
Host-Level Susceptibility and IRF1 Expression Influence the Ability of IFN-γ to Inhibit KSHV Infection in B Lymphocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Lines
2.2. Purification of Cell-Free KSHV Virions
2.3. Isolation of Primary Lymphocytes from Human Tonsils
2.4. Infection of Primary Lymphocytes with KSHV
2.5. Flow Cytometry Analysis
2.6. Statistical Analysis
3. Results
3.1. IFN-γ Inhibits KSHV Infection in Tonsil-Derived Lymphocytes That Are Highly Susceptible to Infection
3.2. IFN-γ Decreases the Frequency of Infected Plasma Cells in Responsive Samples
3.3. IFN-γ Increases Germinal Center B Cell Frequencies in Responsive Samples
3.4. Response to IFN-γ Stimulation Is Associated with Samples That Have Increased Frequency of Immature B Cells and High Baseline IRF1 Expression
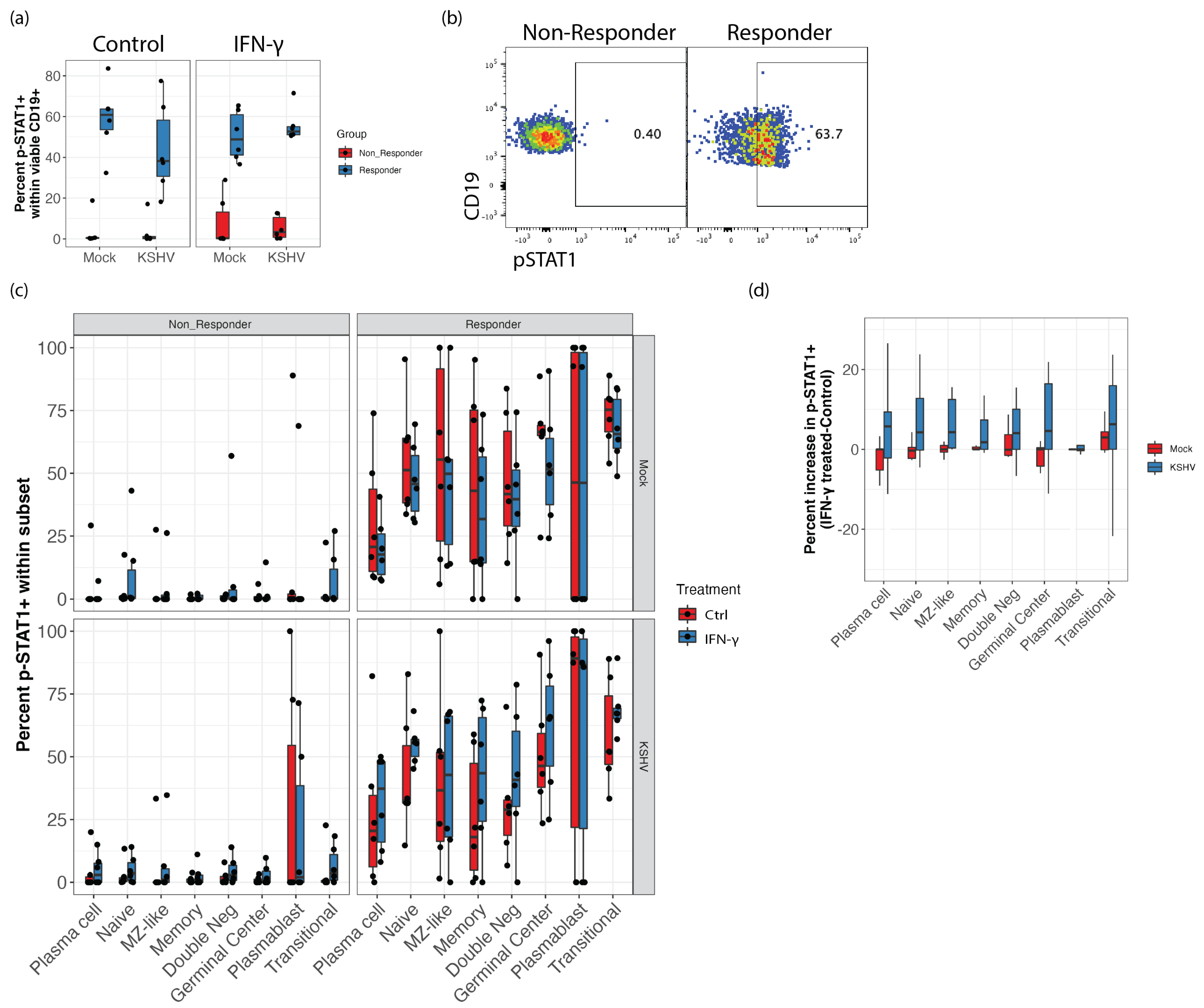
3.5. STAT1 Phosphorylation Is Elevated in IFN-γ Responsive Samples and KSHV Infection Increases STAT1 Phosphorylation Response to IFN-γ Treatment in GCB
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ethel, C.; Yuan, C.; Patrick, S.M.; Jonathan, W.S.; Daniel, M.K. Kaposi’s sarcoma–associated herpesvirus-like DNA sequences in AIDS-related body-cavity–based lymphomas. N. Engl. J. Med. 1995, 332, 1186–1191. [Google Scholar] [CrossRef]
- Soulier, J.; Grollet, L.; Oksenhendler, E.; Cacoub, P.; Cazals-Hatem, D.; Babinet, P.; D’Agay, M.; Clauvel, J.; Raphael, M.; Degos, L.; et al. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease [see comments]. Blood 1995, 86, 1276–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uldrick, T.S.; Wang, V.; O’Mahony, D.; Aleman, K.; Wyvill, K.M.; Marshall, V.; Steinberg, S.M.; Pittaluga, S.; Maric, I.; Whitby, D.; et al. An Interleukin-6–Related Systemic Inflammatory Syndrome in Patients Co-Infected with Kaposi Sarcoma–Associated Herpesvirus and HIV but without Multicentric Castleman Disease. Clin. Infect. Dis. 2010, 51, 350–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, C.; Barnard, S.; Blackbourn, D.J.; Davison, A.J. Transcription mapping of human herpesvirus 8 genes encoding viral interferon regulatory factors. J. Gen. Virol. 2003, 84, 1471–1483. [Google Scholar] [CrossRef]
- Seungmin, H.; Nicole, S.M.; Monique, W.B.; Gautam, G.; Erning, D.; Lei, Z.; Bimmi, S.; Michael, S.D.; Adish, D.; Stanislav, V.S.; et al. Nondegradative role of Atg5-Atg12/Atg16L1 autophagy protein complex in antiviral activity of interferon gamma. Cell Host Microbe 2012, 11, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Changotra, H.; Jia, Y.; Moore, T.N.; Liu, G.; Kahan, S.M.; Sosnovtsev, S.V.; Karst, S.M. Type I and Type II Interferons Inhibit the Translation of Murine Norovirus Proteins. J. Virol. 2009, 83, 5683–5692. [Google Scholar] [CrossRef] [Green Version]
- Gajewski, T.F.; Fitch, F.W. Anti-proliferative effect of IFN-gamma in immune regulation. I. IFN-gamma inhibits the proliferation of Th2 but not Th1 murine helper T lymphocyte clones. J. Immunol. 1988, 140, 4245–4252. [Google Scholar]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17–producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar] [CrossRef]
- Leibson, H.J.; Gefter, M.L.; Zlotnik, A.; Marrack, P.; Kappler, J.W. Role of γ-interferon in antibody-producing responses. Nat. 1984, 309, 799–801. [Google Scholar] [CrossRef]
- Mercader, M.; Taddeo, B.; Panella, J.R.; Chandran, B.; Nickoloff, B.J.; Foreman, K.E. Induction of HHV-8 Lytic Cycle Replication by Inflammatory Cytokines Produced by HIV-1-Infected T Cells. Am. J. Pathol. 2000, 156, 1961–1971. [Google Scholar] [CrossRef]
- Park, M.-K.; Cho, H.; Roh, S.W.; Kim, S.-J.; Myoung, J. Cell Type-Specific Interferon-γ-mediated Antagonism of KSHV Lytic Replication. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozharskaya, V.P.; Weakland, L.L.; Offermann, M.K. Inhibition of infectious human herpesvirus 8 production by gamma interferon and alpha interferon in BCBL-1 cells. J. Gen. Virol. 2004, 85, 2779–2787. [Google Scholar] [CrossRef] [PubMed]
- Aalam, F.; Nabiee, R.; Castano, J.R.; Totonchy, J. Analysis of KSHV B lymphocyte lineage tropism in human tonsil reveals efficient infection of CD138+ plasma cells. PLoS Pathog. 2020, 16, e1008968. [Google Scholar] [CrossRef]
- Nedaa, A.; Farizeh, A.; Romina, N.; Jesus, R.C.; Jennifer, T. IL-21 signaling promotes the establishment of KSHV infection in human tonsil lymphocytes by increasing early targeting of plasma cells. bioRxiv 2022. [Google Scholar] [CrossRef]
- Palmerin, N.; Aalam, F.; Nabiee, R.; Muniraju, M.; Ogembo, J.; Totonchy, J. Suppression of DC-SIGN and gH Reveals Complex, Subset-Specific Mechanisms for KSHV Entry in Primary B Lymphocytes. Viruses 2021, 13, 1512. [Google Scholar] [CrossRef]
- Myoung, J.; Ganem, D. Generation of a doxycycline-inducible KSHV producer cell line of endothelial origin: Maintenance of tight latency with efficient reactivation upon induction. J. Virol. Methods 2011, 174, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; p. 213. [Google Scholar]
- Kassambara, A. ggcorrplot: Visualization of a Correlation Matrix using ‘ggplot2’. Available online: https://CRAN.R-project.org/package=ggcorrplot (accessed on 4 August 2022).
- Schloerke, B.; Di, C.; Joseph, L.; Francois, B.; Moritz, M.; Edwin, T.; Amos, E.; Jason, C. GGally: Extension to ‘ggplot2’. 2018. 713. Available online: https://ggobi.github.io/ggally/reference/GGally-package.html (accessed on 4 August 2022).
- Lander, J.P. R for Everyone: Advanced Analytics and Graphics, 2nd ed.; Addison-Wesley Professional: Boston, MA, USA, 2017; p. 528. [Google Scholar]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 4 August 2022).
- Maloney, N.S.; Thackray, L.B.; Goel, G.; Hwang, S.; Duan, E.; Vachharajani, P.; Xavier, R.; Virgin, H.W. Essential Cell-Autonomous Role for Interferon (IFN) Regulatory Factor 1 in IFN-γ-Mediated Inhibition of Norovirus Replication in Macrophages. J. Virol. 2012, 86, 12655–12664. [Google Scholar] [CrossRef] [Green Version]
- Brien, J.D.; Daffis, S.; LaZear, H.M.; Cho, H.; Suthar, M.S.; Gale, M.; Diamond, M.S. Interferon Regulatory Factor-1 (IRF-1) Shapes Both Innate and CD8+ T Cell Immune Responses against West Nile Virus Infection. PLoS Pathog. 2011, 7, e1002230. [Google Scholar] [CrossRef]
- Mboko, W.P.; Rekow, M.M.; Ledwith, M.P.; Lange, P.T.; Schmitz, K.E.; Anderson, S.; Tarakanova, V.L. Interferon Regulatory Factor 1 and Type I Interferon Cooperate To Control Acute Gammaherpesvirus Infection. J. Virol. 2017, 91, e01444-16. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Zhang, Y.-B.; Gui, J.-F.; Lemon, S.M.; Yamane, D. Interferon regulatory factor 1 (IRF1) and anti-pathogen innate immune responses. PLoS Pathog. 2021, 17, e1009220. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Veettil, M.V.; Chandran, B. Kaposi’s Sarcoma Associated Herpesvirus Entry into Target Cells. Front. Microbiol. 2012, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, T.R.R.; Gonçales, J.P.; Júnior, J.V.J.S.; Lorena, V.M.B.D.; Toscano, A.L.C.C.; Akamatsu, S.M.; Salles, A.C.; Tozetto-Mendoza, T.R.; Morais, V.M.S.D.; Coêlho, M.R.C.D. Association of IL-6, IL-10 and CXCL10 serum concentrations with visceral Kaposi’s sarcoma in people living with HIV/AIDS. Hum. Immunol. 2020, 81, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.D.; Aoki, Y.; Chang, Y.; Moore, P.S.; Yarchoan, R.; Tosato, G. Involvement of interleukin-10 (IL-10) and viral IL-6 in the spontaneous growth of Kaposi’s sarcoma herpesvirus-associated infected primary effusion lymphoma cells. Blood 1999, 94, 2871–2879. Available online: https://pubmed.ncbi.nlm.nih.gov/10515891/ (accessed on 4 August 2022). [CrossRef]
- Goodwin, M.M.; Canny, S.; Steed, A.; Virgin, H.W. Murine Gammaherpesvirus 68 Has Evolved Gamma Interferon and Stat1-Repressible Promoters for the Lytic Switch Gene 50. J. Virol. 2010, 84, 3711–3717. [Google Scholar] [CrossRef] [Green Version]
- Reese, T.A.; Wakeman, B.S.; Choi, H.S.; Hufford, M.M.; Huang, S.C.; Zhang, X.; Buck, M.D.; Jezewski, A.; Kambal, A.; Liu, C.Y.; et al. Helminth infection reactivates latent γ-herpesvirus via cytokine competition at a viral promoter. Science 2014, 345, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Steed, A.; Buch, T.; Waisman, A.; Virgin, H.W. Gamma Interferon Blocks Gammaherpesvirus Reactivation from Latency in a Cell Type-Specific Manner. J. Virol. 2007, 81, 6134–6140. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Wang, S.; Sun, F.; Zheng, G.; Wu, T.; Du, Y.; Zhang, S.; Qian, J.; Sun, R. Inhibition of murine herpesvirus-68 replication by IFN-gamma in macrophages is counteracted by the induction of SOCS1 expression. PLoS Pathog. 2018, 14, e1007202. [Google Scholar] [CrossRef]
- Flaño, E.; Kim, I.-J.; Woodland, D.L.; Blackman, M.A. γ-Herpesvirus Latency Is Preferentially Maintained in Splenic Germinal Center and Memory B Cells. J. Exp. Med. 2002, 196, 1363–1372. [Google Scholar] [CrossRef]
- Laichalk, L.L.; Thorley-Lawson, D.A. Terminal Differentiation into Plasma Cells Initiates the Replicative Cycle of Epstein-Barr Virus In Vivo. J. Virol. 2005, 79, 1296–1307. [Google Scholar] [CrossRef] [Green Version]
- Mboko, W.P.; Olteanu, H.; Ray, A.; Xin, G.; Darrah, E.J.; Kumar, S.N.; Kulinski, J.M.; Cui, W.; Dittel, B.N.; Gauld, S.B.; et al. Tumor Suppressor Interferon-Regulatory Factor 1 Counteracts the Germinal Center Reaction Driven by a Cancer-Associated Gammaherpesvirus. J. Virol. 2016, 90, 2818–2829. [Google Scholar] [CrossRef] [PubMed]
- Zenke, K.; Muroi, M.; Tanamoto, K.-I. IRF1 supports DNA binding of STAT1 by promoting its phosphorylation. Immunol. Cell Biol. 2018, 96, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, K.; Rubtsov, A.V.; van Dyk, L.F.; Kappler, J.W.; Marrack, P. T-box transcription factor T-bet, a key player in a unique type of B-cell activation essential for effective viral clearance. Proc. Natl. Acad. Sci. USA 2013, 110, E3216–E3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.L.; Szabo, S.J.; Glimcher, L.H. T-bet regulates IgG class switching and pathogenic autoantibody production. Proc. Natl. Acad. Sci. USA 2002, 99, 5545–5550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancro, M.P. Age-associated B cells. Annu. Rev. Immunol. 2020, 38, 315–340. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B Lymphocytes | |
|---|---|
| Subset | Molecular Markers |
| Plasma | CD19+, CD20+/−, CD138+(Mid to High), CD38− |
| Transitional | CD19+, CD138−, CD38Mid, IgD+ (Mid to High) |
| Plasmablast | CD19+, CD138−, CD38High, IgD+/− (mostly−) |
| Germinal Center | CD19+, CD138−, CD38Mid, IgD− |
| Naïve | CD19+, CD138−, CD38Low, CD27−, IgD+ (Mid to High) |
| Marginal Zone Like (MZ-Like) | CD19+, CD138−, CD38Low, CD27+ (Mid to High), IgD+ (Mid to High) |
| Memory | CD19+, CD138−, CD38Low, CD27+ (Mid to High), IgD− |
| Double Negative | CD19+, CD138−, CD38Low, CD27−, IgD− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alomari, N.; Totonchy, J. Host-Level Susceptibility and IRF1 Expression Influence the Ability of IFN-γ to Inhibit KSHV Infection in B Lymphocytes. Viruses 2022, 14, 2295. https://doi.org/10.3390/v14102295
Alomari N, Totonchy J. Host-Level Susceptibility and IRF1 Expression Influence the Ability of IFN-γ to Inhibit KSHV Infection in B Lymphocytes. Viruses. 2022; 14(10):2295. https://doi.org/10.3390/v14102295
Chicago/Turabian StyleAlomari, Nedaa, and Jennifer Totonchy. 2022. "Host-Level Susceptibility and IRF1 Expression Influence the Ability of IFN-γ to Inhibit KSHV Infection in B Lymphocytes" Viruses 14, no. 10: 2295. https://doi.org/10.3390/v14102295