

The Molecular Characterization of a New Prunus-Infecting Cheravirus and Complete Genome Sequence of Stocky Prune Virus
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Double-Stranded RNA Extraction, Library Preparation and High-Throughput Sequencing (HTS)
2.3. Total RNA (totRNA) Extraction and HTS
2.4. Total Nucleic Acids (TNAs) Extraction and RT-PCR Detection of the Novel Cheravirus
2.5. HTS Data Analyses
2.6. Genome Sequence Completion for Isolates of the Novel Cheravirus and StPV
2.7. Pairwise Comparisons, Phylogenetic and Recombination Analyses
2.8. Biological Indexing in Greenhouse
3. Results
3.1. Determination of the Complete Genome Sequence of a Newly Discovered Cheravirus and of StPV
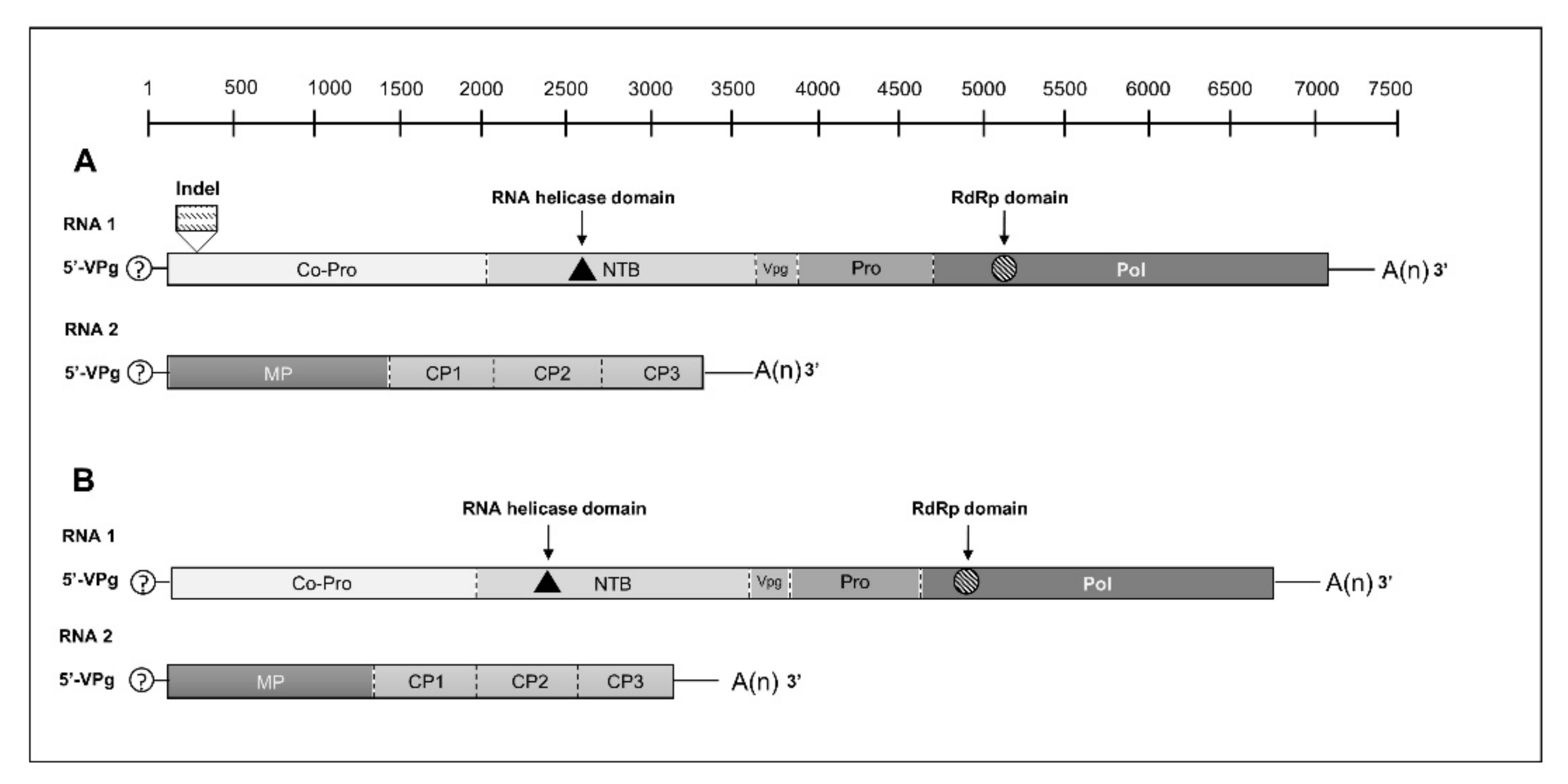
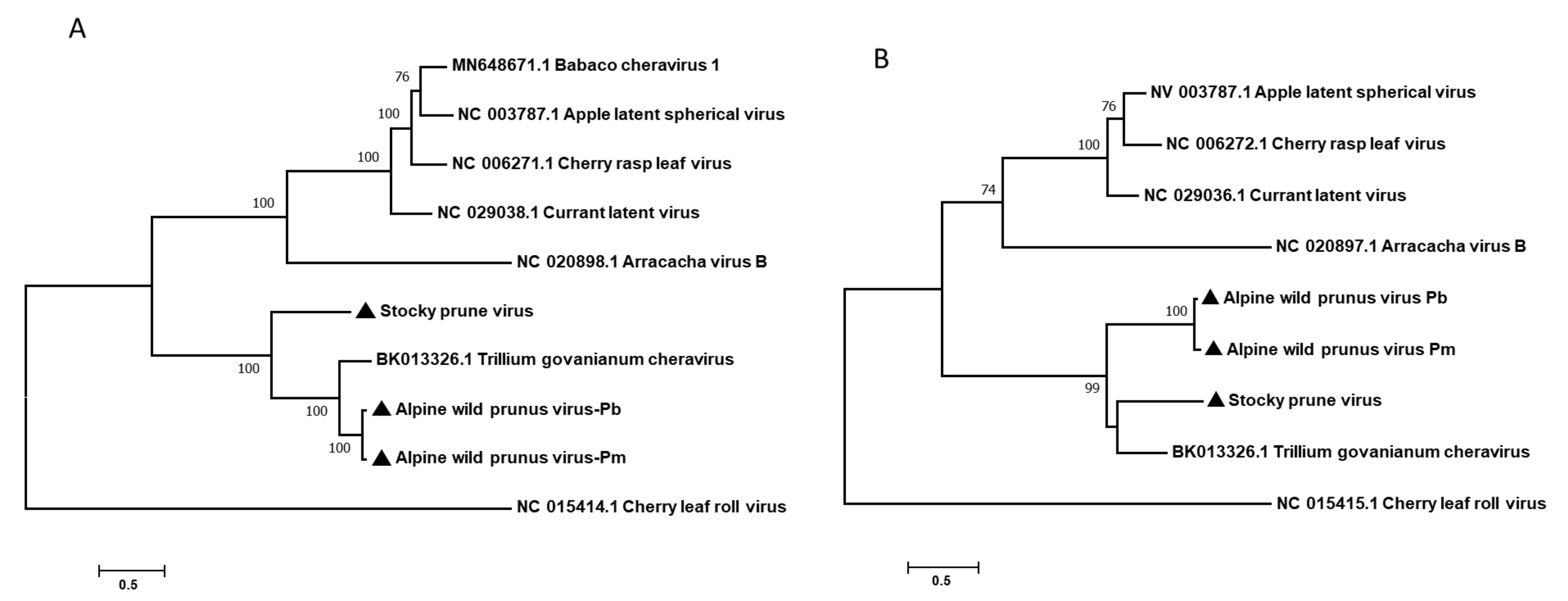
3.2. Genomic Organization of AWPV and StPV and Determination of Their Phylogenetic Relationships
3.3. Biological Indexing of AWPV-Infected Apricot Accession
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, I.P.; Glover, R.; Souza-Richards, R.; Bennett, S.; Hany, U.; Boonham, N. Complete genome sequence of arracacha virus B: A novel cheravirus. Arch. Virol. 2013, 158, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.R.; Perry, K.L.; De Jong, W. A new potato virus in a new lineage of picorna-like viruses. Arch. Virol. 2004, 149, 2141–2154. [Google Scholar] [CrossRef] [PubMed]
- Candresse, T.; Delbos, R.; Le Gall, O.; Dunez, J.; Desvignes, J. Characterization of stocky prune virus, a new nepovirus detected in french prunes. Acta Hortic. 1998, 472, 175–182. [Google Scholar] [CrossRef]
- Candresse, T.; Svanella-Dumas, L.; Le Gall, O. Characterization and partial genome sequence of stocky prune virus, a new member of the genus Cheravirus. Arch. Virol. 2006, 151, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Cornejo-Franco, J.F.; Medina-Salguero, A.; Flores, F.; Chica, E.; Grinstead, S.; Mollov, D.; Quito-Avila, D.F. Exploring the virome of Vasconcellea x heilbornii: The first step towards a sustainable production program for babaco in Ecuador. European J. Plant Pathol. 2020, 157, 961–968. [Google Scholar] [CrossRef]
- James, D.; Upton, C. Nucleotide sequence analysis of RNA-2 of a flat apple isolate of Cherry rasp leaf virus with regions showing greater identity to animal picornaviruses than to related plant viruses. Arch. Virol. 2002, 147, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yoshikawa, N.; Takahashi, T.; Ito, T.; Yoshida, K.; Koganezawa, H. Nucleotide sequence and genome organization of Apple latent spherical virus: A new virus classified into the family Comoviridae. J. Gen. Virol. 2000, 81, 541–547. [Google Scholar] [CrossRef]
- Petrzik, K.; Přibylová, J.; Špak, J.; Havelka, J. Partial genome sequence of currant latent virus, a new chera-like virus related to Apple latent spherical virus. J. Gen. Plant Pathol. 2015, 81, 142–145. [Google Scholar] [CrossRef]
- Sidharthan, V.K.; Kalaivanan, N.S.; Baranwal, V.K. Discovery of putative novel viruses in the transcriptomes of endangered plant species native to India and China. Gene 2021, 786, 145626. [Google Scholar] [CrossRef]
- Hansen, A.J.; Nyland, G.; McElroy, F.D.; Stace-Smith, R. Cherry rasp leaf disease in North America. Phytopathology 1974, 64, 721–727. [Google Scholar] [CrossRef]
- Jones, R.A.C. Tests for transmission of four potato viruses through potato true seed. Ann. Appl. Biol. 1982, 100, 315–320. [Google Scholar] [CrossRef]
- Nakamura, K.; Yamagishi, N.; Isogai, M.; Komori, S.; Ito, T.; Yoshikawa, N. Seed and pollen transmission of Apple latent spherical virus in apple. J. Gen. Plant Pathol. 2011, 77, 48–53. [Google Scholar] [CrossRef]
- Nyland, G.; Lownsbery, B.F.; Lowe, S.K.; Mitchell, J.F. The transmission of cherry rasp leaf virus by Xiphinema americanum. Phytopathology 1969, 59, 1111–1112. [Google Scholar]
- Stace-Smith, R.; Hansen, A.J. Some properties of cherry rasp leaf virus. Acta Hort. 1976, 67, 193–198. [Google Scholar] [CrossRef]
- Wagnon, H.K. Investigations of cherry rasp leaf disease in California. Plant Dis. Rep. 1968, 52, 618–622. [Google Scholar]
- Desvignes, J.C. Stocky prune. In Les Virus des Arbres Fruitiers; Desvignes, J.C., Ed.; CTIFL: Paris, France, 1990; pp. 51–52. [Google Scholar]
- Liu, S.; Decroocq, S.; Harte, E.; Tricon, D.; Chague, A.; Balakisiyeva, G.; Kostriesyna, T.; Turdiev, T.; Fisher-Le Saux, M.; Dallot, S.; et al. Genetic diversity and population structure analyses in the Alpine plum (Prunus brigantina Vill.) confirm its affiliation to the Armeniaca section. TGG 2021, 17, 2. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Bergey, B.; Candresse, T. Viral Double-Stranded RNAs (dsRNAs) from Plants: Alternative Nucleic Acid Substrates for High-Throughput Sequencing. Methods Mol Biol. 2018, 1746, 45–53. [Google Scholar] [CrossRef]
- François, S.; Filloux, D.; Fernandez, E.; Ogliastro, M.; Roumagnac, P. Viral Metagenomics Approaches for High-Resolution Screening of Multiplexed Arthropod and Plant Viral Communities. Methods Mol Biol. 2018, 1746, 77–95. [Google Scholar] [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Foissac, X.; Svanella-Dumas, L.; Gentit, P.; Dulucq, M.J.; Marais, A.; Candresse, T. Polyvalent degenerate oligonucleotides reverse transcription-polymerase chain reaction: A polyvalent detection and characterization tool for trichoviruses, capilloviruses, and foveaviruses. Phytopathology 2005, 95, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Marais, A.; Faure, C.; Couture, C.; Bergey, B.; Gentit, P.; Candresse, T. Characterization by deep sequencing of divergent Plum bark necrosis stem pitting-associated virus (PBNSPaV) isolates and development of a broad-spectrum PBNSPaV detection assay. Phytopathology 2014, 104, 660–666. [Google Scholar] [CrossRef] [Green Version]
- Youssef, F.; Marais, A.; Faure, C.; Barone, M.; Gentit, P.; Candresse, T. Characterization of Prunus-infecting Apricot latent virus-like Foveaviruses: Evolutionary and taxonomic implications. Virus Res. 2011, 155, 440–445. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Gentit, P.; Foissac, X.; Svanella-Dumas, L.; Peypelut, M.; Macquaire, G.; Candresse, T. Biological properties and partial characterisation of two different Foveavirus associated with similar disorders of cherry trees. Acta Hortic 2001, 550, 161–168. [Google Scholar] [CrossRef]
- Thompson, J.R.; Dasgupta, I.; Fuchs, M.; Iwanami, T.; Karasev, A.V.; Petrzik, K.; Sanfaçon, H.; Tzanetakis, I.; van der Vlugt, R.; Wetzel, T.; et al. ICTV Virus Taxonomy Profile: Secoviridae. J. Gen. Virol. 2017, 98, 529–531. [Google Scholar] [CrossRef]
- Kozak, M. Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 1986, 44, 283–292. [Google Scholar] [CrossRef]
- Khalili, M.; Candresse, T.; Koloniuk, I.; Safarova, D.; Brans, Y.; Faure, C.; Delmas, M.; Massart, S.; Aranda, M.A.; Cagalayan, K.; et al. The expanding menagerie of Prunus-infecting luteoviruses. Phytopathology, 2022; Online ahead of print. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Virus-Isolate 1 | Method | Trimmed Reads | Segment | Average Coverage | Mapped Reads (%) | Accession Numbers |
|---|---|---|---|---|---|---|
| AWPV-Pb | dsRNA 2 | 1,893,230 | RNA1 | 8886x | 30.5% | OP328247 |
| RNA2 | 15047x | 22% | OP328248 | |||
| AWPV-Pm | RNA | 61,886,192 | RNA1 | 3503x | 0.2% | OP328249 |
| RNA2 | 10145x | 0.39% | OP328250 | |||
| StPV | RNA | 57,041,248 | RNA1 | 4051x | 0.34% | OP328251 |
| RNA2 | 4476x | 0.18% | OP328252 |
| Virus 1 | Amino Acid Identity (%) | |
|---|---|---|
| Pro-Pol 2 | Capsid Protein 3 | |
| AWPV-Pm | 97.1% | 93.9% |
| StPV | 64.7% | 33.7% |
| ALSV | 38.7% | 16.3% |
| AVB | 41.6% | 12.2% |
| CRLV | 38.9% | 15.9% |
| CuLV | 39.4% | 14.6% |
| TgCV | 86.3% | 41.1% |
| BabChV-1 | 39.2% | na 4 |
| Virus | Segment | Length (nt) | Polyprotein (aa) | 5′ NCR 1 (nt) | 3′ NCR (nt) |
|---|---|---|---|---|---|
| AWPV-Pb | RNA1 | 7354 | 2358 | 50 | 230 |
| RNA2 | 3574 | 1088 | 69 | 240 | |
| AWPV-Pm | RNA1 | 7491 | 2404 | 50 | 229 |
| RNA2 | 3568 | 1086 | 70 | 241 | |
| StPV | RNA1 | 7021 | 2230 | 80 | 251 |
| RNA2 | 3495 | 1056 | 78 | 249 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalili, M.; Candresse, T.; Brans, Y.; Faure, C.; Audergon, J.-M.; Decroocq, V.; Roch, G.; Marais, A. The Molecular Characterization of a New Prunus-Infecting Cheravirus and Complete Genome Sequence of Stocky Prune Virus. Viruses 2022, 14, 2325. https://doi.org/10.3390/v14112325
Khalili M, Candresse T, Brans Y, Faure C, Audergon J-M, Decroocq V, Roch G, Marais A. The Molecular Characterization of a New Prunus-Infecting Cheravirus and Complete Genome Sequence of Stocky Prune Virus. Viruses. 2022; 14(11):2325. https://doi.org/10.3390/v14112325
Chicago/Turabian StyleKhalili, Maryam, Thierry Candresse, Yoann Brans, Chantal Faure, Jean-Marc Audergon, Véronique Decroocq, Guillaume Roch, and Armelle Marais. 2022. "The Molecular Characterization of a New Prunus-Infecting Cheravirus and Complete Genome Sequence of Stocky Prune Virus" Viruses 14, no. 11: 2325. https://doi.org/10.3390/v14112325