
Epidemiology and Seasonality of Endemic Human Coronaviruses in South African and Zambian Children: A Case-Control Pneumonia Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case and Control Definitions
2.2. Specimen Collection and Laboratory Testing
2.3. Statistical Analysis
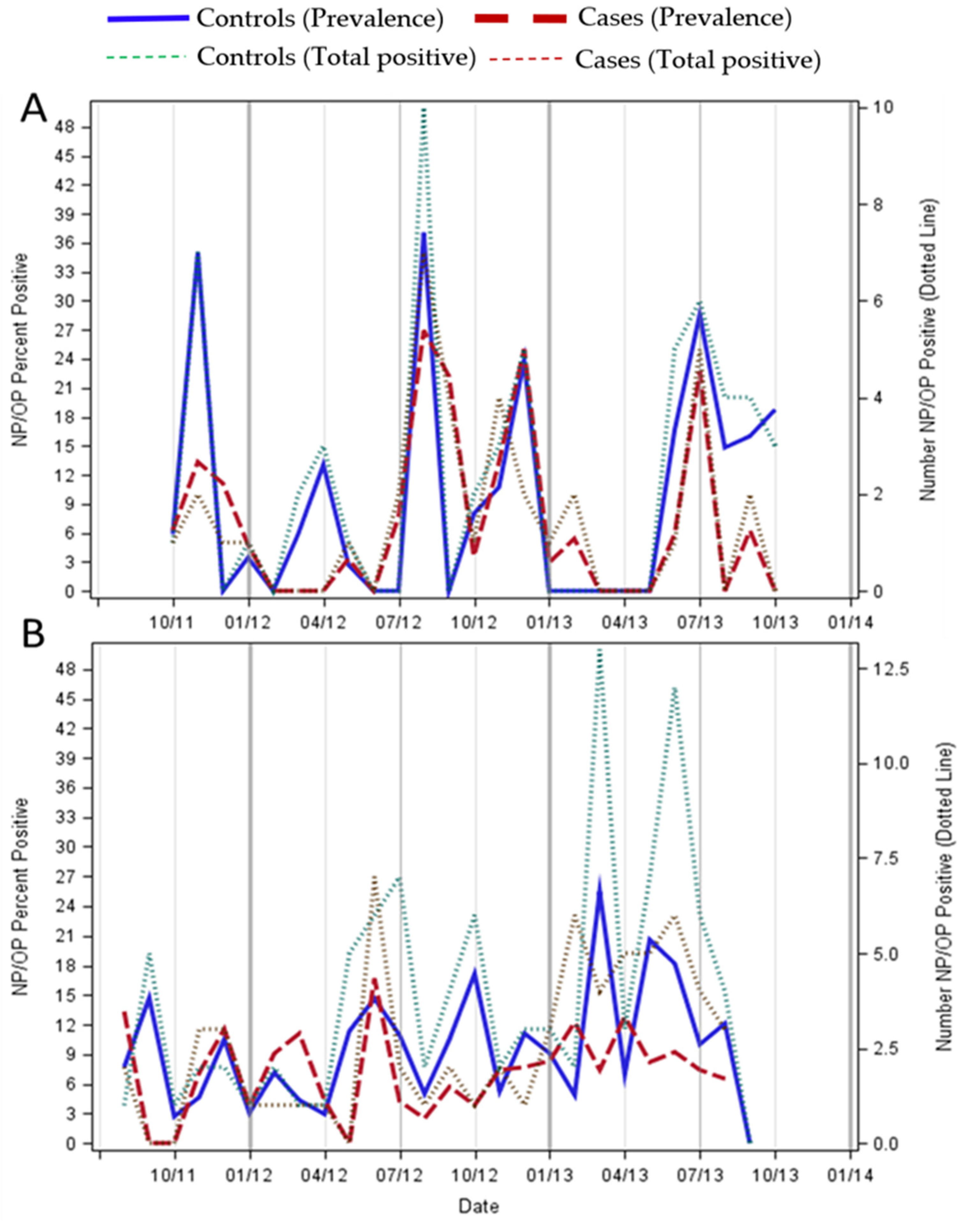
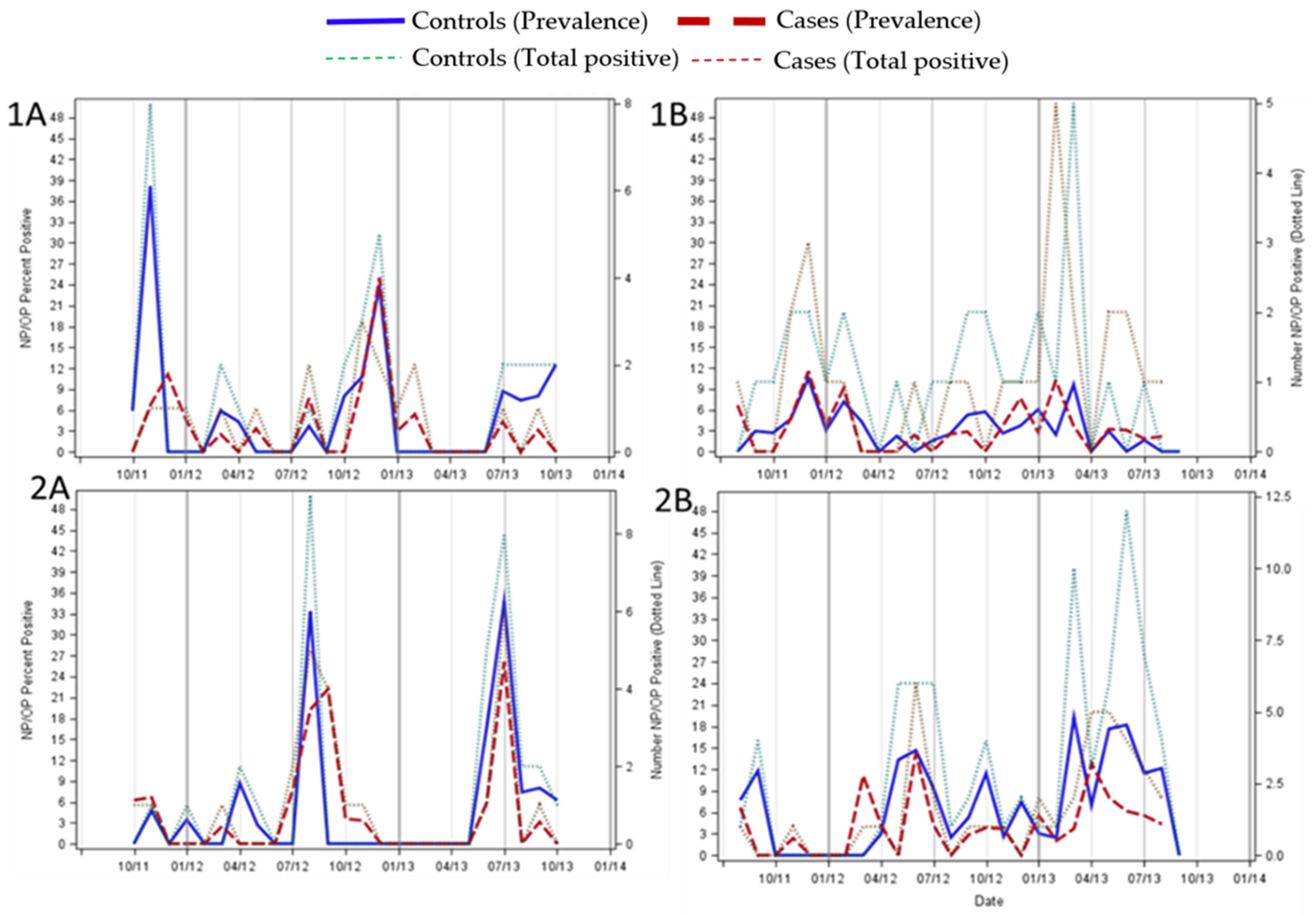
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
- Study Site: South Africa conducted by the Respiratory and Meningeal Pathogens Research Unit, Wits Health Consortium, based at Chris Hani Baragwanath Academic Hospital
- 2.
- Study Site: Lusaka, Zambia at the Boston University Institution
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. In Coronaviruses; Springer: Berlin, Germany, 2015; pp. 1–23. [Google Scholar]
- Cabeça, T.K.; Granato, C.; Bellei, N. Epidemiological and clinical features of human coronavirus infections among different subsets of patients. Influenza Other Respir. Viruses 2013, 7, 1040–1047. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Coronavirus Infections in Children Including COVID-19. Pediatr Infect. Dis J. 2020, 39, 355–368. [Google Scholar] [CrossRef]
- Kuypers, J.; Martin, E.T.; Heugel, J.; Wright, N.; Morrow, R.; Englund, J.A. Clinical disease in children associated with newly described coronavirus subtypes. Pediatrics 2007, 119, e70–e76. [Google Scholar] [CrossRef]
- Baillie, V.L.; Moore, D.P.; Mathunjwa, A.; Morailane, P.; Simões, E.A.; Madhi, S.A. Molecular subtyping of human rhinovirus in children from three sub-Saharan African countries. J. Clin. Microbiol. 2019, 57, e00723-19. [Google Scholar] [CrossRef] [Green Version]
- Levine, O.S.; O’Brien, K.L.; Deloria-Knoll, M.; Murdoch, D.R.; Feikin, D.R.; DeLuca, A.N.; Driscoll, A.J.; Baggett, H.C.; Brooks, W.A.; Howie, S.R. The Pneumonia Etiology Research for Child Health Project: A 21st century childhood pneumonia etiology study. Clin. Infect. Dis. 2012, 54, S93–S101. [Google Scholar] [CrossRef] [Green Version]
- Deloria-Knoll, M.; Feikin, D.R.; Scott, J.A.G.; O’Brien, K.L.; DeLuca, A.N.; Driscoll, A.J.; Levine, O.S.; Group, P.M.W. Identification and selection of cases and controls in the Pneumonia Etiology Research for Child Health project. Clin. Infect. Dis. 2012, 54, S117–S123. [Google Scholar] [CrossRef]
- Murdoch, D.; O’Brien, K.; Driscoll, A.; Karron, R.; Bhat, N. Laboratory methods for determining pneumonia etiology in children. Clin. Infect. Dis. 2012, 54, S146–S152. [Google Scholar] [CrossRef] [Green Version]
- Baggett, H.C.; Watson, N.L.; Deloria Knoll, M.; Brooks, W.A.; Feikin, D.R.; Hammitt, L.L.; Howie, S.R.; Kotloff, K.L.; Levine, O.S.; Madhi, S.A. Density of upper respiratory colonization with Streptococcus pneumoniae and its role in the diagnosis of pneumococcal pneumonia among children aged<5 years in the PERCH study. Clin. Infect. Dis. 2017, 64, S317–S327. [Google Scholar]
- Morpeth, S.C.; Deloria Knoll, M.; Scott, J.A.G.; Park, D.E.; Watson, N.L.; Baggett, H.C.; Brooks, W.A.; Feikin, D.R.; Hammitt, L.L.; Howie, S.R. Detection of pneumococcal DNA in blood by polymerase chain reaction for diagnosing pneumococcal pneumonia in young children from low-and middle-income countries. Clin. Infect. Dis. 2017, 64, S347–S356. [Google Scholar] [CrossRef]
- Prill, M.M.; Iwane, M.K.; Edwards, K.M.; Williams, J.V.; Weinberg, G.A.; Staat, M.A.; Willby, M.J.; Talbot, H.K.; Hall, C.B.; Szilagyi, P.G. Human coronavirus in young children hospitalized for acute respiratory illness and asymptomatic controls. Pediatric Infect. Dis. J. 2012, 31, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Smuts, H.; Workman, L.; Zar, H.J. Role of human metapneumovirus, human coronavirus NL63 and human bocavirus in infants and young children with acute wheezing. J. Med. Virol. 2008, 80, 906–912. [Google Scholar] [CrossRef]
- Taylor, S.; Lopez, P.; Weckx, L.; Borja-Tabora, C.; Ulloa-Gutierrez, R.; Lazcano-Ponce, E.; Kerdpanich, A.; Weber, M.A.R.; de Los Santos, A.M.; Tinoco, J.-C. Respiratory viruses and influenza-like illness: Epidemiology and outcomes in children aged 6 months to 10 years in a multi-country population sample. J. Infect. 2017, 74, 29–41. [Google Scholar] [CrossRef]
- Munywoki, P.K.; Koech, D.C.; Agoti, C.N.; Cane, P.A.; Medley, G.F.; Nokes, D.J. Continuous invasion by respiratory viruses observed in rural households during a respiratory syncytial virus seasonal outbreak in coastal Kenya. Clin. Infect. Dis. 2018, 67, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Smuts, H. Human coronavirus NL63 infections in infants hospitalised with acute respiratory tract infections in South Africa. Influenza Other Respir. Viruses 2008, 2, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Venter, M.; Lassaunière, R.; Kresfelder, T.L.; Westerberg, Y.; Visser, A. Contribution of common and recently described respiratory viruses to annual hospitalizations in children in South Africa. J. Med. Virol. 2011, 83, 1458–1468. [Google Scholar] [CrossRef]
- Madhi, S.A.; Ludewick, H.; Kuwanda, L.; Van Niekerk, N.; Cutland, C.L.; Little, T.; Klugman, K.P. Pneumococcal coinfection with human Metapneumovirus. J. Infect. Dis. 2006, 193, 1236–1943. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan-Gordo, C.; Bassat, Q.; Morais, L.; Díez-Padrisa, N.; Machevo, S.; Nhampossa, T.; Nhalungo, D.; Sanz, S.; Quintó, L.; Alonso, P.L. Etiology and epidemiology of viral pneumonia among hospitalized children in rural Mozambique: A malaria endemic area with high prevalence of human immunodeficiency virus. Pediatric Infect. Dis. J. 2011, 30, 39–44. [Google Scholar] [CrossRef]
- Calvo, C.; Alcolea, S.; Casas, I.; Pozo, F.; Iglesias, M.; Gonzalez-Esguevillas, M.; García-García, M.L. A 14-year prospective study of human coronavirus infections in hospitalized children: Comparison with other respiratory viruses. Pediatric Infect. Dis. J. 2020, 39, 653–657. [Google Scholar] [CrossRef]
- Pretorius, M.A.; Tempia, S.; Walaza, S.; Cohen, A.L.; Moyes, J.; Variava, E.; Dawood, H.; Seleka, M.; Hellferscee, O.; Treurnicht, F.; et al. The role of influenza, RSV and other common respiratory viruses in severe acute respiratory infections and influenza-like illness in a population with a high HIV sero-prevalence, South Africa 2012–2015. J. Clin. Virol. 2016, 75, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Daleno, C.; Scala, A.; Castellazzi, L.; Terranova, L.; Papa, S.S.; Longo, M.; Pelucchi, C.; Principi, N. Impact of rhinovirus nasopharyngeal viral load and viremia on severity of respiratory infections in children. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 41–48. [Google Scholar] [CrossRef]
- Bruning, A.H.; Thomas, X.V.; van der Linden, L.; Wildenbeest, J.G.; Minnaar, R.P.; Jansen, R.R.; de Jong, M.D.; Sterk, P.J.; van der Schee, M.P.; Wolthers, K.C. Clinical, virological and epidemiological characteristics of rhinovirus infections in early childhood: A comparison between non-hospitalised and hospitalised children. J. Clin. Virol. 2015, 73, 120–126. [Google Scholar] [CrossRef]
- Takeyama, A.; Hashimoto, K.; Sato, M.; Sato, T.; Kanno, S.; Takano, K.; Ito, M.; Katayose, M.; Nishimura, H.; Kawasaki, Y. Rhinovirus load and disease severity in children with lower respiratory tract infections. J. Med. Virol. 2012, 84, 1135–1142. [Google Scholar] [CrossRef]
- Ambrosioni, J.; Bridevaux, P.-O.; Aubert, J.-D.; Soccal, P.; Wagner, G.; Kaiser, L. Role of rhinovirus load in the upper respiratory tract and severity of symptoms in lung transplant recipients. J. Clin. Virol. 2015, 64, 1–5. [Google Scholar] [CrossRef]
- Lu, Y.; Li, Y.; Deng, W.; Liu, M.; He, Y.; Huang, L.; Lv, M.; Li, J.; Du, H. Symptomatic infection is associated with prolonged duration of viral shedding in mild coronavirus disease 2019: A retrospective study of 110 children in Wuhan. Pediatric Infect. Dis. J. 2020, 39, e95–e99. [Google Scholar] [CrossRef]
- Heimdal, I.; Moe, N.; Krokstad, S.; Christensen, A.; Skanke, L.H.; Nordbø, S.A.; Døllner, H. Human Coronavirus in Hospitalized Children With Respiratory Tract Infections: A 9-Year Population-Based Study From Norway. J. Infect. Dis. 2019, 219, 1198–1206. [Google Scholar] [CrossRef]
- Monto, A.S.; DeJonge, P.M.; Callear, A.P.; Bazzi, L.A.; Capriola, S.B.; Malosh, R.E.; Martin, E.T.; Petrie, J.G. Coronavirus Occurrence and Transmission Over 8 Years in the HIVE Cohort of Households in Michigan. J. Infect. Dis. 2020, 222, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Fairchok, M.P.; Martin, E.T.; Chambers, S.; Kuypers, J.; Behrens, M.; Braun, L.E.; Englund, J.A. Epidemiology of viral respiratory tract infections in a prospective cohort of infants and toddlers attending daycare. J. Clin. Virol. 2010, 49, 16–20. [Google Scholar] [CrossRef]
- Ofner-Agostini, M.; Wallington, T.; Henry, B.; Low, D.; McDonald, L.; Berger, L.; Mederski, B. Investigation of the second wave (phase 2) of severe acute respiratory syndrome (SARS) in Toronto, Canada. What happened? Can. Commun. Dis. Rep. 2008, 34, 1–11. [Google Scholar]
- Siegfried, N.; van der Merwe, L.; Brocklehurst, P.; Sint, T.T. Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection. Cochrane Database Syst. Rev. 2011. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Group | Total N (%) | HIV+ N (%) | HIV− N (%) | |||
|---|---|---|---|---|---|---|
| Overall | Enrolled | Cases | 1537 | 218 | 1316 | |
| Asymptomatic controls | 1485 | 170 | 1306 | |||
| RTI Controls | 165 | 51 | 113 | |||
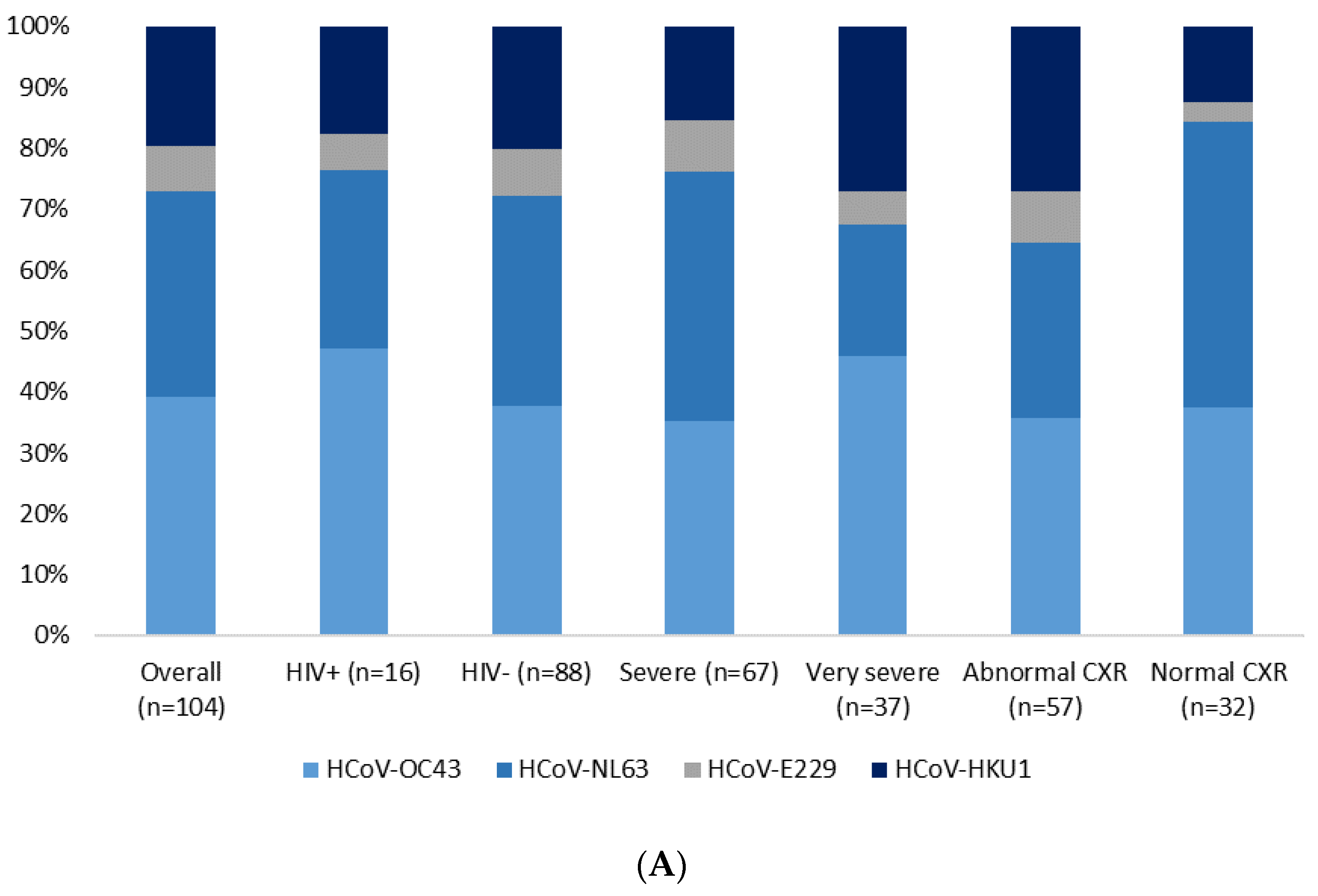
| HCoV g | Cases | 104 (7) | 16 (7) | 88 (7) | ||
| Controls | 164 (11) | 21 (10) | 141 (10) | p = 0.569 f | ||
| p = 0.002 c | p = 0.385 c | p = 0.002 c | ||||
| Asymptomatic control a | 155 (11) | 18 (11) | 135 (11) | p = 0.357 f | ||
| p = 0.001 d | p = 0.320 d | p = 0.001 d | ||||
| RTI control b | 9 (6) | 3 (7) | 6 (6) | p = 0.167 f | ||
| p = 0.698 e | p = 0.817 e | p = 0.738 e | ||||
| AlphaCoV h | Cases | 44 (3) | 6 (3) | 38 (3) | ||
| Controls | 59 (4) | 4 (2) | 54 (4) | p = 0.336 f | ||
| p = 0.232 c | p = 0.362 c | p = 0.140 c | ||||
| Asymptomatic control a | 55 (4) | 1 (2) | 51 (4) | p = 0.258 f | ||
| p = 0.194 d | p = 0.749 d | p = 0.150 d | ||||
| RTI control b | 4 (2) | 3 (2) | 3 (3) | p = 0.538 f | ||
| p = 0.746 e | p = 0.521 e | p = 0.887 e | ||||
| BetaCov i | Cases | 63 (4) | 11 (5) | 52 (4) | ||
| Controls | 111 (7) | 17 (8) | 92 (6) | p = 0.922 f | ||
| p = 0.002 c | p = 0.244 c | p = 0.040 c | ||||
| Asymptomatic control a | 106 (7) | 2 (4) | 89 (7) | p = 0.217 f | ||
| p = 0.002 d | p = 0.736 d | p = 0.001 d | ||||
| RTI control b | 5 (3) | 15 (9) | 3 (3) | p = 0.477 f | ||
| p = 0.505 e | p = 0.140 e | p = 0.492 e |
| Characteristics | Cases N (%) | Asymptomatic Controls N (%) | aOR (95%CI) | p-Value a |
|---|---|---|---|---|
| HCoV epidemiology | ||||
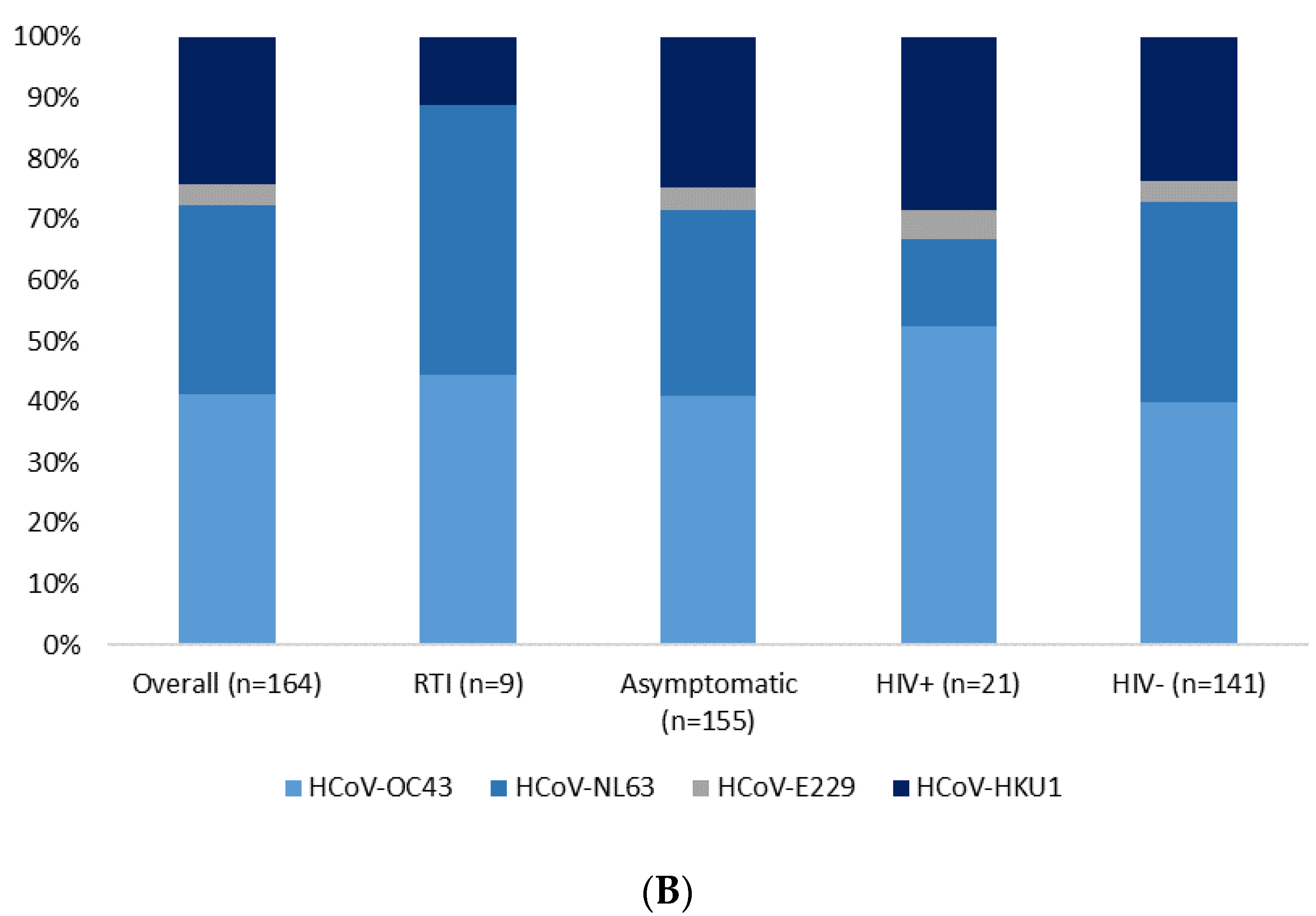
| HCoV detected as single viral infections b | 38 (36) | 89 (57) | 2.64 (1.55–4.53) | <0.001 |
| HCoV-E229 | 8 (8) | 6 (4) | 0.49 (0.16–1.49) | 0.206 |
| HCoV-NL63 | 36 (35) | 49 (32) | 0.91 (0.53–1.56) | 0.730 |
| HCoV-OC43 | 42 (40) | 66 (43) | 1.07 (0.63–1.81) | 0.802 |
| HCoV-HKU1 | 21 (20) | 40 (26) | 1.32 (0.72–2.44) | 0.367 |
| HCoV load, mean (SD) d | 5.29 (0.77) | 5.29 (0.68) | - | 0.973 |
| Load in HCoV single infections | 5.43 (0.27) | 5.33 (0.17) | - | 0.77 |
| Load in HCoV mixed infections | 5.18 (0.22) | 5.23 (0.20) | - | 0.87 |
| Mixed viral infections in the NP/OP | ||||
| Two HCoV species c | 3 (3) | 2 (1) | 0.59 (0.22–2.45) | 0.39 |
| RSV | 24 (23) | 2 (1) | 0.05 (0.01–0.21) | <0.001 |
| InFV (A, B and C) | 4 (4) | 2 (1) | 0.22 (0.03–1.40) | 0.108 |
| AdV | 14 (13) | 10 (6) | 0.34 (0.14–0.82) | 0.017 |
| HBoV | 19 (18) | 17 (11) | 0.47 (0.22–0.98) | 0.045 |
| RV | 20 (19) | 28 (18) | 0.92 (0.48–1.76) | 0.806 |
| PIV (1–4) | 2 (2) | 4 (3) | 1.26 (0.21–7.40) | 0.797 |
| Bacterial co-infections in the NP/OP | ||||
| H. influenzae type b | 5 (5) | 2 (1) | 0.24 (0.05–1.33) | 0.104 |
| B. pertussis | 2 (2) | 1 (1) | 0.41 (0.04–4.72) | 0.473 |
| S. aureus | 21 (20) | 23 (15) | 0.86 (0.43–1.71) | 0.658 |
| H. influenzae | 52 (50) | 73 (47) | 0.79 (0.47–1.33) | 0.377 |
| S. pneumoniae | 74 (71) | 114 (74) | 1.09 (0.61–1.95) | 0.773 |
| M. pneumoniae | 1 (1) | 3 (2) | 1.27 (0.12–13.5) | 0.840 |
| M. catarrhalis | 71 (68) | 116 (75) | 1.39 (0.79–2.47) | 0.257 |
| C. pneumoniae | 0 | 5 (3) | - | 0.082 |
| HCoV and S. pneumoniae co-infections | ||||
| S. pneu load, mean (SD) e | 5.42 (0.16) | 5.77 (1.37) | - | 0.067 |
| HDP in nasopharynx f | 15 (14) | 11 (7) | 2.46 (1.06–5.67) | 0.035 |
| S. pneu detected in WB | 5 (5) | 11 (7) | 1.38 (0.45–4.25) | 0.570 |
| Characteristics | HCoV a + (n = 104) N(%) | AlphaCoV b (n = 44) N(%) | BetaCoV c (n = 63) N(%) | HCoV- (n = 1338) N(%) | p-Value d | p-Value e | p-Value f |
|---|---|---|---|---|---|---|---|
| Demographics and health | |||||||
| Age (months), mean (SD) | 7.9 (8.52) | 9.07 (9.55) | 7.51 (8.05) | 8.9 (10.01) | 0.337 | 0.917 | 0.276 |
| Male | 64 (62) | 27 (61) | 39 (62) | 703 (52) | 0.07 | 0.241 | 0.136 |
| HIV | 16 (15) | 6 (14) | 11 (17) | 188 (14) | 0.654 | 0.921 | 0.409 |
| Clinical features | |||||||
| Very severe pneumonia | 37 (35) | 10 (23) | 27 (43) | 428 (32) | 0.463 | 0.204 | 0.08 |
| Chest X-ray abnormal g | 57 (58) | 22 (53) | 37 (63) | 693 (54) | 0.583 | 0.712 | 0.26 |
| Hypoxic h | 69 (67) | 27 (63) | 44 (70) | 829 (62) | 0.274 | 0.807 | 0.175 |
| Tachycardia i | 59 (57) | 29 (67) | 33 (53) | 737 (55) | 0.696 | 0.111 | 0.606 |
| Tachypnea j | 83 (82) | 38 (86) | 48 (79) | 1081 (82) | 0.999 | 0.439 | 0.607 |
| Wheezing | 24 (23) | 11 (26) | 14 (22) | 321 (24) | 0.948 | 0.785 | 0.855 |
| Convulsions | 2 (2) | 1 (2) | 1 (2) | 38 (3) | 0.613 | 0.793 | 0.582 |
| Diarrhea | 16 (15) | 6 (14) | 10 (16) | 238 (18) | 0.485 | 0.473 | 0.636 |
| Hospital stay > 5 days | 59 (57) | 20 (45) | 39 (62) | 665 (50) | 0.174 | 0.619 | 0.06 |
| Died in Hospital | 9 (9) | 4 (9) | 5 (8) | 127 (9) | 0.755 | 0.912 | 0.643 |
| Bacterial co-infection markers | |||||||
| Fever k | 74 (71) | 30 (68) | 46 (73) | 963 (72) | 0.947 | 0.573 | 0.782 |
| Leukocytosis l | 593 (45) | 51 (50) | 19 (44) | 34 (54) | 0.302 | 0.925 | 0.122 |
| CRP > 40 mg/L m | 35 (34) | 12 (27) | 25 (40) | 381 (28) | 0.186 | 0.848 | 0.034 |
| Blood culture positive n | 8 (8) | 3 (7) | 5 (8) | 47 (4) | 0.039 | 0.266 | 0.084 |
| MCPP o | 1 (1) | 1 (2) | 0 | 11 (1) | 0.829 | 0.339 | 0.47 |
| Bacterial co-infections | |||||||
| S. aureus | 21 (20) | 11 (25) | 10 (16) | 333 (25) | 0.291 | 0.911 | 0.111 |
| S. pneumoniae | 74 (71) | 29 (66) | 48 (76) | 919 (68) | 0.571 | 0.662 | 0.202 |
| M. catarrhalis | 71 (68) | 26 (59) | 47 (75) | 850 (63) | 0.314 | 0.500 | 0.071 |
| B. pertussis | 2 (2) | 0 | 2 (3) | 19 (1) | 0.678 | 0.417 | 0.233 |
| H. influenzae | 52 (50) | 18 (41) | 37 (59) | 669 (50) | 0.977 | 0.226 | 0.152 |
| H. influenzae type b | 5 (5) | 1 (2) | 4 (6) | 38 (3) | 0.254 | 0.757 | 0.122 |
| M. pneumoniae | 1 (1) | 0 | 1 (2) | 7 (1) | 0.561 | 0.615 | 0.258 |
| C. pneumoniae | 0 | 0 | 0 | 16 (1) | 0.263 | 0.480 | 0.395 |
| HDP in nasopharynx p | 15 (14) | 3 (7) | 13 (21) | 144 (11) | 0.245 | 0.436 | 0.006 |
| HDP in blood q | 4 (4) | 3 (7) | 1 (2) | 67 (5) | 0.603 | 0.529 | 0.222 |
| Respiratory viral co-infections in the Nasopharynx | |||||||
| RSV | 24 (23) | 7 (16) | 18 (29) | 320 (24) | 0.76 | 0.234 | 0.478 |
| AdV | 14 (13) | 6 (14) | 9 (14) | 130 (10) | 0.165 | 0.375 | 0.172 |
| HMPV | 3 (3) | 2 (5) | 2 (3) | 94 (7) | 0.117 | 0.529 | 0.249 |
| HBoV | 19 (18) | 8 (18) | 12 (19) | 152 (11) | 0.024 | 0.166 | 0.045 |
| InFV A-C | 4 (4) | 2 (5) | 2 (3) | 59 (5) | 0.618 | 0.861 | 0.546 |
| PIV | 2 (2) | 1 (2) | 1 (2) | 138 (10) | 0.014 | 0.117 | 0.05 |
| RV | 20 (19) | 6 (14) | 15 (24) | 303 (23) | 0.460 | 0.167 | 0.770 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baillie, V.L.; Moore, D.P.; Mathunjwa, A.; Park, D.E.; Thea, D.M.; Kwenda, G.; Mwananyanda, L.; Madhi, S.A. Epidemiology and Seasonality of Endemic Human Coronaviruses in South African and Zambian Children: A Case-Control Pneumonia Study. Viruses 2021, 13, 1513. https://doi.org/10.3390/v13081513
Baillie VL, Moore DP, Mathunjwa A, Park DE, Thea DM, Kwenda G, Mwananyanda L, Madhi SA. Epidemiology and Seasonality of Endemic Human Coronaviruses in South African and Zambian Children: A Case-Control Pneumonia Study. Viruses. 2021; 13(8):1513. https://doi.org/10.3390/v13081513
Chicago/Turabian StyleBaillie, Vicky L., David P. Moore, Azwifarwi Mathunjwa, Daniel E. Park, Donald M. Thea, Geoffrey Kwenda, Lawrence Mwananyanda, and Shabir A. Madhi. 2021. "Epidemiology and Seasonality of Endemic Human Coronaviruses in South African and Zambian Children: A Case-Control Pneumonia Study" Viruses 13, no. 8: 1513. https://doi.org/10.3390/v13081513