
Isolation and Characterization of the First Temperate Virus Infecting Psychrobacillus from Marine Sediments

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation
2.2. Isolation and Purification of PVJ1
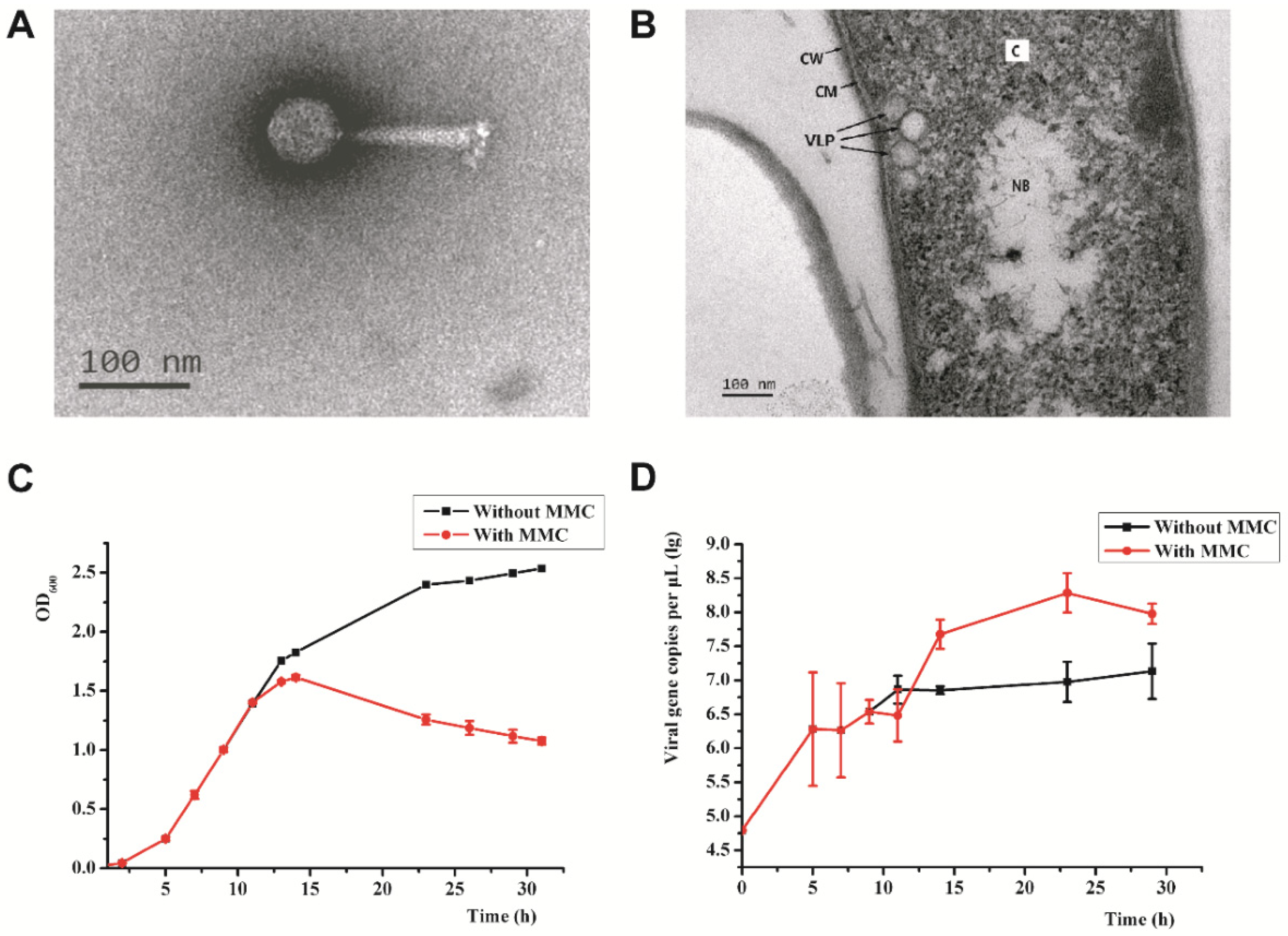
2.3. Transmission Electron Microscopy (TEM)
2.4. DNA Isolation and Genome Analysis
2.5. Quantification of Phage DNA by qPCR
2.6. Host Range Determination
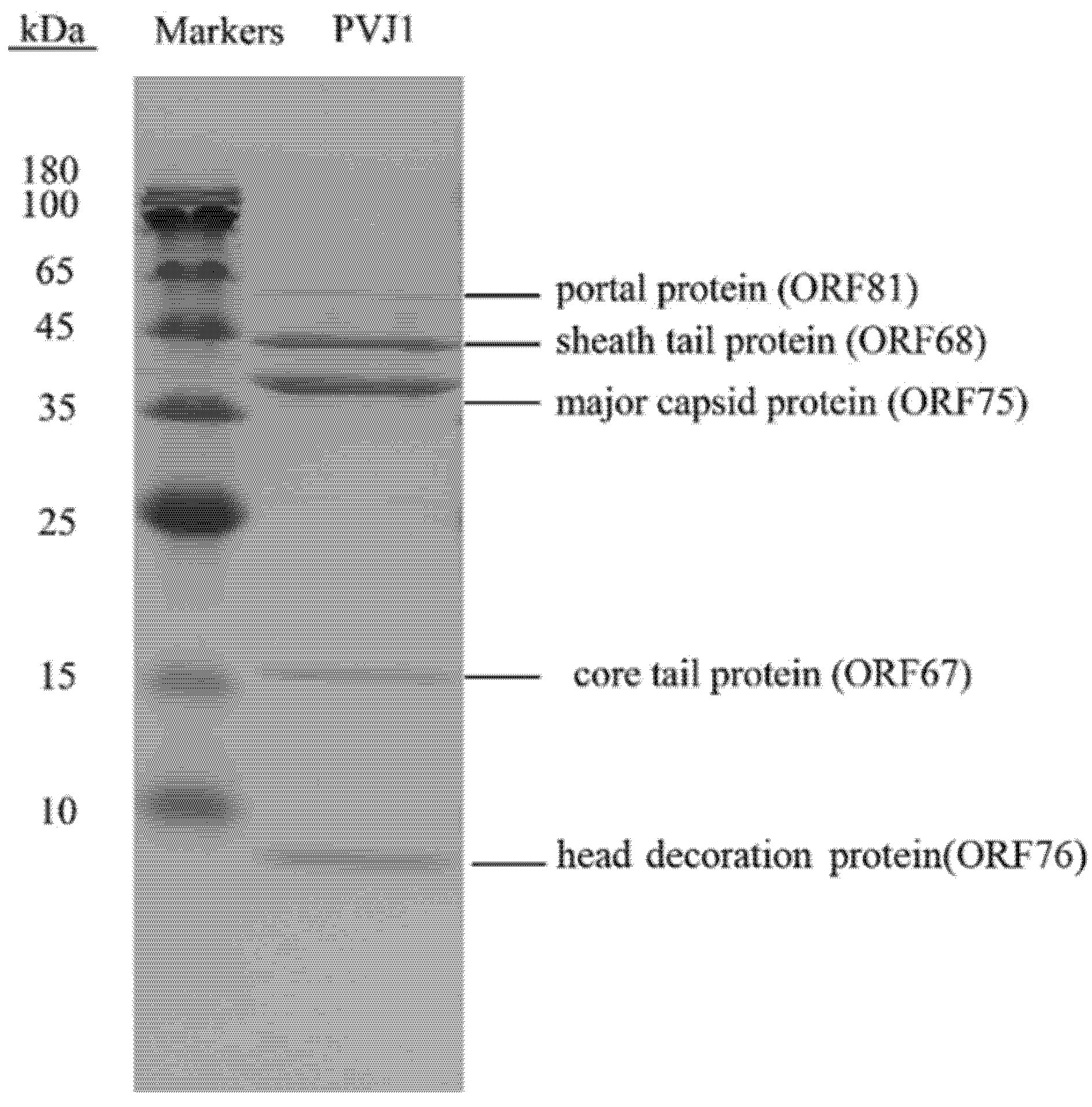
2.7. Protein Analysis
2.8. Atomic Force Microscopy (AFM)
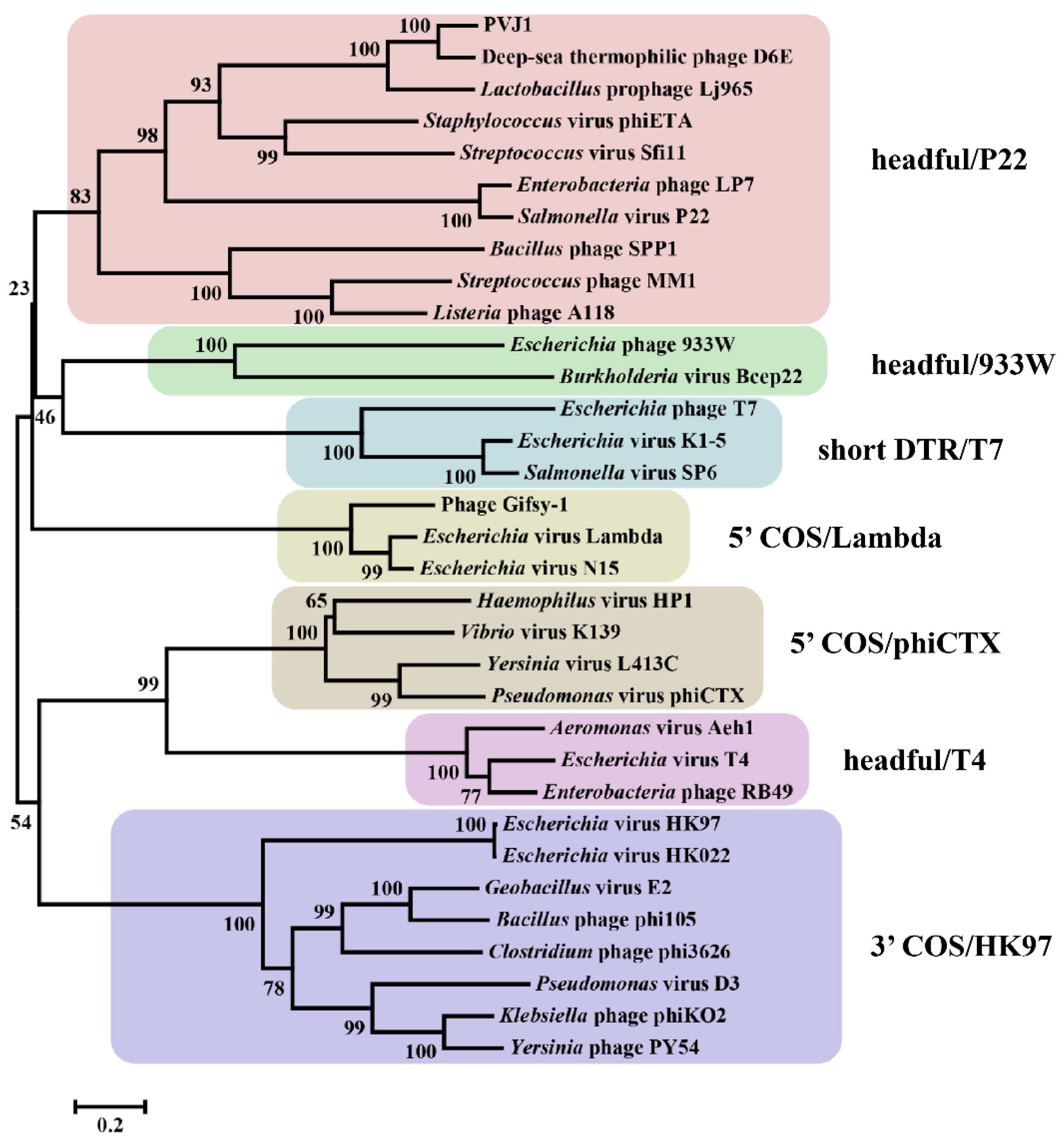
2.9. Phylogenetic Analysis
3. Results and Discussion
3.1. Isolation of Psychrobacillus sp. GC2J1 and Bacteriophage PVJ1
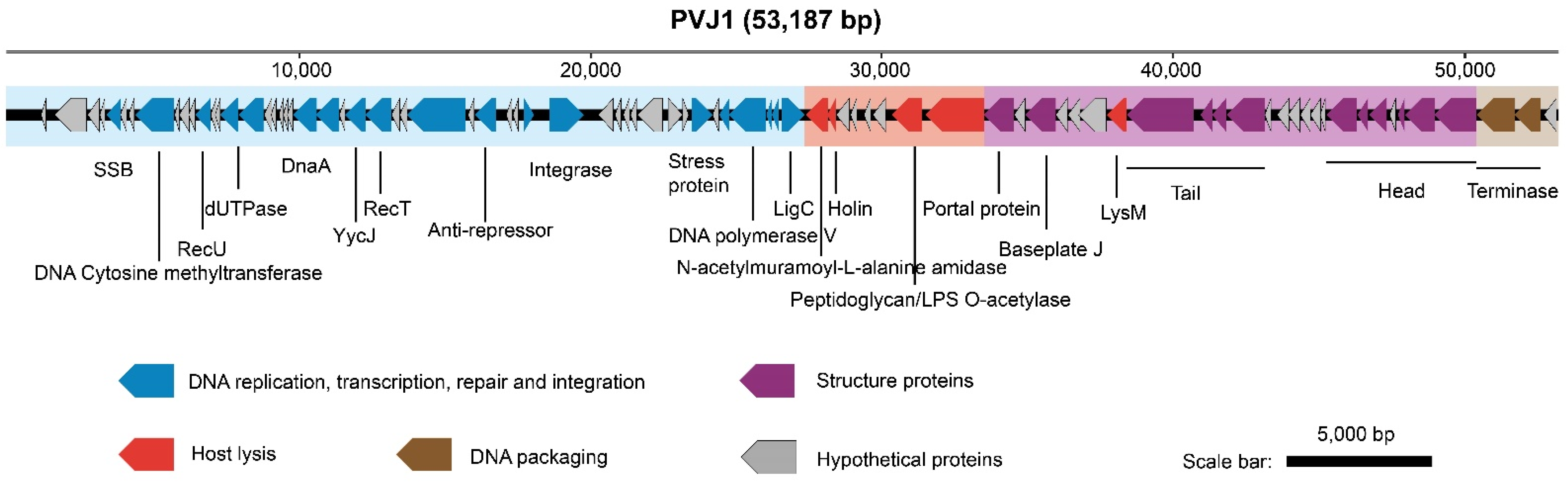
3.2. General Features of the PVJ1 Genome
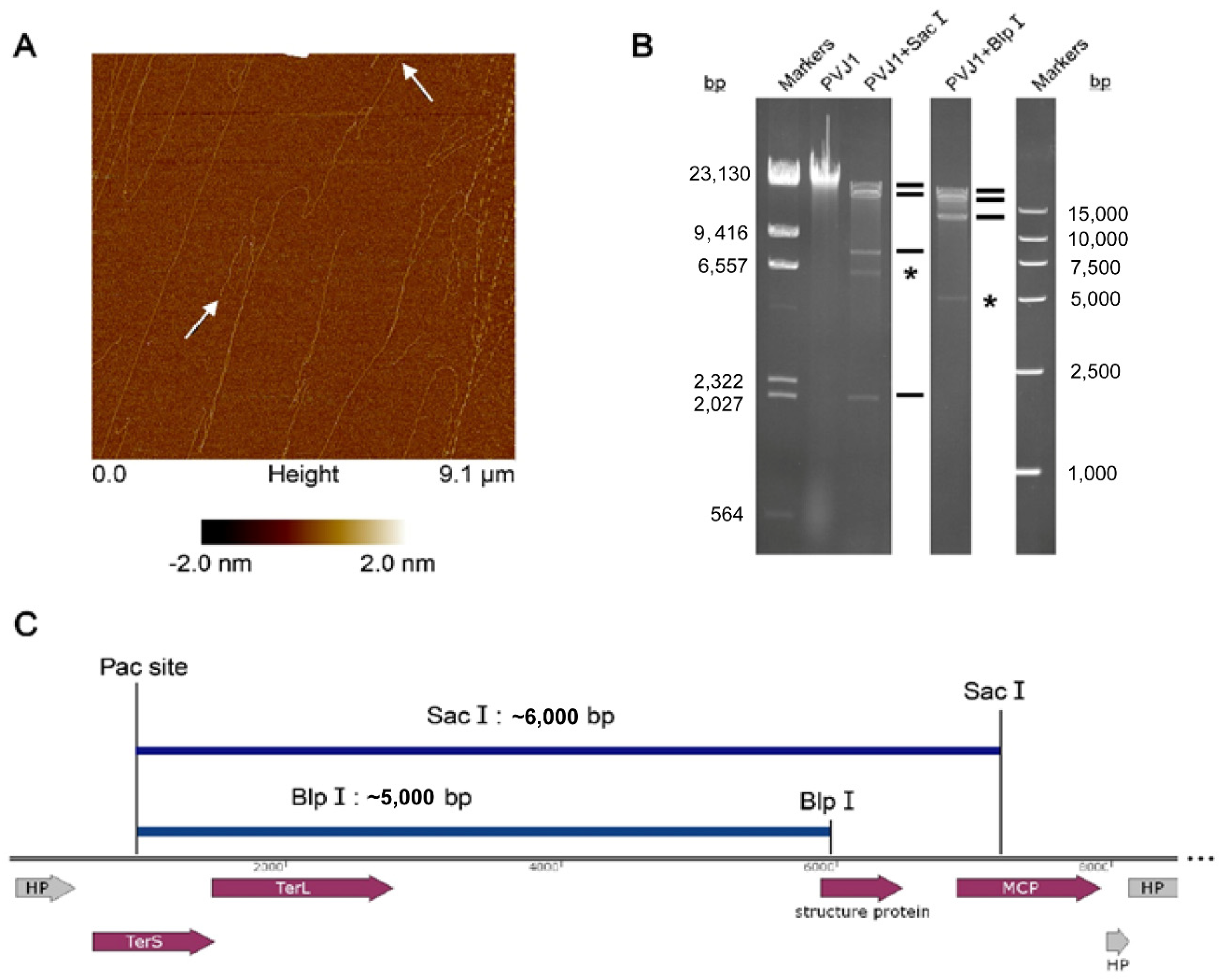
3.3. The PVJ1 Genome Is Linear and Packaged by a Headful Strategy
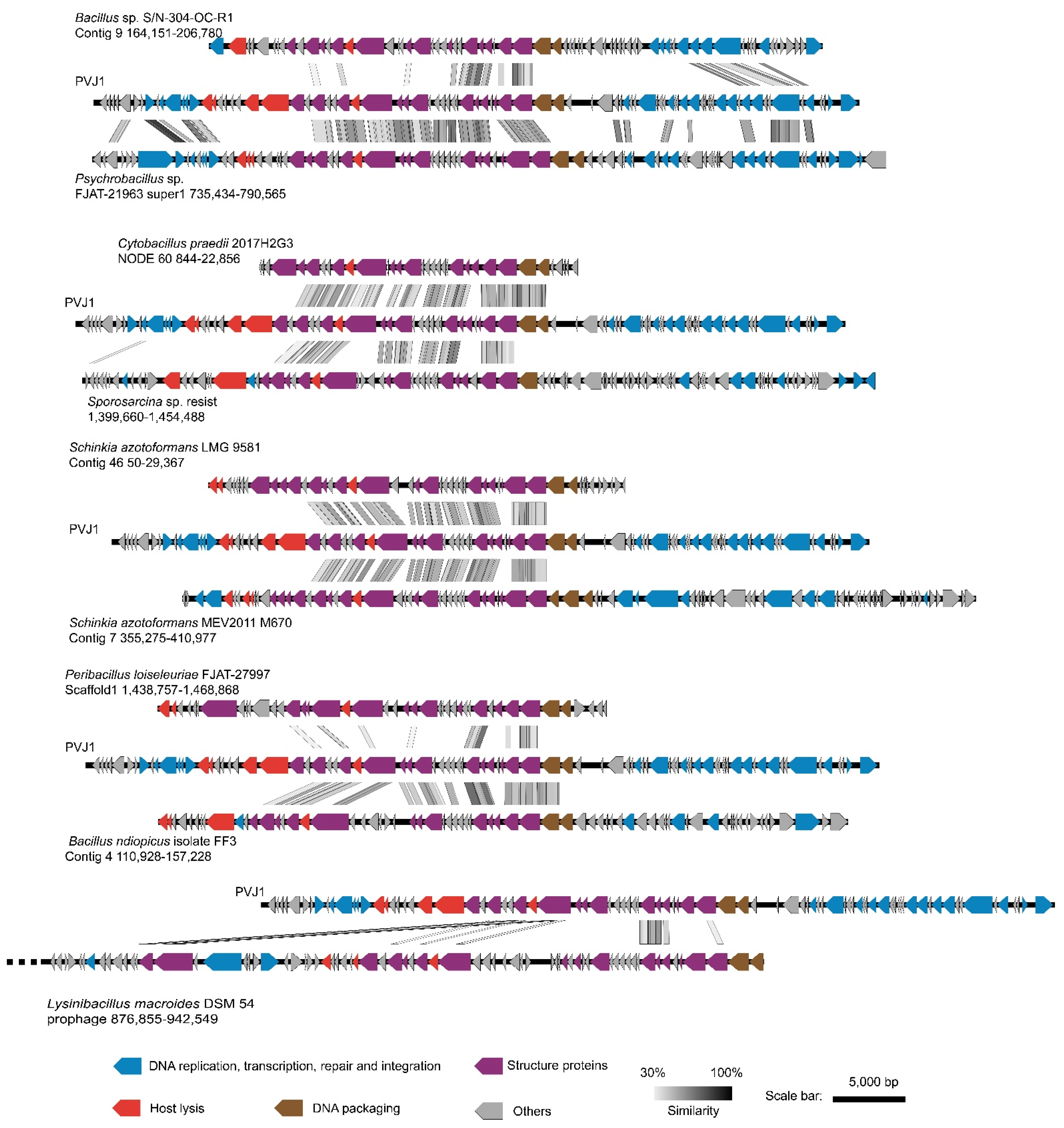
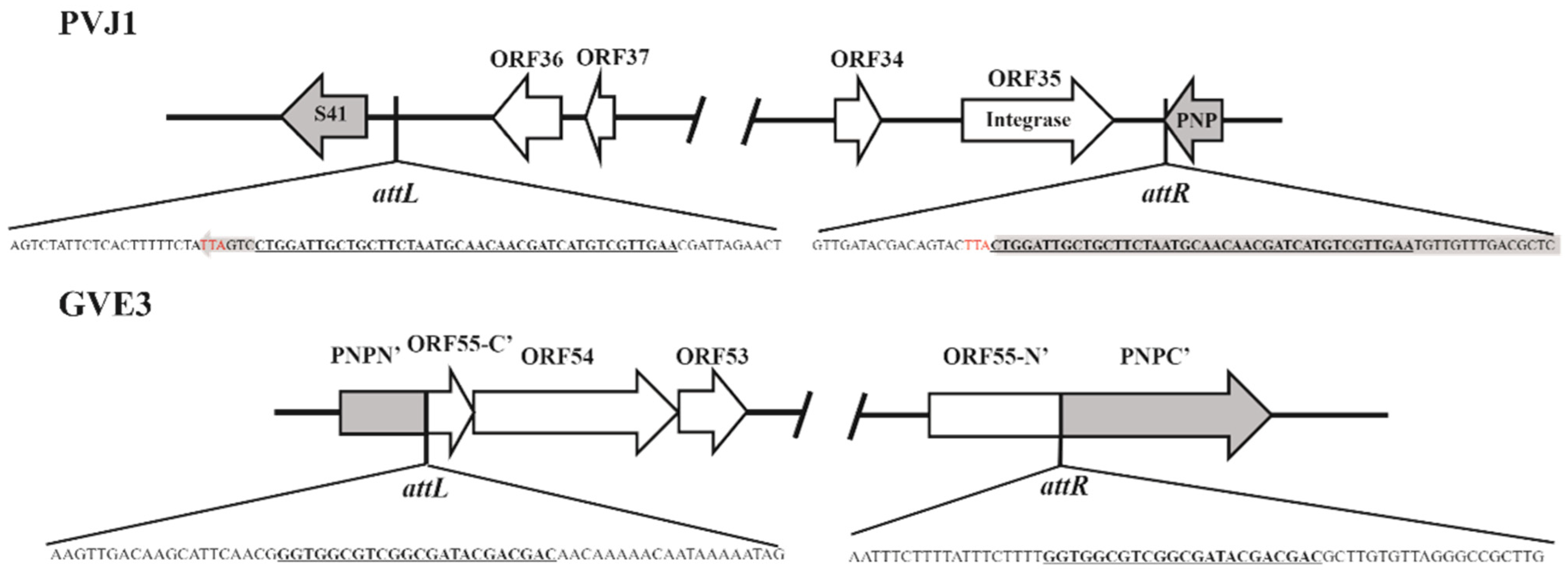
3.4. PVJ1 Integrates into the Host Genome at the Purine Nucleoside Phosphorylase Gene
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cai, L.; Jorgensen, B.B.; Suttle, C.A.; He, M.; Cragg, B.A.; Jiao, N.; Zhang, R. Active and diverse viruses persist in the deep sub-seafloor sediments over thousands of years. ISME J. 2019, 13, 1857–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, T.; Orsi, W.D.; Jorgensen, B.B. Viral activities and life cycles in deep subseafloor sediments. Environ. Microbiol. Rep. 2015, 7, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Liu, W.; Dai, X.; Zhu, Y.; Wang, J.; Zhu, Y.; Zheng, H.; Huang, Y.; Dong, Z.; Du, W.; et al. Extraordinary diversity of viruses in deep-sea sediments as revealed by metagenomics without prior virion separation. Environ. Microbiol. 2021, 23, 728–743. [Google Scholar] [CrossRef]
- Breitbart, M.; Felts, B.; Kelley, S.; Mahaffy, J.M.; Nulton, J.; Salamon, P.; Rohwer, F. Diversity and population structure of a near-shore marine-sediment viral community. Proc. Biol. Sci. 2004, 271, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Serresi, M. Viral density and virus-to-bacterium ratio in deep-sea sediments of the Eastern Mediterranean. Appl. Environ. Microbiol. 2000, 66, 1857–1861. [Google Scholar] [CrossRef] [Green Version]
- Hourdez, S.; Weber, R.E. Molecular and functional adaptations in deep-sea hemoglobins. J. Inorg. Biochem. 2005, 99, 130–141. [Google Scholar] [CrossRef]
- Mercier, C.; Lossouarn, J.; Nesbo, C.L.; Haverkamp, T.H.A.; Baudoux, A.C.; Jebbar, M.; Bienvenu, N.; Thiroux, S.; Dupont, S.; Geslin, C. Two viruses, MCV1 and MCV2, which infect Marinitoga bacteria isolated from deep-sea hydrothermal vents: Functional and genomic analysis. Environ. Microbiol. 2018, 20, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wu, S.; Song, Q.; Zhang, X.; Xie, L. Two novel bacteriophages of thermophilic bacteria isolated from deep-sea hydrothermal fields. Curr. Microbiol. 2006, 53, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, X. Deep-sea thermophilic Geobacillus bacteriophage GVE2 transcriptional profile and proteomic characterization of virions. Appl. Microbiol. Biotechnol. 2008, 80, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X. Genome analysis of deep-sea thermophilic phage D6E. Appl. Environ. Microbiol. 2010, 76, 7861–7866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida-Takashima, Y.; Takaki, Y.; Shimamura, S.; Nunoura, T.; Takai, K. Genome sequence of a novel deep-sea vent epsilonproteobacterial phage provides new insight into the co-evolution of Epsilonproteobacteria and their phages. Extremophiles 2013, 17, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Zablocki, O.; van Zyl, L.J.; Kirby, B.; Trindade, M. Diversity of dsDNA Viruses in a South African Hot Spring Assessed by Metagenomics and Microscopy. Viruses 2017, 9, 348. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, H.W.; Prangishvili, D. Prokaryote viruses studied by electron microscopy. Arch. Virol. 2012, 157, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.E.; Sogin, M.L.; Baross, J.A. Evolutionary strategies of viruses, bacteria and archaea in hydrothermal vent ecosystems revealed through metagenomics. PLoS ONE 2014, 9, e109696. [Google Scholar] [CrossRef] [Green Version]
- Williamson, S.J.; Rusch, D.B.; Yooseph, S.; Halpern, A.L.; Heidelberg, K.B.; Glass, J.I.; Andrews-Pfannkoch, C.; Fadrosh, D.; Miller, C.S.; Sutton, G.; et al. The Sorcerer II Global Ocean Sampling Expedition: Metagenomic characterization of viruses within aquatic microbial samples. PLoS ONE 2008, 3, e1456. [Google Scholar] [CrossRef]
- Ruan, C.J.; Wang, J.; Zheng, X.W.; Song, L.; Zhu, Y.X.; Huang, Y.; Lu, Z.J.; Du, W.B.; Huang, L.; Dai, X. Halovulum marinum sp. nov., isolated from deep-sea water of the Indian Ocean, and emended description of the genus Halovulum. Int. J. Syst. Evol. Microbiol. 2020, 70, 4523–4530. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kubler, J.; Lozajic, M.; Gabler, F.; Soding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, X.; Wang, H.; Jiang, H.; Dong, H.; Huang, L. Novel Sulfolobus Fuselloviruses with Extensive Genomic Variations. J. Virol. 2020, 94, e01624-19. [Google Scholar] [CrossRef]
- Casjens, S.R.; Gilcrease, E.B. Determining DNA packaging strategy by analysis of the termini of the chromosomes in tailed-bacteriophage virions. Methods Mol. Biol. 2009, 502, 91–111. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic. Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Shen, Y.; Fu, Y.; Yu, Y.; Zhao, J.; Li, J.; Li, Y.; Wang, X.; Zhang, J.; Xiang, W. Psychrobacillus lasiicapitis sp. nov., isolated from the head of an ant (Lasius fuliginosus). Int. J. Syst. Evol. Microbiol. 2017, 67, 4462–4467. [Google Scholar] [CrossRef] [PubMed]
- Green, V.; Curtis, F.A.; Sedelnikova, S.; Rafferty, J.B.; Sharples, G.J. Mutants of phage bIL67 RuvC with enhanced Holliday junction binding selectivity and resolution symmetry. Mol. Microbiol. 2013, 89, 1240–1258. [Google Scholar] [CrossRef] [Green Version]
- Frickey, T.; Lupas, A.N. Phylogenetic analysis of AAA proteins. J. Struct. Biol. 2004, 146, 2–10. [Google Scholar] [CrossRef]
- Stamereilers, C.; Fajardo, C.P.; Walker, J.K.; Mendez, K.N.; Castro-Nallar, E.; Grose, J.H.; Hope, S.; Tsourkas, P.K. Genomic Analysis of 48 Paenibacillus larvae Bacteriophages. Viruses 2018, 10, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBlanc, L.; Nezami, S.; Yost, D.; Tsourkas, P.; Amy, P.S. Isolation and characterization of a novel phage lysin active against Paenibacillus larvae, a honeybee pathogen. Bacteriophage 2015, 5, e1080787. [Google Scholar] [CrossRef]
- Young, R. Phage lysis: Three steps, three choices, one outcome. J. Microbiol. 2014, 52, 243–258. [Google Scholar] [CrossRef]
- Pearson, C.R.; Tindall, S.N.; Herman, R.; Jenkins, H.T.; Bateman, A.; Thomas, G.H.; Potts, J.R.; Van der Woude, M.W. Acetylation of Surface Carbohydrates in Bacterial Pathogens Requires Coordinated Action of a Two-Domain Membrane-Bound Acyltransferase. MBio 2020, 11, e01364-20. [Google Scholar] [CrossRef] [PubMed]
- Geagea, H.; Labrie, S.J.; Subirade, M.; Moineau, S. The Tape Measure Protein Is Involved in the Heat Stability of Lactococcus lactis Phages. Appl. Environ. Microbiol. 2018, 84, e02082-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casjens, S.R. Comparative genomics and evolution of the tailed-bacteriophages. Curr. Opin. Microbiol. 2005, 8, 451–458. [Google Scholar] [CrossRef]
- Wu, H.; Sampson, L.; Parr, R.; Casjens, S. The DNA site utilized by bacteriophage P22 for initiation of DNA packaging. Mol. Microbiol. 2002, 45, 1631–1646. [Google Scholar] [CrossRef]
- Buxton, R.S.; Hammer-Jespersen, K.; Hansen, T.D. Insertion of bacteriophage lambda into the deo operon of Escherichia coli K-12 and isolation of plaque-forming lambdadeo+ transducing bacteriophages. J. Bacteriol. 1978, 136, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Zyl, L.J.; Sunda, F.; Taylor, M.P.; Cowan, D.A.; Trindade, M.I. Identification and characterization of a nvel Geobacillus thermoglucosidasius bacteriophage, GVE3. Arch. Virol. 2015, 160, 2269–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.P. Integration sites for genetic elements in prokaryotic tRNA and tmRNA genes: Sublocation preference of integrase subfamilies. Nucleic Acids Res. 2002, 30, 866–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Zheng, X.; Dai, X.; Zhang, Z.; Zhang, W.; Xiao, T.; Huang, L. Isolation and Characterization of the First Temperate Virus Infecting Psychrobacillus from Marine Sediments. Viruses 2022, 14, 108. https://doi.org/10.3390/v14010108
Liu W, Zheng X, Dai X, Zhang Z, Zhang W, Xiao T, Huang L. Isolation and Characterization of the First Temperate Virus Infecting Psychrobacillus from Marine Sediments. Viruses. 2022; 14(1):108. https://doi.org/10.3390/v14010108
Chicago/Turabian StyleLiu, Wang, Xiaowei Zheng, Xin Dai, Zhenfeng Zhang, Wenyan Zhang, Tian Xiao, and Li Huang. 2022. "Isolation and Characterization of the First Temperate Virus Infecting Psychrobacillus from Marine Sediments" Viruses 14, no. 1: 108. https://doi.org/10.3390/v14010108