
Contrasting the Practices of Virus Isolation and Characterization between the Early Period in History and Modern Times: The Case of Japanese Encephalitis Virus
Abstract
:1. Introduction to the Need of Re-Examining the History of Japanese Encephalitis
2. Method
3. Concept and Requirements for “First” Virus Isolation
3.1. Concept
3.2. Requirements for “First” Virus Isolation
3.2.1. Filterability and Submicroscopic Size
3.2.2. Absence of Replication in Culture Media
3.2.3. Identification of Optimal Source (s) in the Body of Infected Animal (or human) for Sampling, and Timing and/or Frequency of Sampling
3.2.4. Selection of Optimal Animal or Organ/Tissue Culture for Virus Propagation and Passage
3.2.5. Proof of Cause-Effect Relationship
3.2.6. Comparison with Known Viruses with Shared Characteristics
3.2.7. Preservation of Infectivity of Virus Isolate and Reproducibility
3.2.8. Proof of Vector-Borne Transmission
4. Brief History of the Early Occurrence of the Outbreaks of an Encephalitic Disease in Japan and Historical Accounts of the “First” Isolation of JEV
4.1. Brief History of Early Outbreaks
4.2. Etiology of JE
4.3. “First” Isolation of JEV in Japan

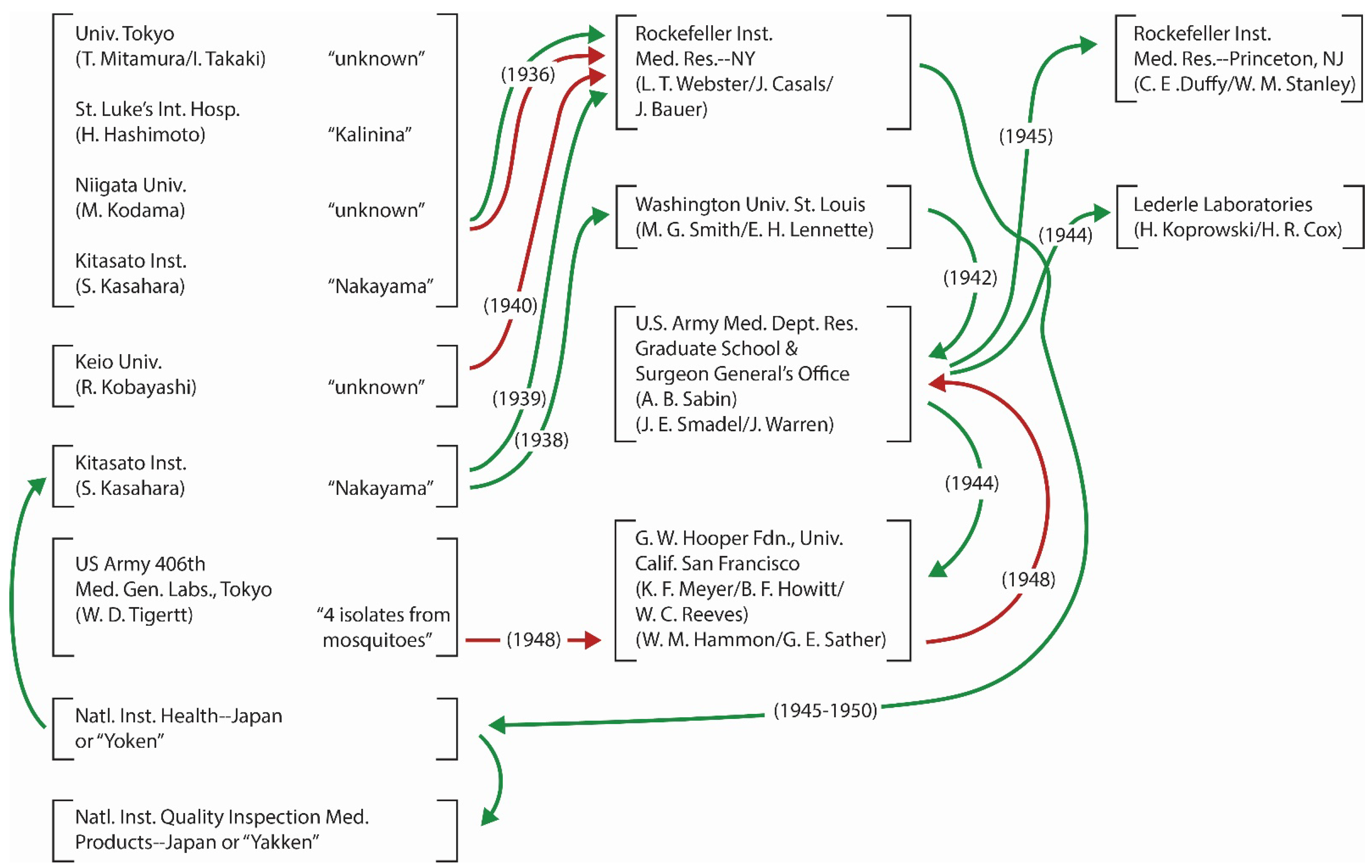{kind=link}
| 1924 | Ito et al. (1925) [49] Kaneko (1925) [50] Kaneko & Aoki (1928) [41] Kojima and Ono (1925) [51] Takagi (1925) [52] | Clarke & Casals (1965) [53] Erlander et al. (2009) [54] Le Flohic & Gonzales (2011) [55] Rivers (1927) [56] Tiroumourougane et al. (2001) [57] |
| 1933/1934 * | Hayashi (1934) [58] | Endy & Nisalak (2002) [59] Halstead & Jacobson (2003) [60] ** Hayashi (2022) [61] ** Holbrook (2017) [62] Huang (1982) [63] Kobayashi (1959) [64] Okubo (1953) [23] Rosen (1986) [65] |
| 1935/1936 * | Asami et al. (1935) [66] Hashimoto et al. (1936) [67] Kasahara et al. (1936a) [68] Kasahara et al. (1936b) [69] Kawamura et al. (1936) [70] Kobayashi et al. (1935) [71] Kudo et al. (1935) [72] Mitamura et al. (1935) [73] Mitamura et al. (1936) [74] Takagi (1935) [75] Takenouchi et al. (1935) [76] Taniguchi et al. (1936) [77] | Burke & Leake (1988) [78] European Centre for Disease Prevention & Control (Online Fact Sheets) (Year unspecified) [79] Innis (1995) [80] Karabatsos (1988) [81] Solomon et al. (2003) [82] |
4.4. Virus Exchange across the Pacific and Other Transfers of JEV Strains
4.5. The Twisted Background of Nakayama-NIH Designation
4.6. The Impacts of Increased Passage History and Change of Hands
4.7. Improvement in Virus Propagation and Storage
5. Early Characterization of JEV
5.1. Classification of JEV
5.2. Question over Overwintering and Persistence of JEV in Temperate Regions
5.3. Roles of Hosts
6. Communication and Information Retrieval
7. Difference in Conclusion among Molecular Studies as a Rich Source of New Ideas and Research Questions as well as Sources of Cautions to Be Exercised
7.1. Metagenomic (or Metatranscriptomic) Discovery of New Virus
7.2. Phylogenetic Identification of New Viruses and Phylogeography of Flaviviruses
7.3. Identification of the Hosts of Flaviviruses by Phylogenetics or Shared Molecular Determinants
7.4. Viral Replication Assay and Persistence of Viral RNA without a Recovery of Infectious Virus
8. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mackenzie, J.S.; Chua, K.B.; Daniels, P.W.; Eaton, B.T.; Field, H.E.; Hall, R.A.; Halpin, K.; Johansen, C.A.; Kirkland, P.D.; Lam, S.K.; et al. Emerging viral diseases of Southeast Asia and the Western Pacific. Emerg. Inf. Dis. 2001, 7, 497–504. [Google Scholar]
- Morita, K.; Nabeshima, T.; Buerano, C.C. Japanese encephalitis. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 441–452. [Google Scholar]
- Monath, T.P. Japanese encephalitis—A plague of the Orient. N. Eng. J. Med. 1988, 319, 641–643. [Google Scholar]
- Igarashi, A.; Tanaka, M.; Morita, K.; Takasu, T.; Ahmed, A.; Akram, D.S.; Wagar, M.A. Detection of West Nile and Japanese encephalitis viral genome sequences in cerebrospinal fluid from acute encephalitis cases in Karachi, Pakistan. Microbiol. Immunol. 1994, 38, 827–830. [Google Scholar]
- Ravanini, P.; Huhtamo, E.; Ilaria, V.; Crobu, M.G.; Nicosia, A.M.; Servino, L.; Rivasi, F.; Allegrini, S.; Miglio, U.; Magri, A.; et al. Japanese encephalitis virus RNA detected in Culex pipiens mosquitoes in Italy. Euro Surveill. 2012, 17, 20221. [Google Scholar]
- Simon Loriere, E.; Faye, Q.; Prot, M.; Fall, G.; Kipela, J.-M.; Fall, I.S.; Holmes, E.C.; Sakuntabhai, A.; Sall, A.A. Autochthonous Japanese encephalitis with yellow fever coinfection in Africa. N. Eng. J. Med. 2017, 376, 15. [Google Scholar]
- Van den Hurk, A.E.; Skinner, E.; Ritchie, S.A.; Mackenzie, J.S. The emergence of Japanese encephalitis virus in Australia in 2022: Existing knowledge of mosquito vectors. Viruses 2022, 14, 1208. [Google Scholar]
- Nabeshima, T.; Loan, H.T.K.; Inoue, S.; Sumiyoshi, M.; Haruta, Y.; Nga, P.T.; Huoung, V.T.Q.; del Carmen Parquet, M.; Hasebe, F.; Morita, K. Evidence of frequent introductions of Japanese encephalitis virus from south-coast Asia and continental East Asia to Japan. J. Gen. Virol. 2009, 90, 827–832. [Google Scholar]
- Yoshikawa, A.; Nabeshima, T.; Inoue, S.; Agoh, M.; Morita, K. Molecular and serological epidemiology of Japanese encephalitis virus (JEV) in a remote island of Western Japan: An implication of JEV migration over the East China Sea. Trop. Med. Health 2016, 44, 8. [Google Scholar]
- Arai, S.; Kuwata, R.; Higa, Y.; Maekawa, Y.; Tsuda, Y.; Roychoudhyry, S.; Bertuso, A.G.; Phong, T.V.; Yen, N.T.; Etoh, T.; et al. Two hidden taxa in the Japanese encephalitis vector mosquito Culex tritaeniorhynchus and the potential for long-distance migration from overseas to Japan. PLoS Negl. Trop. Dis. 2022, 16, e0010543. [Google Scholar]
- Sabin, A.B. Survey of knowledge and problems in field of arthropod-borne virus infections. Archiv. Ges. Virusforsch. 1959, 9, 1–10. [Google Scholar]
- Gao, X.-Y.; Liu, H.; Li, M.-H.; Fu, S.-H.; Liang, G. Insights into the evolutionary history of Japanese encephalitis virus (JEV) based on whole-genome sequences comprising the five genotypes. Virol. J. 2015, 12, 43. [Google Scholar]
- Kumar, K.; Arshad, S.S.; Selvarajah, G.T.; Abu, J.; Toung, O.P.; Abba, Y.; Yasmin, A.R.; Bande, F.; Sharma, R.; Ong, B.L. Japanese encephalitis in Malaysia: An overview and timeline. Acta Trop. 2018, 185, 219–229. [Google Scholar]
- Kuhn, J.H.; Bao, Y.-M.; Bavari, S.; Becker, S.; Bradfute, S.; Brister, J.R.; Bukreyev, A.A.; Chandran, K.; Davey, R.A.; Dolnik, O. Virus nomenclature below the species level: A standardized nomenclature for natural variants of viruses assigned to the family Filoviridae. Archiv. Virol. 2013, 158, 301–311. [Google Scholar]
- Stollar, V.; Thomas, V.L. An agent in the Aedes aegypti cell line (Peleg) which causes fusion of Aedes albopictus cells. Virology 1975, 64, 367–377. [Google Scholar]
- Kuno, G. Early history of laboratory breeding of Aedes aegypti (Diptera: Culicidae) focusing on the origins and use of selected strains. J. Med. Entomol. 2010, 47, 957–971. [Google Scholar]
- Cook, S.; Bennett, S.N.; Holmes, E.C.; De Chesse, R.; Moureau, G.; de Lamballerie, X. Isolation of a new strain of the flavivirus cell fusing agent virus in a natural mosquito population from Puerto Rico. J. Gen. Virol. 2006, 87, 735–748. [Google Scholar]
- Mokili, J.L.; Rohwer, F.; Dutilh, B.E. Metagenomics and future perspectives. I. Virus discovery. Curr. Opin. Virol. 2012, 2, 63–77. [Google Scholar]
- Calisher, C.H.; Horzinek, M.C. (Eds.) 100 Years of Virology; Springer: Wien, Austria, 1999. [Google Scholar]
- Murphy, F.A. The Foundations of Virology, 2nd ed.; American Society for Virology: Galveston, TX, USA, 2020; Available online at the University of Texas Medical Branch [UTMB] home page. Available online: https://asv.org/ (accessed on 9 October 2022).
- Cavaillon, J.-M. Le Mauvais Air: Il Était Une Fois Les Infections Et Les Combattants De L’ombre; EDP Sciences: Les Ulis Cedex A, France, 2022; p. 432. [Google Scholar]
- Wilkinson, L. The development of the virus concept as reflected in corpora of studies on individual pathogens. Med. Hist. 1974, 18, 211–221. [Google Scholar]
- Okubo, T. Propagation of Japanese encephalitis virus in chick embryo. Jpn. J. Microbiol. 1953, 8, 186–190. (In Japanese) [Google Scholar]
- Webster, L.T.; Fite, G.L. Experimental studies on encephalitis. J. Exp. Med. 1935, 61, 103–114. [Google Scholar]
- Kawakita, Y. In vitro propagation of Japanese encephalitis virus. Part I. Jikken Igaku 1939, 23, 1376–1392. (In Japanese) [Google Scholar]
- Shimagoshi, H. A method of mass propagating Japanese encephalitis virus. Osaka Igakukai Zasshi 1939, 38, 419–433. (In Japanese) [Google Scholar]
- Webster, L.T.; Fite, G.L.; Clow, A.D.; Muench, H. Experimental studies on encephalitis: IV. Specific inactivation of virus by sera from persons exposed to encephalitis, St. Louis type, 1933. with a note on the evaluation of the results of mouse tests of sera. J. Exp. Med. 1935, 62, 827–847. [Google Scholar]
- Mitamura, T.; Kitaoka, M.; Watanabe, S.; Okubo, K.; Tenjin, T. Immune response in Japanese encephalitis. Tokyo Iji Shinshi 1936, 3006, 3149–3156. (In Japanese) [Google Scholar]
- Frobisher, M. Complement fixation test in yellow fever. Proc. Soc. Exp. Biol. Med. 1929, 26, 846–848. [Google Scholar]
- Sawyer, W.A.; Lloyd, W. The use of mice in tests of immunity against yellow fever. J. Exp. Med. 1931, 54, 533–555. [Google Scholar]
- Casals, J.; Palacios, R. The complement fixation test in the diagnosis of virus infections of the central nervous system. J. Exp. Med. 1941, 74, 409–426. [Google Scholar]
- Hirota, M. Complement fixation test for epidemic Japanese encephalitis. Jpn. J. Microbiol. 1947, 2, 9–10. (In Japanese) [Google Scholar]
- Matsumoto, M. Recent research on Japanese encephalitis in the United States. Jpn. J. Microbiol. 1948, 3, 59–70. (In Japanese) [Google Scholar]
- Hirst, G.K. The quantitative determination of influenza virus and antibodies by means of red blood cell agglutination. J. Exp. Med. 1942, 75, 49–64. [Google Scholar]
- Casals, J. Immunological relationships among central nervous system viruses. J. Exp. Med. 1944, 79, 341–359. [Google Scholar]
- Huddleston, J.H.; Elgin, W.F.; Gilliland, S.H.; Williams, A.W. Report of Committee on Standard Methods of Preparing Smallpox Vaccine. Am. J. Public Health 1910, 20, 690–703. [Google Scholar]
- Sawyer, W.A.; Lloyd, W.D.M.; Kitchen, S.F. The preservation of yellow fever virus. J. Exp. Med. 1929, 50, 1–13. [Google Scholar]
- Flosdorf, E.W.; Mudd, S. Procedure and approaches for preservation in “lyophile” form of serum and other biologicl substances. J. Immunol. 1935, 29, 389–425. [Google Scholar]
- Mitamura, T.; Kitaoka, M.; Mori, K.; Okubo, K. Isolation of the virus of Japanese encephalitis from mosquitoes caught in nature. Reports to the Ninth Meeting of the Committee on encephalitis. Tokyo Iji Shinshi 1938, 3076, 820–824. (In Japanese) [Google Scholar]
- Mitamura, T.; Kitaoka, M.; Mori, K.; Okubo, K.; Tenjin, T. Über den Infektionsmodus der japanischen epidemischen Enzephalitis mit besonderer Berücksichtingung der experimentaellen und epidemiiologischen beweise für die Krankheit-sübertragung durch Mücken. Trans. Soc. Pathol. Japan 1938, 28, 135–145. [Google Scholar]
- Kaneko, R.; Aoki, Y. Ergebnisse der Encephalitis epidemica in Japan. Erg. Inn. Med. Und Der Kinderheilkd. 1928, 34, 342–456. [Google Scholar]
- Kinoshita, N. Statistical records of the frequency of the encephalitic epidemic in the towns and cities in the region of Okayama-ken since 1927. Igakkai Zasshi 1943, 55, 124–140. (In Japanese) [Google Scholar]
- Kono, R.; Kim, K.-H. Comparative epidemiological features of Japanese encephalitis in the Republic of Korea, China (Taiwan) and Japan. Bull. World Health Organ. 1969, 40, 263–277. [Google Scholar]
- Kamimura, K. Why did outbreaks and incidences of malaria and Japanese encephalitis decline in Japan? Med. J. Toyama Phamaco-Med. Coll. 1998, 11, 1–7. (In Japanese) [Google Scholar]
- Wang, H.-Y.; Li, Y.-X.; Liang, X.-F.; Liang, G.-D. Japanese encephalitis in mainland China. Jpn. J. Infect. Dis. 2009, 62, 331–336. [Google Scholar]
- von Economo, C.A. Encephalitis lethergica. Wien Klin. Weshr. 1917, 30, 581–585. [Google Scholar]
- Takagi, I. On the etiology of encephalitis that occurred epidemically in Japan in 1924. Jpn. Med. World 1925, 5, 147. (In Japanese) [Google Scholar]
- Inada, R. Compte rendu des recherches sur l’encéphalite epidémique au Japan. Office Int. Hyg. Publ. 1937, 29, 1389–1401. [Google Scholar]
- Ito, T. Studiens über dem Erreger der Encephalitis epidemica (lethergica). Saikingaku Zasshi 1925, 349, 352. [Google Scholar]
- Kaneko, R. On the epidemic encephalitis outbreak which occurred in Japan. Jpn. Med. World 1925, 5, 237. (In Japanese) [Google Scholar]
- Kojima, M.; Ono, M. Über Untersuchungen über die Ätiologie der epidemischen Encephalitis. I. Eiseigaku Densenbyogaku Zasshi 1925, 20, 6. [Google Scholar]
- Takagi, I. Studien über den Erreger der Encephalitis epidemica bei der Epidemie 1924. I. Mitteilung. Nisshin Igaku 1925, 14, 12. [Google Scholar]
- Clarke, D.H.; Casals, J.; Arboviruses Group, B. Viral and Rickettsial Infection of Man, 4th ed.; Horsfall, F.L., Tamm, I., Eds.; J.B. Lippincott Co.: Philadelphia, PA, USA; Toronto, ON, Canada, 1965; pp. 606–658. [Google Scholar]
- Erlander, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar]
- Le Frohic, G.; Gonzales, J.-P. When Japanese encephalitis virus invaded Eastern Hemisphere-the history of the spread of virus genotypes. In Flavivirus Encephalitis; Rúžek, D., Ed.; Intechopen: London, UK, 2011; Chapter 4; pp. 405–426. [Google Scholar]
- Rivers, T.M. Filterable agents. J. Bacteriol. 1927, 14, 217–258. [Google Scholar]
- Tiroumourougane, S.V.; Raghava, P.; Srinivasan, S. Japanese viral encephalitis. Postgrad. Med. 2002, 78, 205–215. [Google Scholar]
- Hayashi, M. Transmission of the encephalitis epidemica virus to monkeys. Proc. Imp. Acad. Tokyo 1934, 10, 41–44. [Google Scholar]
- Endy, T.P.; Nisalak, A. Japanese encephalitis virus: Ecology and epidemiology. Curr. Top. Microbiol. Immunol. 2002, 267, 11–48. [Google Scholar]
- Halstead, S.B.; Jacobson, J. Japanese encephalitis. Adv. Virus Res. 2003, 61, 103–138. [Google Scholar]
- Hayashi, M. Recent status of the epidemiology of flaviviral infections. Neuroinfection 2022, 27, 104–110. (In Japanese) [Google Scholar]
- Holbrook, H.R. Historical perspectives on flavivirus research. Viruses 2017, 9, 97. [Google Scholar]
- Huang, C.H. Studies of Japanese encephalitis in China. Adv. Virus Res. 1982, 27, 71–101. [Google Scholar]
- Kobayashi, T. Antigenic difference between two derivatives of Nakayama strain of Japanese encephalitis virus. Uirusu 1959, 9, 475–482. (In Japanese) [Google Scholar]
- Rosen, L. The natural history of Japanese encephalitis virus. Annu. Rev. Microbiol. 1986, 40, 395–414. [Google Scholar]
- Asami, S.; Oshiro, T.; Nakanoin, T. Searching the etiologic agent of Japanese encephalitis. Tohoku Ikai-Shi 1936, 19, 174–192. (In Japanese) [Google Scholar]
- Hashimoto, H.; Kudo, M.; Uraguchi, K. Experiences in the summer epidemic of acute encephalitis in Tokyo. J. Am. Med. Assoc. 1936, 106, 1266–1268. [Google Scholar]
- Kasahara, S.; Ueda, M.; Hamano, R.; Yamada, R.; Okamoto, Y.; Kohno, M. Experimental studies on epidemic encephalitis. 2. Further studies on epidemic encephalitis. 2. Further transmission test of the Japanese encephalitis. I. On the virus isolated later. Kitasato Arch. Exp. Med. 1936a, 13, 248–268. [Google Scholar]
- Kasahara, S.; Ueda, M.; Okamoto, Y.; Yoshida, S.; Hamano, R.; Yamada, R. Experimental studies on epidemic encephalitis. I. Transmission test of the Japanese encephalitis in 1935 and some characteristics of the infectious agent. Kitasato Arch. Exp. Med. 1936b, 13, 48–65. [Google Scholar]
- Kawamura, R.; Kodama, M.; Ito, T.; Yasaki, T.; Kobayakawa, Y. Epidemic encephalitis in Japan. The causative agent compared with that in the St. Louis epidemic. Arch. Pathol. 1936, 22, 510–523. [Google Scholar]
- Kobayashi, R.; Kobayashi, E.; Ando, K.; Nogi, M.; Fukutome, I.; Momose, E.; Ikegaki, R.; Kaibara, T.; Horimichi, N.; Miyoshi, S.; et al. Research on the etiologic agent of epidemic encephalitis that prevailed in Tokyo and Yokohama areas in 1935–1936. Tokyo Iji Shinshi 1935, 2958, 3127–3129. (In Japanese) [Google Scholar]
- Kudo, M.; Uraguchi, K.; Okazaki, M.; Hashimoto, M. A Japanese encephalitis outbreak in Tokyo area in 1935: The etiologic agent. Nihon Iji Shimpo 1935, 686, 3273–3283. (In Japanese) [Google Scholar]
- Mitamura, T.; Kitaoka, M.; Watanabe, T.; Okubo, K.; Tenjin, S.; Yamada, S.; Mori, K.; Asada, T. Epidemic encephalitis (summer encephalitis). I. Animal experiments and mosquito transmission of the etiologic agent. Kansai Iji 1935, 260, 3–4. (In Japanese) [Google Scholar]
- Mitamura, T.; Hazato, H.; Kitaoka, M. Comparison of Japanese encephalitis virus and St. Louis encephalitis virus and a possibility of the involvement of multiple etiologic agents. Tokyo Iji Shimpo 1936, 3006, 3157–3161. (In Japanese) [Google Scholar]
- Takagi, I. Experimental infection of laboratory animals with an isolated agent from an encephalitis patient. Tokyo Iji Shinshi 1935, 2958, 3129–3131. (In Japanese) [Google Scholar]
- Takenouchi, M.; Sato, J.; Yamamoto, I.; Fujimori, M.; Yamamoto, M.; Yoda, K.; Nagahama, R.; Takizawa, M. Research on the etiologic agent of epidemic encephalitis. Tokyo Iji Shinshi 1935, 2957, 3060–3062. (In Japanese) [Google Scholar]
- Taniguchi, T.; Hosokawa, N.; Kuga, S. A virus isolated in 1935 epidemic of summer encephalitis of Japan. Jpn. Exp. Med. 1936, 14, 185–196. (In Japanese) [Google Scholar]
- Burke, D.S.; Leake, C.J. Japanese encephalitis. In The Arboviruses: Epidemiology and Ecology; Monath, T.P., Ed.; CRC Press: Boca Raton, FL, USA, 1988; Volume 3, pp. 63–92. [Google Scholar]
- European Centre for Disease Prevention and Control. Fact Sheet about Japanese Encephalitis. Available online: https://www.ecdc.europa.eu/en/japanese-encephalitis/facts (accessed on 9 October 2022).
- Innis, B.L. Japanese encephalitis. In Exotic Viral Infections; Porterfield, J.S., Ed.; Chapman & Hall: London, UK, 1995; pp. 147–174. [Google Scholar]
- Karabatsos, N. (Ed.) International Catalogue of Arboviruses including Certain Other Viruses of Vertebrates, 3rd ed.; American Society of Tropical Medicine and Hygiene: San Antonio, TX, USA, 1985; p. 1147. [Google Scholar]
- Solomon, T.; Ni, H.-L.; Beasley, D.W.C.; Skkelenkaap, M.; Cardosa, M.J.; Barrett, A.D.T. Origin and evolution of Japanese encephalitis virus in Southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar]
- Theiler, M.; Downs, W.G. The Arthropod-Borne Viruses of Vertebrates; Yale University Press: New Haven, CT, USA, 1973; p. 578. [Google Scholar]
- Duffy, C.E.; Stanley, W.M. Studies on the biochemical, biophysical, and immunogenic properties of Japanese B type encephalitis virus and vaccine. J. Exp. Med. 1945, 82, 385–410. [Google Scholar]
- Hammon, W.M.; Tigertt, W.D.; Sather, G.; Schenker, H. Isolations of Japanese B encephalitis virus from naturally infected Culex tritaeniorhynchus collected in Japan. Am. J. Hyg. 1948, 50, 51–56. [Google Scholar]
- Koprowski, H.; Cox, H.R. Propagation of Japanese B encephalitis virus in the developing chick embryos. J. Immunol. 1946, 52, 171–186. [Google Scholar]
- Kobayashi, R.; Ando, K.; Nogi, M.; Fukutomi, I.; Momose, E.; Ikegaki, R.; Kaibara, T.; Hori, M.; Miyoshi, S.; Nakanishi, R.; et al. Investigation of the etiologic agent involved in the epidemic encephalitis in Tokyo and Yokohama in 1935–1936. Tokyo Iji Shinshi 1939, 3029, 1151–1155. (In Japanese) [Google Scholar]
- Kawamura, R.; Kodama, M.; Ito, T.; Yazaki, T.; Kohayagawa, K. Investigation of the encephalitis outbreak in 1935: Part I. Tokyo Iji Shinshi 1935, 2955, 2665–2668. (In Japanese) [Google Scholar]
- Kobayashi, Y.; Hasegawa, H.; Oyama, T.; Tamoi, T.; Kusaba, T. Antigenic analysis of Japanese encephalitis virus by using monoclonal antibodies. Infect. Immun. 1984, 44, 117–123. [Google Scholar]
- Japanese Society of Microbiology. 50th Anniversary panel discussion: Reminiscence of the past events and progresses. Jpn. J. Bacteriol. 1979, 33, 423–429. (In Japanese) [Google Scholar]
- Sabin, A.B. The St. Louis and Japanese B types of epidemic encephalitis. J. Am. Med. Assoc. 1943, 122, 477–486. [Google Scholar]
- Sabin, A.B.; Tigertt, W.D.; Ando, K.; Berge, T.O.; Blender, J.X.; Burns, K.F.; Kitaoka, M.; Kitayama, M.; Satterwhite, J.P. Evolution of Japanese B encephalitis vaccine.1. General background and methods. Am. J. Hyg. 1957, 63, 217–227. [Google Scholar]
- Webster, L.T. Japanese B encephalitis virus: Its differentiation from St. Louis encephalitis virus and its relationship to louping ill virus. J. Exp. Med. 1938, 67, 609–618. [Google Scholar]
- Smadel, J.R.; Randall, R.; Warren, J. Preparation of Japanese encephalitis vaccine. U.S. Army Med. Dept. Bull. 1947, 7, 963–973. [Google Scholar]
- Warren, J.; Hough, R.G. A vaccine against Japanese B encephalitis prepared from infected chick embryos. Proc. Soc. Exp. Biol. Med. 1946, 61, 109–113. [Google Scholar]
- Smithburn, K.C. Differentiation of the West Nile virus from the viruses of St. Louis and Japanese B encephalitis. J. Immunol. 1942, 44, 25–31. [Google Scholar]
- Haagen, E.; Crodel, B. Untersuchung über das japonische Encephalitivirus. Züchtungs Imunitäts und Neutralisationsversuche. Zbl. Bakt. I. Orig. 1938, 142, 269–279. [Google Scholar]
- Pette, H. Gibt es in Deutschland eine Enzephalitis von Charakter der Encephalitis japonica? Münchener Med. Wochenschr. 1938, 85, 1137–1140. [Google Scholar]
- Smadel, J.E. The Viral and Rickettsial registry, USA. Ann. N. Y. Acad. Sci. 1953, 56, 612–614. [Google Scholar]
- Paterson, P.Y.; Ley, H.L.; Wisseman, C.L.; Pond, W.L.; Smadel, J.E.; Diercks, F.H.; Hetherington, H.D.G.; Lancaster, W.E. Japanese encephalitis in Malay. I. Isolation of virus and serological evidence of human and equine infections. Am. J. Trop. Med. Hyg. 1952, 66, 320–330. [Google Scholar]
- Hale, J.H.; Lim, K.A.; Chee, P.H. Japanese type B encephalitis in Malaya. Ann. Trop. Med. 1952, 46, 220–226. [Google Scholar]
- Okuno, T.; Okada, T.; Kondo, A.; Suzuki, M.; Kobayashi, M.; Oya, A. Immunotyping of different strains of Japanese encephalitis virus by antibody-absorption, haemagglutionation-inhibition and complement-fixation tests. Bull. World Health Org. 1969, 38, 547–563. [Google Scholar]
- US-NIH. Minimum Requirement: Japanese Encephalitis Vaccine. Issued 5 February, 1945 and Japanese Encephalitis Vaccine Chick Embryo Type, Dried; Federal Security Agency, National Institute of Health: Bethesda, MD, USA, Issued 7 August 1947.
- Yamauchi, T. Immunological analysis of Japanese encephalitis virus using anti-Kamiyama monoclonal antibodies. Kansensho Gakkai Zasshi 1989, 63, 387–399. (In Japanese) [Google Scholar]
- Kovalev, S.Y.; Mukhacheva, T.A.; Kokorev, V.S.; Belyaeva, I.V. Tick-borne encephalitis virus: Reference strain Sofjin and problem of its authenticity. Virus Genes 2011, 44, 217–224. [Google Scholar]
- Davis, E.H.; Beck, A.S.; Li, L.; White, M.M.; Greenberg, M.B.; Thompson, J.K.; Wilden, S.G.; Barrett, A.D.T.; Bourne, N. Japanese encephalitis virus live attenuated vaccine strains display altered immunogenicity, virulence and genetic diversity. Npj Vaccine 2021, 6, 112. [Google Scholar]
- Taylor, R.M. Catalogue of Arthropod-Borne Viruses of the World: A Collection of Data on Registered Arthropod-Borne Animal Viruses, Public Health Service Publication No. 1760, 1st ed.; U.S. Government Printing Office: Washington, DC, USA, 1967.
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic relationships between flaviviruses as determined by cross- neutralization tests with polyclonal antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar]
- Lee, H.W. Study on overwintering mechanism of Japanese encephalitis virus in Korea. J. Korean Med. Assoc. 1971, 14, 871–878. [Google Scholar]
- Hayashi, K.; Mifune, K.; Shichijyo, A.; Matsuo, S.; Makino, Y.; Akashi, M.; Wada, Y.; Oda, T.; Mogi, M.; Mori, A. Ecology of Japanese encephalitis virus in Japan III. The results of investigations in Amami Island, southern part of Japan from 1973 to 1975. Trop. Med. 1975, 17, 129–142. [Google Scholar]
- Ura, M. Ecology of encephalitis in Okinawa, Japan. I. The investigation on pig and mosquito infection of the virus in Okinawa Island from 1966 to 1976. Trop. Med. 1976, 18, 151–163. [Google Scholar]
- Takashima, I.; Watanabe, T.; Ouchi, N.; Hashimoto, N. Ecological studies of Japanese encephalitis virus in Hokkaido: Interepidemic outbreaks of swine abortion and evidence for the virus to overwinter locally. Am. J. Trop. Med. Hyg. 1988, 38, 420–427. [Google Scholar]
- Nga, P.T.; del Carmen Parquet, M.; Cuong, V.D.; Ma, S.P.; Hasebe, F.; Inoue, S.; Makino, Y.; Takagi, M.; Nam, V.S.; Morita, K. Shift on Japanese encephalitis virus (JEV) genotype circulating in northern Vietnam: Implication for frequent introductions of JEV from Southeast Asia to East Asia. J. Gen. Virol. 2004, 85, 1625–1631. [Google Scholar]
- Schuh, A.J.; Ward, M.J.; Brown, A.J.L.; Barrett, A.D.T. Phylogeography of Japanese encephalitis virus: Genotype is associated with climate. PLoS Negl. Trop. Dis. 2013, 7, e2411. [Google Scholar]
- Hodes, H.L. Experimental transmission of Japanese B encephalitis by mosquitoes and mosquito larvae. Johns Hopkins Hosp. Bull. 1946, 79, 358–360. [Google Scholar]
- Shiroi, N.; Ando, K.; Sato, H.; Okubo, K.; Nakayama, T.; Ichikawa, O.; Yamada, M. Investigation of the etiology of epidemic encephalitis in horses. Jikken Igakushi 1937, 21, 117–146. (In Japanese) [Google Scholar]
- Tabuchi, E.; Hosoda, T.; Akiyama, Y.; Narita, R. Studies on Japanese B encephalitis and abortion. Kitasato Arch. Exp. Med. 1951, 23. Available online: https://www.cabdirect.org/cabdirect/welcome/?target=%2fcabdirect%2fabstract%2f19512702337 (accessed on 9 October 2022).
- Mitamura, T.; Kitaoka, M.; Mori, K.; Kobayashi, E.; Okubo, K.; Tenjin, S. Evidence of inapparent infection with Japanese B encephalitis virus in so-called normal persons and animals: Abortive form of Japanese B encephalitis. Tokyo Iji Shinshi 1938, 62, 779–789. (In Japanese) [Google Scholar]
- Buescher, E.L.; Scherer, W.F.; McClure, H.E.; Moyer, J.T.; Rosenberg, M.Z.; Yoshii, M.; Okada, Y. Ecologic studies of Japanese encephalitis virus in Japan. IV. Avian infection. Am. J. Trop. Med. Hyg. 1959, 8, 678–688. [Google Scholar]
- Buescher, E.L.; Scherer, W.F. Ecologic studies of Japanese encephalitis virus in Japan. IX. Epidemiologic correlations and conclusions. Am. J. Trop. Med. Hyg. 1959, 8, 719–722. [Google Scholar]
- Shimizu, T.; Kawakami, Y. Studies on swine stillbirth, especially on its relation to Japanese encephalitis (In Japanese). Rep. Natl. Inst. Anim. Health Jpn. 1949, 22, 117–128. (In Japanese) [Google Scholar]
- Konno, J.; Endo, K.; Agatsuma, H.; Ishida, N. Cyclic outbreaks of Japanese encephalitis among pigs and humans. Am. J. Epidemiol. 1966, 84, 292–300. [Google Scholar]
- Burns, K.F. Congenital Japanese B encephalitis infection of swine. Proc. Soc. Exp. Biol. Med. 1950, 75, 621–625. [Google Scholar]
- Hale, J.H.; Lim, K.A.; Colless, D.H. Investigation of domestic pigs and a potential reservoir as potential reservoir of Japanese encephalitis virus in Singapore Is. Ann. Trop. Med. Parasitol. 1959, 51, 374–379. [Google Scholar]
- Hurlbut, H.S. The pig-mosquito cycle of Japanese encephalitis virus in Taiwan. J. Med. Entomol. 1964, 1, 301–307. [Google Scholar]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, F.H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar]
- Howerth, E.W.; Stallknecht, D.E.; Dornng, M.; Pisell, T.; Clarke, G.R. Experimental vesicular stomatitis in swine: Effects of route of inoculation and steroid treatment. J. Vet. Diagn. Invest. 1997, 9, 136–142. [Google Scholar]
- Andeson, E.C.; Hutchings, G.H.; Mukarati, N.; Wilkinson, P.J. African swine fever virus infection of the bushpig (Potomochoerus porces) and its significance in the epidemiology of the disease. Vet. Microbiol. 1998, 62, 1–15. [Google Scholar]
- Kuno, G. Transmission of arboviruses without involvement of arthropod vectors. Acta Virol. 2001, 45, 139–150. [Google Scholar]
- Diallo, A.O.; Chevalier, V.; Cappelle, J.; Duong, V.; Fontenille, D.; Duboz, R. How much does direct transmission between pigs contribute to Japanese encephalitis virus circulation? A modelling approach in Cambodia. PLoS ONE 2018, 13, e0201209. [Google Scholar]
- Xu, C.-X.; Zhang, W.-J.; Pan, Y.-F.; Wang, G.-W.; Yin, Q.-K.; Fu, S.-H.; Li, F.; He, Y.; Xu, S.-T.; Wang, Z.-H. A bibliometric analysis of global research on Japanese encephalitis from 1934 to 2020. Front. Cell. Infect. Microbiol. 2022, 12, 833701. [Google Scholar]
- Oliveira, A.R.S.; Strate, E.; Etcherry, L.; Cohnstaedt, L.W.; McVey, D.S.; Piaggig, J.; Cernicchiara, N. Assessment of data on vector and host competence for Japanese encephalitis virus: A systematic review of the literature. Prev. Vet. Med. 2018, 154, 71–89. [Google Scholar]
- Vasilakis, N.; Tesh, R.B.; Popov, V.L.; Widen, S.G.; Wood, T.G.; Forester, N.L.; Gonzalez, J.P.; Saluzzo, J.F.; Alkhovsky, S.; Lam, S.K.; et al. Exploiting the legacy of the arbovirus hunters. Viruses 2019, 11, 471. [Google Scholar]
- Mulvey, P.; Duong, V.; Boyer, S.; Burgess, G.; Williams, D.T.; Dussart, P.; Hovwood, P.F. The ecology and evolution of Japanese encephalitis virus. Pathogens 2021, 10, 1534. [Google Scholar]
- Patel, J.P.; Verma, K.; Singh, V. Japanese encephalitis (JE): A curse for people living in Uttar Pradesh, India. J. Vaccines Immunol. 2021, 7, 36–40. [Google Scholar]
- Zlobin, V.I.; Pogodina, V.V.; Kahl, O. A brief history of the discovery of tick-borne encephalitis virus in the late 1930s (based on reminiscences of members of the expeditions, their colleagues and relatives). Ticks Tick-Borne Dis. 2017, 8, 813–820. [Google Scholar]
- Delwart, E.L. Viral metagenomics. Rev. Med. Virol. 2007, 17, 115–131. [Google Scholar]
- Calisher, C.H.; Blair, C.D.; Bowen, M.D.; Casals, J.; Drebot, M.A.; Henchal, E.A.; Karabatsos, N.; LeDuc, J.W.; Repik, P.M.; Roehrig, J.T.; et al. Identification of arboviruses and certain rodent-borne viruses: Reevaluation of the paradigm. Emerg. Infect. Dis. 2001, 7, 756–758. [Google Scholar]
- Wimmer, E. The test tube synthesis of a chemical called poliovirus. EMBO Rep. 2006, 7, S3–S9. [Google Scholar]
- Garfinkel, M.; Endy, D.; Epstein, G.; Friedman, R. Synthetic genomics: Options for governance. Biosecur. Bioterr. 2007, 5, 359–362. [Google Scholar]
- Atlas, R. Statement on scientific publication and security. Nature 2003, 421, 771. [Google Scholar]
- Cobbin, J.C.A.; Charon, J.; Harvey, E.; Holmes, E.C.; Mahar, J.E. Current challenges to virus discovery by metatranscriptomics. Curr. Top. Virol. 2021, 51, 48–55. [Google Scholar]
- Gorbalenya, A.E.; Lauber, C. Bioionformatics of virus taxonomy: Foundations and tools for developing sequence-based hierarchial classification. Curr. Opin. Virol. 2022, 52, 48–56. [Google Scholar]
- Gorbalenya, A.E.; Lauber, C. Phylogeny of viruses. Ref. Modul. Biomed. Sci. 2017, B978-0-12-801238-3.95723-4. Available online: https://www.sciencedirect.com/science/article/pii/B9780128012383957234?via%3Dihub (accessed on 9 October 2016). [CrossRef]
- Shapiro, J.T.; Viquez-R, L.; Leopardi, S.; Vicente-Santos, A.; Mendenhall, I.H.; Frick, W.F.; Kading, R.C.; Medellín, R.A.; Racey, P.; Kingston, T. Setting the terms for zoonotic diseases: Effective communication for research, conservation, and public policy. Viruses 2021, 13, 1356. [Google Scholar]
- Mohammed, M.A.F.; Galbraith, S.E.; Radford, A.D.; Dove, W.; Takasaki, T.; Kurane, I.; Solomon, T. Molecular phylogenetic and evolutionary analysis of Muar strain of Japanese encephalitis virus reveal it is the missing fifth genotype. Infect. Genet. Evol. 2011, 11, 855–862. [Google Scholar]
- Cook, S.; Holmes, E.C. A multigene analysis of the phylogenetic relationships among the flaviruses (Family Flaviviridae) and the evaluation of vector transmission. Arch. Virol. 2005, 151, 309–325. [Google Scholar]
- Kuno, G.; Chang, G.-J.J.; Tsuchiya, R.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the Genus. Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar]
- Kuno, G.; Chang, G.-J.J.; Chien, L.-J. Correlations of phylogenetic relation with host range, length of ORF or genes, and viral classification among the members of the Genus Flavivirus. In Viral Genes: Diversity, Properties and Parameters; Feng, Z., Long, M., Eds.; Nova Biomedical Books: New York, NY, USA, 2009; pp. 1–33. [Google Scholar]
- Kuno, G. The boundaries of arboviruses: Complexities revealed in their host ranges, virus-host interactions and evolutionary relationships. In Arboviruses: Molecular Biology, Evolution and Control, Vasilakis, N., Gubler, D.J., Eds.; Caister Academic Press: Norfolk, UK, 2016; pp. 219–268. [Google Scholar]
- Hale, J.E.; Lee, L.H. A serological investigation of six encephalitis viruses isolated in Malaysia. Brit. J. Exp. Pathol. 1954, 35, 426–433. [Google Scholar]
- Hasegawa, H.; Yoshida, M.; Fujita, S.; Kobayashi, Y. Comparison of structural proteins among antigenically different Japanese encephalitis virus strains. Vaccine 1994, 12, 841–844. [Google Scholar]
- Zanotto, P.M.; Gao, G.F.; Gritsun, T.; Marin, M.S.; Jiang, W.R.; Vanugopal, K.; Reid, H.W.; Gould, E.A. An arbovirus cline across the northern hemisphere. Virology 1995, 210, 152–159. [Google Scholar]
- Heinze, D.M.; Gould, E.A.; Forrester, N.L. Revisiting the clinal concept of evolution and dispersal for the tick-borne flaviviruses by using phylogeographic and biogeographic analyses. J. Virol. 2012, 86, 8663–8671. [Google Scholar]
- Gaunt, M.W.; Pettersson, J.H.-O.; Kuno, G.; Gaunt, B.; de Lamballerie, X.; Gould, E.A. Widespread interspecific phylogenetic tree incongruence between mosquito-borne and insect-specific flaviviruses at hotspots originally identified in Zika virus. Virus Evol. 2022, 8, 1–17. [Google Scholar]
- Calisher, C.H. Antigenic classification and taxonomy of flaviviruses (Family Flaviviridae) emphasizing a universal system for the taxonomy of viruses causing tick-borne encephalitis. Acta Virol. 1988, 32, 469–478. [Google Scholar]
- Kitchen, A.; Shackleton, L.A.; Holmes, E.C. Family level phylogenies reveal modes of macroevolution in RNA viruses. Proc. Nat. Acad. Sci. USA 2011, 108, 238–243. [Google Scholar]
- Babayan, S.A.; Orton, R.J.; Streicker, D.G. Predicting reservoir hosts and arthropod vectors from evolutionary significances in RNA virus genomes. Science 2018, 362, 577–580. [Google Scholar]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic flaviviruses. Nat. Comm. 2018, 9, 5425. [Google Scholar]
- Thompson, C.W.; Phelps, K.L.; Allard, M.W.; Cook, J.A.; Dunnum, J.L.; Ferguson, A.W.; Gelang, M.; Khan, F.A.A.; Paul, D.L.; Reeder, D.M.; et al. Preserve a voucher specimen! The critical need for integrating natural history collections in infectious disease studies. mBio 2021, 12, e0269820. [Google Scholar]
- Kuno, G.; Mackenzie, J.S.; Junglen, S.; Hubálek, Z.; Plyusnin, A.; Gubler, D.J. Vertebrate reservoirs of arboviruses: Myth, synonym of amplifier, or reality? Viruses 2017, 9, 185. [Google Scholar]
- Gaunt, M.W.; Sall, A.A.; de Lamballerie, X.; Falconar, A.K.I.; Dzhivanian, T.I.; Gould, E.A. Phylogentic relationships of flaviviruses correlate with their epidemiology, disease association and biogeography. J. Gen. Virol. 2001, 82, 1867–1876. [Google Scholar]
- Reisen, W.K.; Borham, P.F.L. Host selection patterns of some Pakistan mosquitoes. Am. J. Trop. Med. Hyg. 1979, 28, 408–421. [Google Scholar]
- Reuben, R.; Thenomozhi, V.; Samuel, P.P.; Gajanana, A.; Mani, T.R. Mosqjuito blood feeding patterns as a factor in the epidemiology of Japanese encephalitis in Southern India. Am. J. Trop. Med. Hyg. 1992, 46, 654–663. [Google Scholar]
- Mwadawiro, C.; Boots, M.; Tuno, N.; Suwungkerd, W.; Tuda, Y. Heterogeneity in the host preference of Japanese encephalitis vectors in Chiang Mai, northern Thailand. Tr. R. Soc. Trop. Med. Hyg. 2000, 94, 238–242. [Google Scholar]
- Aubert, W.F. Possible role of ticks as reservoir of rabies virus (in French). Comp. Immunol. Microbiol. Infect. Dis. 1982, 5, 343–349. [Google Scholar]
- Traavik, T.; Mehl, R.; Kjeldsberg, E. “Runde” virus, a coronavirus-like agent associated with seabirds and ticks. Arch. Virol. 1977, 55, 25–38. [Google Scholar]
- Slayer, K.A.; Barbet, A.F.; Chamberlain, C.; Clapp, W.L.; Aleman, R.; Loeb, J.C.; Lednicky, J.A. Isolation of Tacaribe virus, a Caribbean arenavirus, from host-seeking Amblyomma americanum ticks in Florida. PLoS ONE 2014, 9, e115769. [Google Scholar]
- Hajnická, V.; Kúdelová, M.; Štibrániová, I.; Slovák, M.; Bartiková, P.; Halásová, Z.; Pancik, P.; Belvoncikova, P.; Vibora, M.; Holikova, V. Tick-borne transmission of murine gammaherpes virus 68. Front. Cell. Infect. Microbiol. 2017, 7, 458. [Google Scholar]
- Kobayashi, D.; Kuwata, R.; Kimura, T.; Faizah, A.N.; Azerigyik, F.A.; Higa, Y.; Hayashi, T.; Sawabe, K.; Isawa, H. Detection of Japanese encephalitis virus RNA in host-questing ticks in Japan, 2019–2020. Am. J. Trop. Med. Hyg. 2022, 106, 1725–1728. [Google Scholar]
- Coggershall, L.T. The transmission of lymphocytic choriomeningitis by mosquitoes. Science 1939, 89, 515–516. [Google Scholar]
- van den Bosch, C.A. Is endemic Burkitt’s lymphoma an alliance between three infections and a tumour promoter? Lancet Oncol. 2004, 5, 738–746. [Google Scholar]
- Zhang, Y.; Liu, D.; Feng, D.; Tang, R.; Li, Y.; You, X. An animal study on transmission of hepatitis B virus through mosquitoes. Clin. Med. J. 1995, 108, 895–897. [Google Scholar]
- Humphrey-Smith, L.; Donker, G.; Turzo, A.; Chastel, C.; Schmidt-Mayerova, H. Evaluation of mechanical transmission of HIV by African soft tick, Ornithodoros moubata. AIDS 1993, 7, 341–347. [Google Scholar]
- Buck, C.; Pauline, G.P.; Medina, D.J.; Hsiung, G.D.; Campbell, T.W.; Walsh, M.T. Isolation of St. Louis encephalitis virus from a killer whale. Clin. Diagn. Virol. 1993, 1, 109–112. [Google Scholar]
- Miller, D.L.; Mauel, M.J.; Baldwin, C.; Batle, G.; Ingram, D.; Hines, M.E.; Frazier, K. West Nile virus in farmed alligators. Emerg. Infect. Dis. 2003, 9, 794–799. [Google Scholar]
- Li, X.-D.; Qiao, M.-M.; Deng, X.-Y.; Chen, X.; Sung, S.-Y.; Zhang, Q.; Zhang, W.J.; Tan, F.-F.; Sun, Z.; Chen, Z.-H.; et al. Lethal encephalitis in seals with Japanese encephalitis virus infection, China, 2017. Emerg. Infect. Dis. 2019, 25, 1539–1542. [Google Scholar]
- Hoffmann, H.-H.; Schneider, W.M.; Rozen-Gagnon, K.; Milo, L.A.; Schuster, F.; Razook, B.; Jacobson, E.; Wu, X.-F.; Yi, S.; Rudin, C.M.; et al. TMEM41B is a pan-flavivirus host factor. Cell 2021, 184, 133–148. [Google Scholar]
- Hameed, M.; Wahaab, A.; Nawaz, M.; Khan, S.; Nazir, J.; Liu, K.; Wei, J.-C.; Ma, Z.-Y. Potential role of birds in Japanese encephalitis virus zoonotic transmission and genotype shift. Viruses 2021, 13, 357. [Google Scholar]
- Tiawsirisup, S.; Nuchprayoon, S. Mosquito distribution and Japanese encephalitis virus infection in the immigration bird (Asian open-billed stork) nested area in Puthum Thani Province, Central Thailand. Parasitol. Res. 2010, 106, 907–910. [Google Scholar]
- Changbunjong, T.; Weluwanarak, T.; Taowan, N.; Suksai, P.; Chamsai, T.; Sedwisai, P. Seasonal abundance and potential of Japanese encephalitis virus infection in mosquitoes at the nesting colony of ardeid birds, Thailand. Asian J. Trop. Biomed. 2013, 3, 207–210. [Google Scholar]
- Park, S.L.; Huang, Y.-J.H.; Vanlandingham, D.L. Re-examining the importance of pigs in the transmission of Japanese encephalitis virus. Pathogens 2022, 22, 575. [Google Scholar]
- Miyake, M.; Takaki, F. Morphologische Studien über die experimentelle epidemische Encephalitis (Sommer encephalitis) bei Affen I. Mitteilung: Über die anatomisch-histologischen Verunderungen bei nasal angesteckten Affen, insbesondere der inneren Organe. J. Jpn. Assoc. Pathol. 1940, 30, 110–117. [Google Scholar]
- Schmaljohn, C.; Blair, C.D. Persistent infection of cultured mammalian cells by Japanese encephalitis virus. J. Virol. 1977, 24, 580–589. [Google Scholar]
- Takegami, T.; Hotta, S. Synthesis and localization of Japanese encephalitis virus RNAs in the infected cells. Microbiol. Immunol. 1990, 34, 849–857. [Google Scholar]
- Lin, K.-C.; Chang, H.-L.; Chang, R.-Y. Accumulation of a 3’-terminal genome fragment in Japanese encephalitis virus-infected mammalian and mosquito cells. J. Virol. 2004, 78, 5133–5138. [Google Scholar]
- Griffin, D.E. Why does viral RNA sometimes persist after recovery from acute infections? PLoS Biol. 2002, 20, e3001687. [Google Scholar]
- Yamanishi, S.; Fujii, Y.; Kameyama, T.; Miki, K. Detection of Japanese encephalitis virus genomes in the blood of pigs in winter. Jpn. Soc. Vet. Med. 1995, 18, 803–808. (In Japanese) [Google Scholar]
- Chapagain, S.; Singh, P.P.; Le, K.; Safronetz, D.; Wood, H.; Karniychuk, U. Japanese encephalitis virus persists in the human reproductive epithelium and porcine reproductive tissues. PLoS Negl. Trop. Dis. 2022, 16, e0010656. [Google Scholar]
- Habu, A.; Murakami, Y.; Ogasa, A.; Fujisaki, Y. Dysfunction of spermatogenesis by Japanese encephalitis viral infection of swine and viral shedding into semen. Uirusu 1977, 27, 21–26. (In Japanese) [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuno, G. Contrasting the Practices of Virus Isolation and Characterization between the Early Period in History and Modern Times: The Case of Japanese Encephalitis Virus. Viruses 2022, 14, 2640. https://doi.org/10.3390/v14122640
Kuno G. Contrasting the Practices of Virus Isolation and Characterization between the Early Period in History and Modern Times: The Case of Japanese Encephalitis Virus. Viruses. 2022; 14(12):2640. https://doi.org/10.3390/v14122640
Chicago/Turabian StyleKuno, Goro. 2022. "Contrasting the Practices of Virus Isolation and Characterization between the Early Period in History and Modern Times: The Case of Japanese Encephalitis Virus" Viruses 14, no. 12: 2640. https://doi.org/10.3390/v14122640