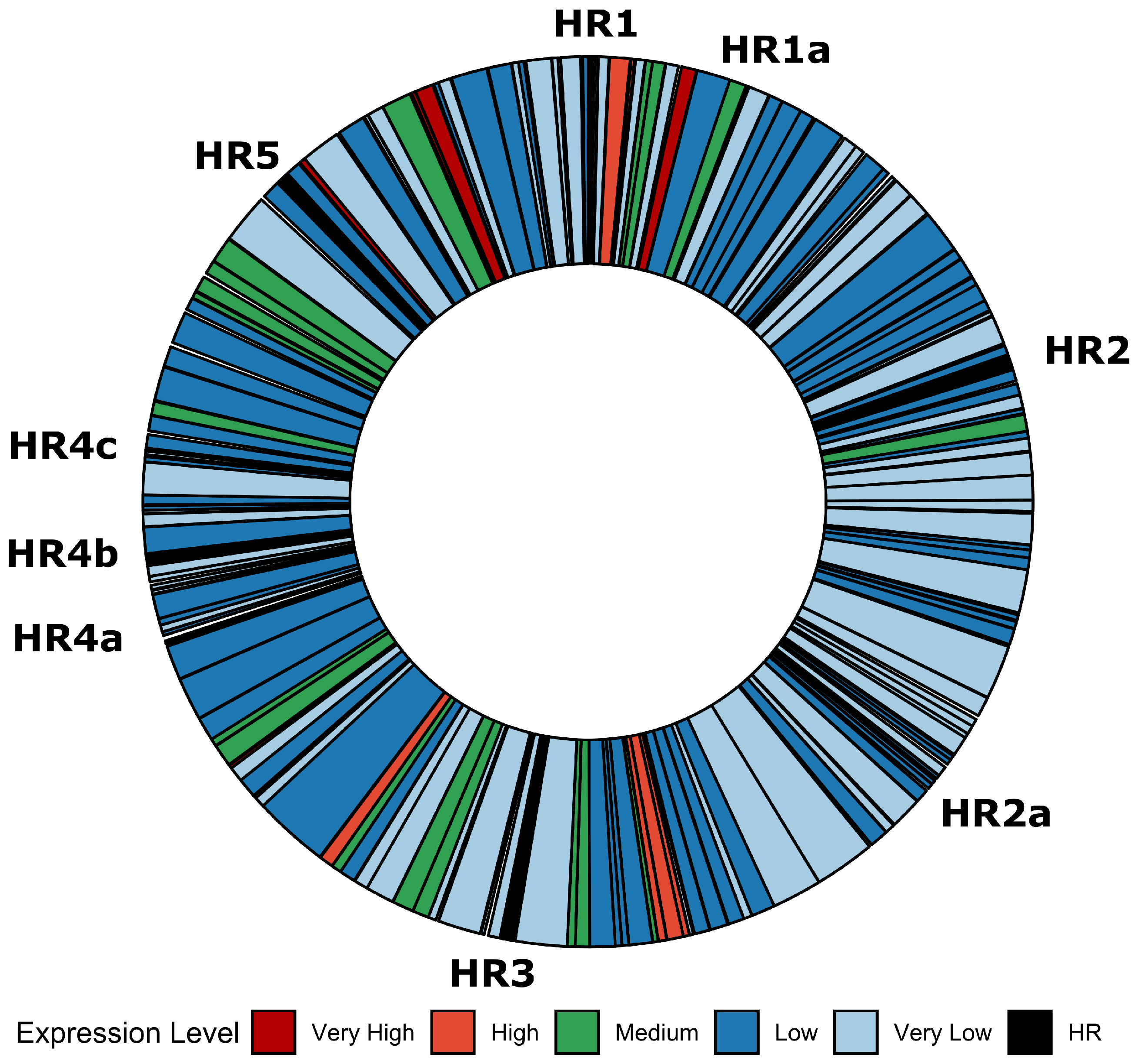
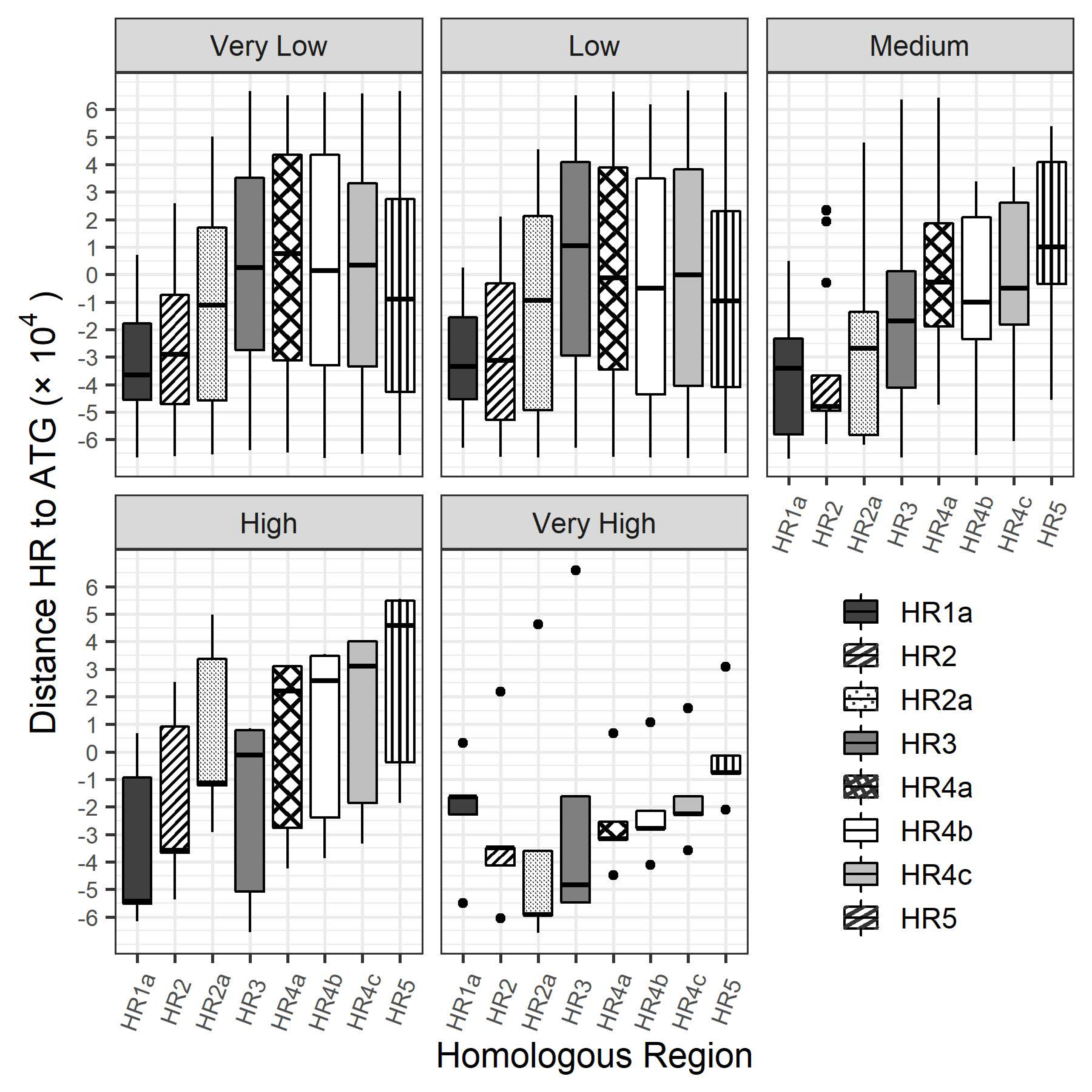
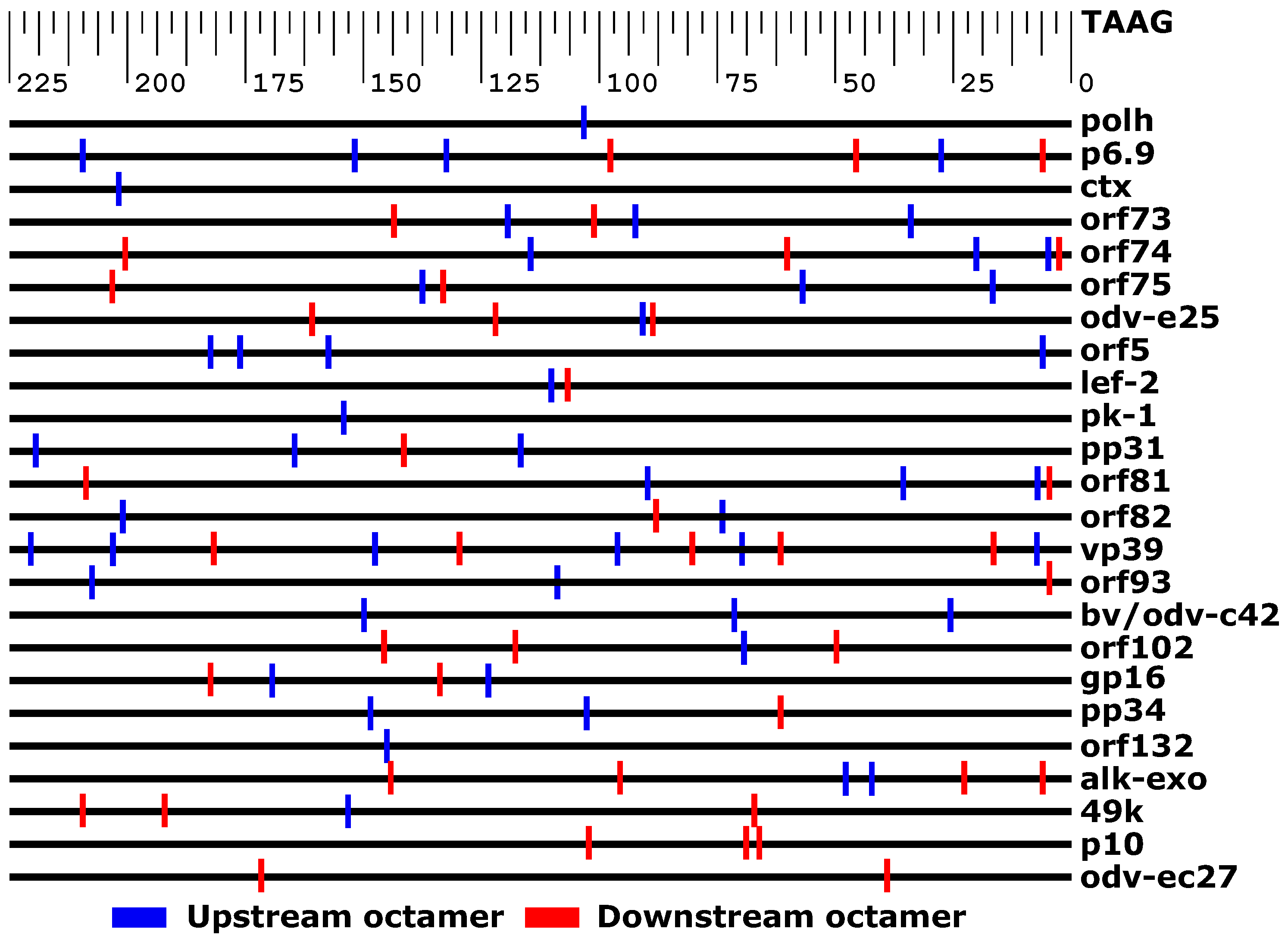
3.3. Bioinformatics Exploration of Sequence and Genomic Architecture Determinants That Contribute to Promoter Activity
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AcMNPV | Autographa californica multiple nucleopolyhedrovirus |
| au | arbitrary units |
| BV | budded virus |
| BEVS | baculovirus expression vectors system |
| rBEV | recombinant baculovirus expression vector |
| eGFP | enhanced green fluorescent protein |
| EPDA | end-point dilution assay |
| GFP | green fluorescent protein |
| hpi | hours post infection |
| IVT | infectious virus titer |
| KOV | knockout virus |
| MOI | multiplicity of infection |
| ORF | open reading frame |
| VLP | virus-like particle |
References
- Felberbaum, R.S. The baculovirus expression vector system: A commercial manufacturing platform for viral vaccines and gene therapy vectors. Biotechnol. J. 2015, 10, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Creech, C.B.; Walker, S.C.; Samuels, R.J. SARS-CoV-2 Vaccines. JAMA 2021, 325, 1318–1320. [Google Scholar] [CrossRef]
- Sokolenko, S.; George, S.; Wagner, A.; Tuladhar, A.; Andrich, J.M.S.; Aucoin, M.G. Co-expression vs. co-infection using baculovirus expression vectors in insect cell culture: Benefits and drawbacks. Biotechnol. Adv. 2012, 30, 766–781. [Google Scholar] [CrossRef] [PubMed]
- Oers, M.M.v.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus-insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Solís, M.; Herrero, S.; Targovnik, A.M. Engineering of the baculovirus expression system for optimized protein production. Appl. Microbiol. Biotechnol. 2019, 103, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Higgins, M.K.; Demir, M.; Tate, C.G. Calnexin co-expression and the use of weaker promoters increase the expression of correctly assembled Shaker potassium channel in insect cells. Biochim. Biophys. Acta (BBA) Biomembr. 2003, 1610, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Jarvis, D.L. Utility of temporally distinct baculovirus promoters for constitutive and baculovirus-inducible transgene expression in transformed insect cells. J. Biotechnol. 2013, 165, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, R.; Ernst, W.; Doblhoff-Dier, O.; Sara, M.; Katinger, H. Expression of foreign proteins on the surface of Autographa californica nuclear polyhedrosis virus. BioTechniques 1997, 22, 730–735. [Google Scholar] [CrossRef]
- Palomares, L.A.; López, S.; Ramírez, O.T. Strategies for manipulating the relative concentration of recombinant rotavirus structural proteins during simultaneous production by insect cells. Biotechnol. Bioeng. 2002, 78, 635–644. [Google Scholar] [CrossRef]
- Aucoin, M.G.; Perrier, M.; Kamen, A.A. Production of adeno-associated viral vectors in insect cells using triple infection: Optimization of baculovirus concentration ratios. Biotechnol. Bioeng. 2006, 95, 1081–1092. [Google Scholar] [CrossRef]
- Palomares, L.A.; Mena, J.A.; Ramírez, O.T. Simultaneous expression of recombinant proteins in the insect cell-baculovirus system: Production of virus-like particles. Methods 2012, 56, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Roldão, A.; Vieira, H.L.; Alves, P.M.; Oliveira, R.; Carrondo, M.J. Intracellular dynamics in rotavirus-like particles production: Evaluation of multigene and monocistronic infection strategies. Process Biochem. 2006, 41, 2188–2199. [Google Scholar] [CrossRef]
- Kato, T.; Manohar, S.L.; Kanamasa, S.; Ogata, M.; Park, E.Y. Improvement of the transcriptional strength of baculovirus very late polyhedrin promoter by repeating its untranslated leader sequences and coexpression with the primary transactivator. J. Biosci. Bioeng. 2012, 113, 694–696. [Google Scholar] [CrossRef] [PubMed]
- López-Vidal, J.; Gómez-Sebastián, S.; Sánchez-Ramos, I.; Escribano, J.M. Characterization of a Trichoplusia ni hexamerin-derived promoter in the AcMNPV baculovirus vector. J. Biotechnol. 2013, 165, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Solís, M.; Gómez-Sebastián, S.; Escribano, J.M.; Jakubowska, A.K.; Herrero, S. A novel baculovirus-derived promoter with high activity in the baculovirus expression system. PeerJ 2016, 4, e2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Sebastián, S.; López-Vidal, J.; Escribano, J.M. Significant productivity improvement of the baculovirus expression vector system by engineering a novel expression cassette. PLoS ONE 2014, 9, e96562. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26, D49–D53. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.R.; Zhong, S.; Fei, Z.; Hashimoto, Y.; Xiang, J.Z.; Zhang, S.; Blissard, G.W. The transcriptome of the baculovirus Autographa californica multiple nucleopolyhedrovirus in Trichoplusia ni cells. J. Virol. 2013, 87, 6391–6405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayres, M.D.; Howard, S.C.; Kuzio, J.; Lopez-Ferber, M.; Possee, R.D. The complete DNA sequence of Autographa californica nuclear polyhedrosis virus. Virology 1994, 202, 586–605. [Google Scholar] [CrossRef]
- Karasawa, S.; Araki, T.; Yamamoto-Hino, M.; Miyawaki, A. A Green-emitting fluorescent protein from Galaxeidae coral and its monomeric version for use in fluorescent labeling. J. Biol. Chem. 2003, 278, 34167–34171. [Google Scholar] [CrossRef]
- Bruder, M.R.; Walji, S.D.; Aucoin, M.G. Comparison of CRISPR—Cas9 Tools for Transcriptional Repression and Gene Disruption 589 in the BEVS. Viruses 2021, 13, 1925. [Google Scholar] [CrossRef] [PubMed]
- Yang, J. Small-Scale Production of Recombinant Proteins Using the Baculovirus Expression Vector System. Methods Mol. Biol. 1995, 1350, 225–239. [Google Scholar]
- Ahmed, M.; Kim, D.R. pcr: An R package for quality assessment, analysis and testing of qPCR data. PeerJ 2018, 6, e4473. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.L.; Salem, T.Z.; Turney, C.M.; Cheng, X.W. Strategy of the use of 28S rRNA as a housekeeping gene in real-time quantitative PCR analysis of gene transcription in insect cells infected by viruses. J. Virol. Methods 2010, 163, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Bodenhofer, U.; Bonatesta, E.; Horejš-Kainrath, C.; Hochreiter, S. msa: An R package for multiple sequence alignment. Bioinformatics 2015, 31, 3997–3999. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Cook, D.; Lawrence, M. ggbio: An R package for extending the grammar of graphics for genomic data. Genome Biol. 2012, 13, R77. [Google Scholar] [CrossRef] [Green Version]
- Thiem, S.M.; Miller, L.K. Identification, sequence, and transcriptional mapping of the major capsid protein gene of the baculovirus Autographa californica nuclear polyhedrosis virus. J. Virol. 1989, 63, 2008–2018. [Google Scholar] [CrossRef] [Green Version]
- Hou, D.; Chen, X.; Zhang, L.K. Proteomic Analysis of Mamestra Brassicae Nucleopolyhedrovirus Progeny Virions from Two Different Hosts. PLoS ONE 2016, 11, e0153365. [Google Scholar] [CrossRef] [Green Version]
- Moldován, N.; Tombácz, D.; Szűcs, A.; Csabai, Z.; Balázs, Z.; Kis, E.; Molnár, J.; Boldogkői, Z. Third-generation Sequencing Reveals Extensive Polycistronism and Transcriptional Overlapping in a Baculovirus. Sci. Rep. 2018, 8, 8604. [Google Scholar] [CrossRef] [Green Version]
- George, S.; Jauhar, A.M.; Mackenzie, J.; Kieβlich, S.; Aucoin, M.G. Temporal characterization of protein production levels from baculovirus vectors coding for GFP and RFP genes under non-conventional promoter control. Biotechnol. Bioeng. 2015, 112, 1822–1831. [Google Scholar] [CrossRef] [PubMed]
- Urabe, M.; Nakakura, T.; Xin, K.Q.; Obara, Y.; Mizukami, H.; Kume, A.; Kotin, R.M.; Ozawa, K. Scalable generation of high-titer recombinant adeno-associated virus type 5 in insect cells. J. Virol. 2006, 80, 1874–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Miller, L.K. Activation of baculovirus very late promoters by interaction with very late factor 1. J. Virol. 1999, 73, 3404–3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, T.D.; Miller, L.K. Mutational analysis of a baculovirus major late promoter. Gene 1994, 140, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Sokal, N.; Nie, Y.; Willis, L.G.; Yamagishi, J.; Blissard, G.W.; Rheault, M.R.; Theilmann, D.A. Defining the roles of the baculovirus regulatory proteins IE0 and IE1 in genome replication and early gene transactivation. Virology 2014, 468–470, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Gwak, W.S.; Bae, S.M.; Choi, J.B.; Han, B.K.; Woo, S.D. Increased productivity of the baculovirus expression vector system by combining enhancing factors. J. Asia-Pac. Entomol. 2018, 21, 1079–1084. [Google Scholar] [CrossRef]
- Koczka, K.; Peters, P.; Ernst, W.; Himmelbauer, H.; Nika, L.; Grabherr, R. Comparative transcriptome analysis of a Trichoplusia ni cell line reveals distinct host responses to intracellular and secreted protein products expressed by recombinant baculoviruses. J. Biotechnol. 2018, 270, 61–69. [Google Scholar] [CrossRef]
- Weyer, U.; Possee, R.D. Functional analysis of the p10 gene 5′ leader sequence of the Autographa californica nuclear polyhedrosis virus. Nucleic Acids Res. 1988, 16, 3635–3653. [Google Scholar] [CrossRef] [Green Version]
- Sano, K.I.; Maeda, K.; Oki, M.; Maéda, Y. Enhancement of protein expression in insect cells by a lobster tropomyosin cDNA leader sequence. FEBS Lett. 2002, 532, 143–146. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, Y.; Yao, L.; Hao, H.; Fu, X.; Yang, Z.; Du, E. Enhanced production of porcine circovirus type 2 (PCV2) virus-like particles in Sf9 cells by translational enhancers. Biotechnol. Lett. 2015, 37, 1765–1771. [Google Scholar] [CrossRef]
- Tiwari, P.; Saini, S.; Upmanyu, S.; Benjamin, B.; Tandon, R.; Saini, K.S.; Sahdev, S. Enhanced expression of recombinant proteins utilizing a modified baculovirus expression vector. Mol. Biotechnol. 2010, 46, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.R.; Chou, C.C.; Wu, T.Y.; Yuen, J.P.Y.; Chao, Y.C. Novel baculovirus DNA elements strongly stimulate activities of exogenous and endogenous promoters. J. Biol. Chem. 2002, 277, 5256–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkaiah, B.; Viswanathan, P.; Habib, S.; Hasnain, S.E. An Additional Copy of the Homologous Region (hr1) Sequence in the Autographa californica Multinucleocapsid Polyhedrosis Virus Genome Promotes Hyperexpression of Foreign Genes. Biochemistry 2004, 43, 8143–8151. [Google Scholar] [CrossRef] [PubMed]
- Rodems, S.M.; Friesen, P.D. The hr5 transcriptional enhancer stimulates early expression from the Autographa californica nuclear polyhedrosis virus genome but is not required for virus replication. J. Virol. 1993, 67, 5776–5785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possee, R.; Howard, S. Analysis of the polyhedrin gene promoter of the Autographa californica nuclear polyhedrosis virus. Nucleic Acids Res. 1987, 15, 10233–10248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peakman, T.C.; GiCharles, I.; Sydenham, M.A.; Gewert, D.R.; Page, M.J.; Makoff, A.J. Enhanced expression of recombinant proteins in insect cells using a baculovirus vector containing a bacterial leader sequence. Nucleic Acids Res. 1992, 20, 6111–6112. [Google Scholar] [CrossRef] [Green Version]
- Hills, D.; Crane-Robinson, C. Baculovirus expression of human basic fibroblast growth factor from a synthetic gene: Role of the Kozak consensus and comparison with bacterial expression. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 1995, 1260, 14–20. [Google Scholar] [CrossRef]
- Chang, M.J.; Kuzio, J.; Blissard, G.W. Modulation of Translational Efficiency by Contextual Nucleotides Flanking a Baculovirus Initiator AUG Codon. Virology 1999, 259, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Paulsson, J. Models of stochastic gene expression. Phys. Life Rev. 2005, 2, 157–175. [Google Scholar] [CrossRef]
- McLeay, R.C.; Bailey, T.L. Motif Enrichment Analysis: A unified framework and an evaluation on ChIP data. BMC Bioinform. 2010, 11, 165. [Google Scholar] [CrossRef] [Green Version]
- Ooi, B.G.; Rankin, C.; Miller, L.K. Downstream sequences augment transcription from the essential initiation site of a baculovirus polyhedrin gene. J. Mol. Biol. 1989, 210, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Rankin, C.; Ooi, B.G.; Miller, L.K. Eight base pairs encompassing the transcriptional start point are the major determinant for baculovirus polyhedrin gene expression. Gene 1988, 70, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Weyer, U.; Possee, R.D. Analysis of the promoter of the Autographa californica nuclear polyhedrosis virus p10 gene. J. Gen. Virol. 1989, 70 Pt 1, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Scheper, G.C.; Vries, R.G.; Broere, M.; Usmany, M.; Voorma, H.O.; Vlak, J.M.; Thomas, A.A. Translational properties of the untranslated regions of the p10 messenger RNA of Autographa californica multicapsid nucleopolyhedrovirus. J. Gen. Virol. 1997, 78 Pt 3, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Liu, A.; Weaver, R.F. Studies on the Control Region of the p10 Gene of the Autographa californica Nuclear Polyhedrosis Virus. J. Gen. Virol. 1989, 70, 1273–1279. [Google Scholar] [CrossRef]
- Yang, S.; Miller, L.K. Control of Baculovirus Polyhedrin Gene Expression by Very Late Factor 1. Virology 1998, 248, 131–138. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Promoter | Coordinates † | Predicted Promoter Motifs ‡ | Direction | Class |
|---|---|---|---|---|
| polh | 4428…4519 | TAAG | + | Very Late |
| p6.9 | 86,889…87,204 | CAGT, TAAG | − | Late |
| ctx | 2246…2447 | TAAG | − | Late |
| orf75 | 63,528…63,912 | TAAG | − | Late |
| vp39 | 76,578…77,103 | TAAG | − | Late |
| 39k/pp31 | 30,070…30,398 | TATA, CAGT, TAAG | − | Delayed Early |
| gp64 | 109,718…110,022 | TATA, CAGT, TAAG | − | Early/Late |
| 38k | 85,984…86,276 | TAAG | − | Late |
| p10 | 118,635…118,808 | TAAG | + | Very Late |
| Class | RPKM Range | ORFs |
|---|---|---|
| Very High | RPKM ≥ 50,000 | 5 |
| High | 20,000 ≥ RPKM < 50,000 | 6 |
| Medium | 10,000 ≥ RPKM < 20,000 | 17 |
| Low | 1000 ≥ RPKM < 10,000 | 65 |
| Very Low | RPKM < 1000 | 56 |
| Promoter | Rank (48 hpi) | ||||
|---|---|---|---|---|---|
| ORF | GFP | SEAP | |||
| RPKM | Protein | qPCR | Protein | qPCR | |
| polh | 1 | 1 | 2 | 2 | 2 |
| p6.9 | 2 | 2 | 1 | 1 | 1 |
| ctx | 3 | 5 | 4 | 5 | 7 |
| orf75 | 4 | 6 | 6 | 6 | 4 |
| vp39 | 5 | 4 | 5 | 3 | 3 |
| 39k | 6 | 3 | 3 | 4 | 5 |
| gp64 | 7 | 9 | 9 | 8 | 8 |
| 38k | 8 | 8 | 7 | 9 | 9 |
| p10 | n/a | 7 | 8 | 7 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruder, M.R.; Aucoin, M.G. Utility of Alternative Promoters for Foreign Gene Expression Using the Baculovirus Expression Vector System. Viruses 2022, 14, 2670. https://doi.org/10.3390/v14122670
Bruder MR, Aucoin MG. Utility of Alternative Promoters for Foreign Gene Expression Using the Baculovirus Expression Vector System. Viruses. 2022; 14(12):2670. https://doi.org/10.3390/v14122670
Chicago/Turabian StyleBruder, Mark R., and Marc G. Aucoin. 2022. "Utility of Alternative Promoters for Foreign Gene Expression Using the Baculovirus Expression Vector System" Viruses 14, no. 12: 2670. https://doi.org/10.3390/v14122670