
Confound, Cause, or Cure: The Effect of Cannabinoids on HIV-Associated Neurological Sequelae

{kind=link}
Abstract
:1. Introduction
1.1. HIV Infection in the CNS and Neurotoxic Sequelae
1.2. The Endocannabinoid System and Cannabinoid Pharmacology
1.3. Method of Literature Review
2. Cannabinoids × HIV: Prevalence, Cross Talk, and Clinical Outcomes
2.1. Purpose and Prevalence of Exocannabinoid Use in PLWH
2.2. Effect of HIV Infection on the Endocannabinoid System
2.3. Effect of Cannabinoids on HIV Infection Dynamics and Treatment Adherence
2.4. Effect of Cannabinoids on Central and Peripheral Inflammation and Immunity in HIV
2.5. Cannabinoids as a Modifier for HAND Risk
3. Cannabinoids × HIV: Receptor- and Cell-Specific Mechanisms in Preclinical Models
3.1. CB1-Mediated Effects on HIV Neuropathology
3.2. CB2-Mediated Effects on HIV Neuropathology
3.2.1. Viral Replication and Immunogenicity
3.2.2. Inflammation and Immune Response
3.2.3. Neurotoxicity
3.3. BBB Integrity
3.4. Other Putative Cannabinoid Receptors
3.5. Endocannabinoid Enzyme Modification
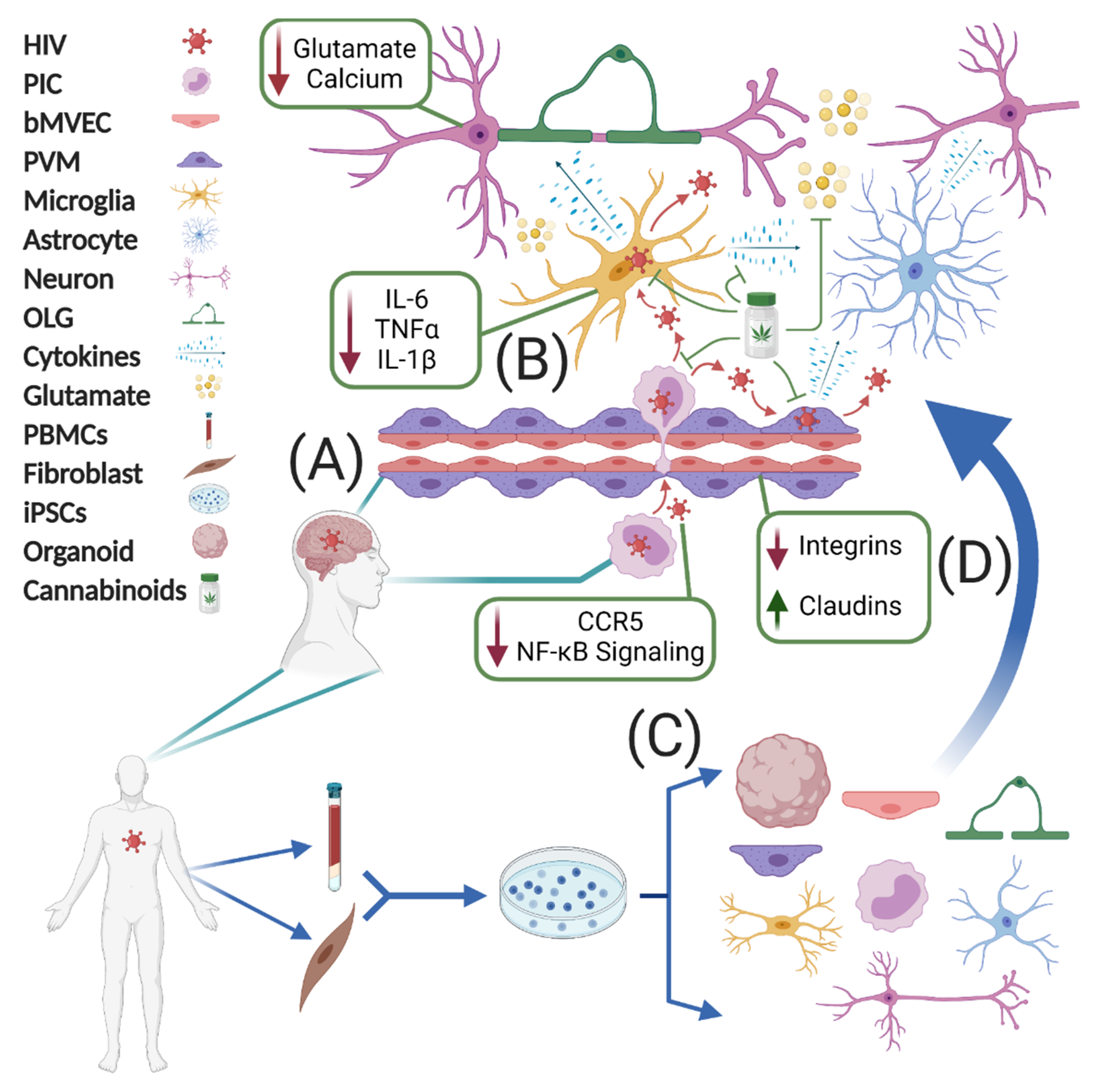
4. Human iPSC Modeling of HIV in the CNS
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UNAIDS. UNAIDS 2020 Fact Sheet; UNAIDS: Geneva, Switzerland, 2020. [Google Scholar]
- Sakamoto, M.; Marcotte, T.D.; Umlauf, A.; Franklin, D.; Heaton, R.K.; Ellis, R.J.; Letendre, S.; Alexander, T.; McCutchan, J.A.; Morgan, E.E.; et al. Concurrent Classification Accuracy of the HIV Dementia Scale for HIV-Associated Neurocognitive Disorders in the CHARTER Cohort. J. Acquir. Immune Defic. Syndr. 2013, 62, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, R.K.; Franklin, D.R.; Ellis, R.J.; McCutchan, J.A.; Letendre, S.L.; Leblanc, S.; Corkran, S.H.; Duarte, N.A.; Clifford, D.B.; Woods, S.P.; et al. HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: Differences in rates, nature, and predictors. J. NeuroVirol. 2011, 17, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Saylor, D.; Dickens, A.; Sacktor, N.; Haughey, N.; Slusher, B.; Pletnikov, M.; Mankowski, J.L.; Brown, A.; Volsky, D.J.; McArthur, D. HIV-associated neurocognitive disorder—pathogenesis and prospects for treatment. Nat. Rev. Neurol. 2016, 12, 234–248. [Google Scholar] [CrossRef]
- Cysique, L.A.; Maruff, P.; Brew, B.J. Prevalence and pattern of neuropsychological impairment in human immunodeficiency vi-rus-infected/acquired immunodeficiency syndrome (HIV/AIDS) patients across pre- and post-highly active antiretroviral therapy eras: A combined study of two cohorts. J. Neurovirol. 2004, 10, 350–357. [Google Scholar] [CrossRef]
- Amusan, P.; Power, C.; Gill, M.J.; Gomez, D.; Johnson, E.; Rubin, L.H.; Fujiwara, E. Lifetime antiretroviral exposure and neurocognitive impairment in HIV. J. NeuroVirology 2020, 26, 743–753. [Google Scholar] [CrossRef]
- Gannon, P.J.; Akay-Espinoza, C.; Yee, A.C.; Briand, L.A.; Erickson, M.A.; Gelman, B.B.; Gao, Y.; Haughey, N.J.; Zink, M.C.; Clements, J.E.; et al. HIV Protease Inhibitors Alter Amyloid Precursor Protein Processing via beta-Site Amyloid Precursor Protein Cleaving Enzyme-1 Translational Up-Regulation. Am. J. Pathol. 2017, 187, 91–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akay, C.; Cooper, M.; Odeleye, A.; Jensen, B.; White, M.; Vassoler, F.; Gannon, P.J.; Mankowski, J.; Dorsey, J.L.; Buch, A.M.; et al. Antiretroviral drugs induce oxidative stress and neuronal damage in the central nervous system. J. NeuroVirol. 2014, 20, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Ryan, S.K.; Gonzalez, M.V.; Garifallou, J.P.; Bennett, F.C.; Williams, K.S.; Sotuyo, N.P.; Mironets, E.; Cook, K.; Hakonarson, H.; Anderson, S.A.; et al. Neuroinflammation and EIF2 Signaling Persist despite Antiretroviral Treatment in an hiPSC Tri-culture Model of HIV Infection. Stem Cell Rep. 2020, 14, 703–716. [Google Scholar] [CrossRef]
- Stern, A.L.; Lee, R.N.; Panvelker, N.; Li, J.; Harowitz, J.; Jordan-Sciutto, K.L.; Akay-Espinoza, C. Differential Effects of Antiretroviral Drugs on Neurons In Vitro: Roles for Oxidative Stress and Integrated Stress Response. J. Neuroimmune Pharmacol. 2017, 13, 64–76. [Google Scholar] [CrossRef]
- Jensen, B.K.; Monnerie, H.; Mannell, M.V.; Gannon, P.J.; Espinoza, C.A.; Erickson, M.A.; Bruce-Keller, A.J.; Gelman, B.B.; Briand, L.A.; Pierce, R.C.; et al. Altered Oligodendrocyte Maturation and Myelin Maintenance: The Role of Antiretrovirals in HIV-Associated Neu-rocognitive Disorders. J. Neuropathol. Exp. Neurol. 2015, 74, 1093–1118. [Google Scholar] [CrossRef] [Green Version]
- De Benedetto, I.; Trunfio, M.; Guastamacchia, G.; Bonora, S.; Calcagno, A. A review of the potential mechanisms of neuronal toxicity associated with antiretroviral drugs. J. NeuroVirol. 2020, 26, 642–651. [Google Scholar] [CrossRef]
- Kallianpur, K.J.; Birn, R.; Ndhlovu, L.C.; Souza, S.A.; Mitchell, B.; Paul, R.; Chow, D.C.; Kohorn, L.; Shikuma, C.M. Impact of Cannabis Use on Brain Structure and Function in Suppressed HIV Infection. J. Behav. Brain Sci. 2020, 10, 344–370. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, L.; Bagot, K.; Brown, A.; Haeny, A.M. The Association Between Marijuana Use and HIV Continuum of Care Outcomes: A Systematic Review. Curr. HIV/AIDS Rep. 2019, 16, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Thames, A.D.; Mahmood, Z.; Burggren, A.C.; Karimian, A.; Kuhn, T.P. Combined effects of HIV and marijuana use on neurocognitive functioning and immune status. AIDS Care 2016, 28, 628–632. [Google Scholar] [CrossRef]
- Thames, A.D.; Kuhn, T.P.; Williamson, T.J.; Jones, J.; Mahmood, Z.; Hammond, A. Marijuana effects on changes in brain structure and cognitive function among HIV+ and HIV− adults. Drug Alcohol Depend. 2017, 170, 120–127. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, G.; Matson, P.A.; Grady, C.D.; Nahvi, S.; Merenstein, D.; Weber, K.M.; Greenblatt, R.; Burian, P.; Wilson, T.E. Medicinal and recreational marijuana use among HIV-infected women in the Women’s Interagency HIV Study (WIHS) cohort, 1994–2010. J. Acquir. Immune Defic. Syndr. 2012, 61, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Tovanabutra, S.; Sirijatuphat, R.; Pham, P.T.; Bonar, L.; Harbolick, E.A.; Bose, M.; Song, H.; Chang, D.; Oropeza, C.; O’Sullivan, A.M.; et al. Deep Sequencing Reveals Central Nervous System Compartmentalization in Multiple Transmitted/Founder Virus Acute HIV-1 Infection. Cells 2019, 8, 902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spudich, S.; Peterson, J.; Fuchs, D.; Price, R.W.; Gisslen, M. Potential for early antiretroviral therapy to reduce central nervous system HIV-1 persistence. AIDS 2019, 33, S135–S144. [Google Scholar] [CrossRef]
- Williams, D.; Veenstra, M.; Gaskill, P.J.; Morgello, S.; Calderon, T.M.; Berman, J.W. Monocytes mediate HIV neuropathogenesis: Mechanisms that contribute to HIV associated neu-rocognitive disorders. Curr. HIV Res. 2014, 12, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Cassol, E.; Alfano, M.; Biswas, P.; Poli, G. Monocyte-derived macrophages and myeloid cell lines as targets of HIV-1 replication and persistence. J. Leukoc. Biol. 2006, 80, 1018–1030. [Google Scholar] [CrossRef]
- Valdebenito, S.; Castellano, P.; Ajasin, D.; Eugenin, E.A. Astrocytes are HIV reservoirs in the brain: A cell type with poor HIV infectivity and replication but efficient cell-to-cell viral transfer. J. Neurochem. 2021. [Google Scholar] [CrossRef]
- Gill, A.J.; Kovacsics, C.E.; Vance, P.J.; Collman, R.G.; Kolson, D.L. Induction of Heme Oxygenase-1 Deficiency and Associated Glutamate-Mediated Neurotoxicity Is a Highly Conserved HIV Phenotype of Chronic Macrophage Infection That Is Resistant to Antiretroviral Therapy. J. Virol. 2015, 89, 10656–10667. [Google Scholar] [CrossRef] [Green Version]
- Zyskind, J.W.; Wang, Y.; Cho, G.; Ting, J.H.; Kolson, D.L.; Lynch, D.R.; Jordan-Sciutto, K.L. E2F1 in neurons is cleaved by calpain in an NMDA receptor-dependent manner in a model of HIV-induced neu-rotoxicity. J. Neurochem. 2015, 132, 742–755. [Google Scholar] [CrossRef] [Green Version]
- Cross, S.A.; Cook, D.R.; Chi, A.W.S.; Vance, P.J.; Kolson, L.L.; Wong, B.J.; Jordan-Sciutto, K.L.; Kolson, D.L. Dimethyl Fumarate, an Immune Modulator and Inducer of the Antioxidant Response, Suppresses HIV Replication and Macrophage-Mediated Neurotoxicity: A Novel Candidate for HIV Neuroprotection. J. Immunol. 2011, 187, 5015–5025. [Google Scholar] [CrossRef] [Green Version]
- Gill, A.; Kovacsics, C.E.; Cross, S.A.; Vance, P.J.; Kolson, L.L.; Jordan-Sciutto, K.L.; Gelman, B.B.; Kolson, D.L. Heme oxygenase-1 deficiency accompanies neuropathogenesis of HIV-associated neurocognitive disorders. J. Clin. Investig. 2014, 124, 4459–4472. [Google Scholar] [CrossRef] [Green Version]
- Gelman, B.B. Neuropathology of HAND with Suppressive Antiretroviral Therapy: Encephalitis and Neurodegeneration Reconsidered. Curr. HIV/AIDS Rep. 2015, 12, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Basavarajappa, B.S.; Shivakumar, M.; Joshi, V.; Subbanna, S. Endocannabinoid system in neurodegenerative disorders. J. Neurochem. 2017, 142, 624–648. [Google Scholar] [CrossRef] [PubMed]
- El Sohly, M.A.; Slade, D. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci. 2005, 78, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Vergara, D.; Bidwell, L.C.; Gaudino, R.; Torres, A.; Du, G.; Ruthenburg, T.C.; DeCesare, K.; Land, D.P.; Hutchison, K.E.; Kane, N.C. Compromised External Validity: Federally Produced Cannabis Does Not Reflect Legal Markets. Sci. Rep. 2017, 7, srep46528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soethoudt, M.; Grether, U.; Fingerle, J.; Grim, T.W.; Fezza, F.; De Petrocellis, L.; Ullmer, C.; Rothenhäusler, B.; Perret, C.; Van Gils, N.; et al. Cannabinoid CB2 receptor ligand profiling reveals biased signalling and off-target activity. Nat. Commun. 2017, 8, 13958. [Google Scholar] [CrossRef]
- Prentiss, D.; Power, R.; Balmas, G.; Tzuang, G.; Israelski, D.M. Patterns of Marijuana Use Among Patients With HIV/AIDS Followed in a Public Health Care Setting. J. Acquir. Immune Defic. Syndr. 2004, 35, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolridge, E.; Barton, S.; Samuel, J.; Osorio, J.; Dougherty, A.; Holdcroft, A. Cannabis Use in HIV for Pain and Other Medical Symptoms. J. Pain Symptom Manag. 2005, 29, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Sandfort, T.G.M.; Knox, J.; Alcala, C.; El-Bassel, N.; Kuo, I.; Smith, L.R. Substance Use and HIV Risk Among Men Who Have Sex with Men in Africa: A Systematic Review. J. Acquir. Immune Defic. Syndr. 2017, 76, e34–e46. [Google Scholar] [CrossRef]
- Furler, M.D.; Einarson, T.R.; Millson, M.; Walmsley, S.; Bendayan, R. Medicinal and Recreational Marijuana Use by Patients Infected with HIV. AIDS Patient Care STDs 2004, 18, 215–228. [Google Scholar] [CrossRef]
- Deimel, D.; Stöver, H.; Hößelbarth, S.; Dichtl, A.; Graf, N.; Gebhardt, V. Drug use and health behaviour among German men who have sex with men: Results of a qualitative, multi-centre study. Harm Reduct. J. 2016, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin, N.; Velasco, C.; De Pourcq, J.T.; Lopez, B.; Gutierrez, M.D.M.; Haro, J.M.; Feliu, A.; Mangues, M.A.; Trilla, A. Corrigendum: Recreational drug use among individuals living with HIV in Europe: Review of the prevalence, comparison with the general population and HIV guidelines recommendations. Front. Microbiol. 2015, 6, 1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacek, L.R.; Towe, S.L.; Hobkirk, A.L.; Nash, D.; Goodwin, R.D. Frequency of Cannabis Use and Medical Cannabis Use Among Persons Living with HIV in the United States: Findings from a Nationally Representative Sample. AIDS Educ. Prev. 2018, 30, 169–181. [Google Scholar] [CrossRef]
- Grabovac, I.; Meilinger, M.; Schalk, H.; Leichsenring, B.; Dorner, T.E. Prevalence and Associations of Illicit Drug and Polydrug Use in People Living with HIV in Vienna. Sci. Rep. 2018, 8, 8046. [Google Scholar] [CrossRef]
- Hartzler, B.; Dombrowski, J.C.; Crane, H.M.; Eron, J.J.; Geng, E.H.; Mathews, W.C.; Mayer, K.H.; Moore, R.D.; Mugavero, M.J.; Napravnik, S.; et al. Prevalence and Predictors of Substance Use Disorders Among HIV Care Enrollees in the United States. AIDS Behav. 2017, 21, 1138–1148. [Google Scholar] [CrossRef]
- Ware, M.A.; Rueda, S.; Singer, J.; Kilby, D. Cannabis Use by Persons Living with HIV/AIDS: Patterns and Prevalence of Use. J. Cannabis Ther. 2003, 3, 3–15. [Google Scholar] [CrossRef]
- Sohler, N.L.; Starrels, J.L.; Khalid, L.; Bachhuber, M.; Arnsten, J.H.; Nahvi, S.; Jost, J.; Cunningham, C.O. Cannabis Use is Associated with Lower Odds of Prescription Opioid Analgesic Use Among HIV-Infected Individuals with Chronic Pain. Subst. Use Misuse 2018, 53, 1602–1607. [Google Scholar] [CrossRef]
- Hartzler, B.; Carlini, B.H.; Newville, H.; Crane, H.M.; Eron, J.J.; Geng, E.H.; Mathews, W.C.; Mayer, K.H.; Moore, R.D.; Mugavero, M.J.; et al. Identifying HIV care enrollees at-risk for cannabis use disorder. AIDS Care 2017, 29, 846–850. [Google Scholar] [CrossRef]
- Haney, M.; Gunderson, E.W.; Rabkin, J.; Hart, C.L.; Vosburg, S.K.; Comer, S.D.; Foltin, R.W. Dronabinol and Marijuana in HIV-Positive Marijuana Smokers: Caloric Intake, Mood, and Sleep. JAIDS J. Acquir. Immune Defic. Syndr. 2007, 45, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struwe, M.; Kaempfer, S.H.; Geiger, C.J.; Pavia, A.T.; Plasse, T.F.; Shepard, K.V.; Ries, K.; Evans, T.G. Effect of Dronabinol on Nutritional Status in HIV Infection. Ann. Pharmacother. 1993, 27, 827–831. [Google Scholar] [CrossRef]
- Riggs, P.K.; Vaida, F.; Rossi, S.S.; Sorkin, L.S.; Gouaux, B.; Grant, I.; Ellis, R.J. A pilot study of the effects of cannabis on appetite hormones in HIV-infected adult men. Brain Res. 2012, 1431, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Lutge, E.; Gray, A.; Siegfried, N. The medical use of cannabis for reducing morbidity and mortality in patients with HIV/AIDS. Cochrane Database Syst. Rev. 2013, CD005175. [Google Scholar] [CrossRef]
- Abrams, D.I.; Jay, C.A.; Shade, S.B.; Vizoso, H.; Reda, H.; Press, S.; Kelly, M.E.; Rowbotham, M.C.; Petersen, K.L. Cannabis in painful HIV-associated sensory neuropathy: A randomized placebo-controlled trial. Neurology 2007, 68, 515–521. [Google Scholar] [CrossRef]
- Ellis, R.J.; Toperoff, W.; Vaida, F.; Brande, G.V.D.; Gonzales, J.; Gouaux, B.; Bentley, H.; Atkinson, J.H. Smoked Medicinal Cannabis for Neuropathic Pain in HIV: A Randomized, Crossover Clinical Trial. Neuropsychopharmacology 2008, 34, 672–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, T.J.C.; Cherry, C.L.; Cox, S.; Marshall, S.J.; Rice, A.S.C. Pharmacological Treatment of Painful HIV-Associated Sensory Neuropathy: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. PLoS ONE 2010, 5, e14433. [Google Scholar] [CrossRef] [PubMed]
- Eibach, L.; Scheffel, S.; Cardebring, M.; Lettau, M.; Celik, M.Ö.; Morguet, A.; Roehle, R.; Stein, C. Cannabidivarin for HIV-Associated Neuropathic Pain: A Randomized, Blinded, Controlled Clinical Trial. Clin. Pharmacol. Ther. 2021, 109, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Wardell, J.D.; Shuper, P.A.; Hendershot, C.S. A longitudinal investigation of the association between cannabis use and alcohol use among people living with HIV. Drug Alcohol Depend. 2018, 193, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tyurina, A.; Krupitsky, E.; Cheng, D.; Coleman, S.M.; Walley, A.Y.; Bridden, C.; Gnatienko, N.; Zvartau, E.; Raj, A.; Samet, J.H. Is cannabis use associated with HIV drug and sex risk behaviors among Russian HIV-infected risky drinkers? Drug Alcohol Depend. 2013, 132, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Gorbach, P.M.; Gorbach, P.M.; Javanbakht, M.; Shover, C.L.; Bolan, R.K.; Ragsdale, A.; Shoptaw, S. Associations Between Cannabis Use, Sexual Behavior, and Sexually Transmitted Infections/Human Immunodefi-ciency Virus in a Cohort of Young Men Who Have Sex with Men. Sex Transm. Dis. 2019, 46, 105–111. [Google Scholar] [CrossRef]
- Coates, R.A.; Read, S.E.; MacFadden, D.K.; Calzavara, L.M.; Johnson, J.K.; Fanning, M.M.; Farewell, V.T.; Raboud, J.; Shepherd, F.A. Cofactors of Progression to Acquired Immunodeficiency Syndrome in A Cohort of Male Sexual Contacts of Men with Human Immunodeficiency Virus Disease. Am. J. Epidemiol. 1990, 132, 717–722. [Google Scholar] [CrossRef]
- Adams, J.W.; Bryant, K.J.; Edelman, J.E.; Fiellin, D.A.; Gaither, J.R.; Gordon, A.J.; Gordon, K.S.; Kraemer, K.L.; Mimiaga, M.J.; Operario, D.; et al. Association of Cannabis, Stimulant, and Alcohol use with Mortality Prognosis Among HIV-Infected Men. AIDS Behav. 2018, 22, 1341–1351. [Google Scholar] [CrossRef]
- Cosenza-Nashat, M.A.; Bauman, A.; Zhao, M.-L.; Morgello, S.; Suh, H.-S.; Lee, S.C. Cannabinoid receptor expression in HIV encephalitis and HIV-associated neuropathologic comorbidities. Neuropathol. Appl. Neurobiol. 2011, 37, 464–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, C.; Kim, W.K.; Chavarría, I.; Hillard, C.J.; Mackie, K.; Tolón, R.M.; Williams, K.; Romero, J. A glial endogenous cannabinoid system is upregulated in the brains of macaques with simian immunodeficiency vi-rus-induced encephalitis. J. Neurosci. 2005, 25, 2530–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winsauer, P.J.; Molina, P.E.; Amedee, A.M.; Filipeanu, C.M.; McGoey, R.R.; Troxclair, D.A.; Walker, E.M.; Birke, L.L.; Stouwe, C.V.; Howard, J.M.; et al. Tolerance to chronic delta-9-tetrahydrocannabinol (Delta(9)-THC) in rhesus macaques infected with simian im-munodeficiency virus. Exp. Clin. Psychopharmacol. 2011, 19, 154–172. [Google Scholar] [CrossRef] [Green Version]
- Bredt, B.M.; Higuera-Alhino, D.; Shade, S.B.; Hebert, S.J.; McCune, J.M.; Abrams, D.I. Short-Term Effects of Cannabinoids on Immune Phenotype and Function in HIV-1-Infected Patients. J. Clin. Pharmacol. 2002, 42, 82S–89S. [Google Scholar] [CrossRef]
- Chaillon, A.; Nakazawa, M.; Anderson, C.; Christensen-Quick, A.; Ellis, R.J.; Franklin, D.; Morris, S.R.; Gianella, S. Effect of Cannabis Use on Human Immunodeficiency Virus DNA During Suppressive Antiretroviral Therapy. Clin. Infect. Dis. 2020, 70, 140–143. [Google Scholar] [CrossRef]
- Milloy, M.-J.; Marshall, B.; Kerr, T.; Richardson, L.; Hogg, R.; Guillemi, S.; Montaner, J.S.G.; Wood, E. High-intensity cannabis use associated with lower plasma human immunodeficiency virus-1 RNA viral load among recently infected people who use injection drugs. Drug Alcohol Rev. 2015, 34, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slawek, D.E.; Arnsten, J.; Sohler, N.; Zhang, C.; Grossberg, R.; Stein, M.; Cunningham, C.O. Daily and near-daily cannabis use is associated with HIV viral load suppression in people living with HIV who use cocaine. AIDS Care 2020, 1–8. [Google Scholar] [CrossRef]
- Amedee, A.M.; Nichols, W.A.; Le Capitaine, N.J.; Stouwe, C.V.; Birke, L.L.; Lacour, N.; Winsauer, P.J.; Molina, P.E. Chronic Δ9-Tetrahydrocannabinol Administration May Not Attenuate Simian Immunodeficiency Virus Disease Progression in Female Rhesus Macaques. AIDS Res. Hum. Retrovir. 2014, 30, 1216–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, P.E.; Winsauer, P.; Zhang, P.; Walker, E.; Birke, L.; Amedee, A.; Stouwe, C.V.; Troxclair, D.; McGoey, R.; Varner, K.; et al. Cannabinoid Administration Attenuates the Progression of Simian Immunodeficiency Virus. AIDS Res. Hum. Retrovir. 2011, 27, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Ghosn, J.; Leruez-Ville, M.; Blanche, J.; Delobelle, A.; Beaudoux, C.; Mascard, L.; Lecuyer, H.; Canestri, A.; Landman, R.; Zucman, D.; et al. HIV-1 DNA Levels in Peripheral Blood Mononuclear Cells and Cannabis Use are Associated with Intermittent HIV Shedding in Semen of Men Who Have Sex with Men on Successful Antiretroviral Regimens. Clin. Infect. Dis. 2014, 58, 1763–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.T.; Saag, L.A.; Kipp, A.M.; Logan, J.; Shepherd, B.E.; Koethe, J.R.; Turner, M.; Bebawy, S.; Sterling, T.R.; Hulgan, T. Self-reported Cannabis Use and Changes in Body Mass Index, CD4 T-Cell Counts, and HIV-1 RNA Suppression in Treated Persons with HIV. AIDS Behav. 2019, 24, 1275–1280. [Google Scholar] [CrossRef]
- Kipp, A.M.; Rebeiro, P.F.; Shepherd, B.E.; Brinkley-Rubinstein, L.; Turner, M.; Bebawy, S.; Sterling, T.R.; Hulgan, T. Daily Marijuana Use is Associated with Missed Clinic Appointments Among HIV-Infected Persons Engaged in HIV Care. AIDS Behav. 2017, 21, 1996–2004. [Google Scholar] [CrossRef]
- Bonn-Miller, M.O.; Oser, M.L.; Bucossi, M.M.; Trafton, J.A. Cannabis use and HIV antiretroviral therapy adherence and HIV-related symptoms. J. Behav. Med. 2014, 37, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Slawson, G.; Milloy, M.-J.; Balneaves, L.; Simo, A.; Guillemi, S.; Hogg, R.; Montaner, J.; Wood, E.; Kerr, T. High-Intensity Cannabis Use and Adherence to Antiretroviral Therapy Among People Who Use Illicit Drugs in a Canadian Setting. AIDS Behav. 2015, 19, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Rosen, M.I.; Black, A.C.; Arnsten, J.H.; Goggin, K.; Remien, R.H.; Simoni, J.M.; Golin, C.E.; Bangsberg, D.R.; Liu, H. Association Between Use of Specific Drugs and Antiretroviral Adherence: Findings from MACH 14. AIDS Behav. 2013, 17, 142–147. [Google Scholar] [CrossRef]
- Paschen-Wolff, M.M.; Campbell, A.N.; Tross, S.; Choo, T.-H.; Pavlicova, M.; Jarlais, D.D. DSM-5 substance use disorder symptom clusters and HIV antiretroviral therapy (ART) adherence. AIDS Care 2020, 32, 645–650. [Google Scholar] [CrossRef]
- Peretti-Watel, P.; Spire, B.; Lert, F.; Obadia, Y. Drug use patterns and adherence to treatment among HIV-positive patients: Evidence from a large sample of French outpatients (ANRS-EN12-VESPA 2003). Drug Alcohol Depend. 2006, 82, S71–S79. [Google Scholar] [CrossRef]
- Lake, S.; Kerr, T.; Capler, R.; Shoveller, J.; Montaner, J.; Milloy, M.-J. High-intensity cannabis use and HIV clinical outcomes among HIV-positive people who use illicit drugs in Vancouver, Canada. Int. J. Drug Policy 2017, 42, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Vidot, D.C.; Lerner, B.; Gonzalez, R. Cannabis Use, Medication Management and Adherence Among Persons Living with HIV. AIDS Behav. 2017, 21, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Socias, M.E.; Milloy, M.-J. Substance Use and Adherence to Antiretroviral Therapy: What Is Known and What Is Unknown. Curr. Infect. Dis. Rep. 2018, 20, 36. [Google Scholar] [CrossRef]
- Stout, S.M.; Cimino, N.M. Exogenous cannabinoids as substrates, inhibitors, and inducers of human drug metabolizing enzymes: A systematic review. Drug Metab. Rev. 2014, 46, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosel, B.W.; Aweeka, F.T.; Benowitz, N.L.; Shade, S.B.; Hilton, J.F.; Lizak, P.S.; Abrams, D.I. The effects of cannabinoids on the pharmacokinetics of indinavir and nelfinavir. AIDS 2002, 16, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Keen, L.; Abbate, A.; Blanden, G.; Priddie, C.; Moeller, F.G.; Rathore, M. Confirmed marijuana use and lymphocyte count in black people living with HIV. Drug Alcohol Depend. 2019, 198, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.D.; Crawford, R.B.; Henriquez, J.E.; Aldhamen, Y.A.; Gulick, P.; Amalfitano, A.; Kaminski, N.E. HIV-infected cannabis users have lower circulating CD16+ monocytes and IP-10 levels compared to non-using HIV patients. Aids 2017, 32, 419–429. [Google Scholar] [CrossRef]
- Manuzak, J.A.; Gott, T.M.; Kirkwood, J.S.; Coronado, E.; Hensley-McBain, T.; Miller, C.; Cheu, R.K.; Collier, A.C.; Funderburg, N.T.; Martin, J.N.; et al. Heavy Cannabis Use Associated with Reduction in Activated and Inflammatory Immune Cell Frequencies in Antiretroviral Therapy–Treated Human Immunodeficiency Virus–Infected Individuals. Clin. Infect. Dis. 2018, 66, 1872–1882. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.J.; Peterson, S.N.; Li, Y.; Schrier, R.; Iudicello, J.; Letendre, S.; Morgan, E.; Tang, B.; Grant, I.; Cherner, M. Recent cannabis use in HIV is associated with reduced inflammatory markers in CSF and blood. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e809. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.D.O.F.D.; Silva, J.M.; Dorneles, G.P.; Barros, J.; Ribeiro, C.B.; Noronha, I.; Barbosa, G.R.; Souza, L.; Guilarde, A.O.; Pereira, A.; et al. Distinct inflammatory profiles in HIV-infected individuals under antiretroviral therapy using cannabis, cocaine or cannabis plus cocaine. AIDS 2019, 33, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Vidot, D.C.; Manuzak, J.; Klatt, N.R.; Pallikkuth, S.; Roach, M.; Dilworth, S.E.; Pahwa, S.; Carrico, A.W. Brief Report: Hazardous Cannabis Use and Monocyte Activation Among Methamphetamine Users with Treated HIV Infection. J. Acquir. Immune Defic. Syndr. 2019, 81, 361–364. [Google Scholar] [CrossRef]
- Santos, M.E.; Protopopescu, C.; Sogni, P.; Yaya, I.; Piroth, L.; Bailly, F.; Marcellin, F.; Esterle, L.; Wittkop, L.; Rosenthal, E.; et al. HCV-Related Mortality Among HIV/HCV Co-infected Patients: The Importance of Behaviors in the HCV Cure Era (ANRS CO13 HEPAVIH Cohort). AIDS Behav. 2019, 24, 1069–1084. [Google Scholar] [CrossRef]
- Nordmann, S.; Vilotitch, A.; Roux, P.; Esterle, L.; Spire, B.; Marcellin, F.; Salmon-Ceron, D.; Dabis, F.; Chas, J.; Rey, D.; et al. Daily cannabis and reduced risk of steatosis in human immunodeficiency virus and hepatitis C virus-co-infected patients (ANRS CO13-HEPAVIH). J. Viral Hepat. 2017, 25, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, M.P.; Serfaty, L.; Vilotitch, A.; Winnock, M.; Poizot-Martin, I.; Loko, M.-A.; Lions, C.; Lascoux-Combe, C.; Roux, P.; Salmon-Ceron, D.; et al. Cannabis Use and Reduced Risk of Insulin Resistance in HIV-HCV Infected Patients: A Longitudinal Analysis (ANRS CO13 HEPAVIH). Clin. Infect. Dis. 2015, 61, 40–48. [Google Scholar] [CrossRef]
- Brunet, L.; Moodie, E.E.M.; Rollet, K.; Cooper, C.; Walmsley, S.; Potter, M.; Klein, M.B.; for the Canadian Co-infection Cohort Investigators. Marijuana Smoking Does Not Accelerate Progression of Liver Disease in HIV–Hepatitis C Coinfection: A Longitudinal Cohort Analysis. Clin. Infect. Dis. 2013, 57, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Marcellin, F.; Miailhes, P.; Santos, M.; Mercié, P.; Di Beo, V.; Salmon-Céron, D.; Barré, T.; Wittkop, L.; Protopopescu, C.; Zucman, D.; et al. Cannabis use and plasma HIV RNA levels in HIV-HCV co-infected patients receiving antiretroviral therapy: Data from the ANRS CO13 HEPAVIH cohort. Clin. Infect. Dis. 2020, 71, 2536–2538. [Google Scholar] [PubMed]
- Marcellin, F.; Lions, C.; Rosenthal, E.; Roux, P.; Sogni, P.; Wittkop, L.; Protopopescu, C.; Spire, B.; Salmon-Céron, M.; Dabis, F.; et al. No significant effect of cannabis use on the count and percentage of circulating CD4 T-cells in HIV-HCV co-infected patients (ANRS CO13-HEPAVIH French cohort). Drug Alcohol Rev. 2016, 36, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Sagnelli, C.; Uberti-Foppa, C.; Hasson, H.; Bellini, G.; Minichini, C.; Salpietro, S.; Messina, E.; Barbanotti, D.; Merli, M.; Punzo, F.; et al. Cannabinoid receptor 2-63 RR variant is independently associated with severe necroinflammation in HIV/HCV coinfected patients. PLoS ONE 2017, 12, e0181890. [Google Scholar] [CrossRef] [Green Version]
- Sagnelli, C.; Uberti-Foppa, C.; Hasson, H.; Bellini, G.; Minichini, C.; Salpietro, S.; Messina, E.; Barbanotti, D.; Merli, M.; Punzo, F.; et al. In vivo evidence that the cannabinoid receptor 2–63 RR variant is associated with the acquisition and/or expansion of HIV infection. HIV Med. 2018, 19, 597–604. [Google Scholar] [CrossRef]
- Lorkiewicz, S.A.; Ventura, A.S.; Heeren, T.C.; Winter, M.R.; Walley, A.Y.; Sullivan, M.; Samet, J.; Saitz, R. Lifetime marijuana and alcohol use, and cognitive dysfunction in people with human immunodeficiency virus infection. Subst. Abus. 2018, 39, 116–123. [Google Scholar] [CrossRef]
- Skalski, L.M.; Towe, S.L.; Sikkema, K.J.; Meade, C.S. Memory Impairment in HIV-Infected Individuals with Early and Late Initiation of Regular Marijuana Use. AIDS Behav. 2017, 22, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Schouten, J.; Su, T.; Wit, F.W.; Kootstra, N.A.; Caan, M.W.; Geurtsen, G.; Schmand, B.A.; Stolte, I.G.; Prins, M.; Majoie, C.B.; et al. Determinants of reduced cognitive performance in HIV-1-infected middle-aged men on combination antiretroviral therapy. AIDS 2016, 30, 1027–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crook, C.L.; Savin, M.J.; Byrd, D.; Summers, A.C.; Guzman, V.A.; Morris, E.P.; Tureson, K.; Aghvinian, M.; Cham, H.; Mindt, M.R. The neurocognitive effects of a past cannabis use disorder in a diverse sample of people living with HIV. AIDS Care 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.W.-M.; Paolillo, E.W.; Morgan, E.E.; Umlauf, A.; Sundermann, E.E.; Ellis, R.J.; Letendre, S.; Marcotte, T.D.; Heaton, R.K.; Grant, I. Cannabis Exposure is Associated with a Lower Likelihood of Neurocognitive Impairment in People Living With HIV. JAIDS J. Acquir. Immune Defic. Syndr. 2020, 83, 56–64. [Google Scholar] [CrossRef]
- Saloner, R.; Campbell, L.M.; Serrano, V.; Montoya, J.L.; Pasipanodya, E.; Paolillo, E.W.; Franklin, D.; Ellis, R.J.; Letendre, S.L.; Collier, A.C.; et al. Neurocognitive SuperAging in Older Adults Living With HIV: Demographic, Neuromedical and Everyday Functioning Correlates. J. Int. Neuropsychol. Soc. 2019, 25, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.A.; Lalee, Z.; Bell, R.P.; Towe, S.L.; Meade, C.S. Synergistic effects of HIV and marijuana use on functional brain network organization. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 104, 110040. [Google Scholar] [CrossRef]
- Wang, H.A.; Liang, H.-J.; Ernst, T.M.; Oishi, K.; Chang, L. Microstructural brain abnormalities in HIV+ individuals with or without chronic marijuana use. J. Neuroinflammation 2020, 17, 1–15. [Google Scholar] [CrossRef]
- Fennema-Notestine, C.; Ellis, R.J.; Archibald, S.L.; Jernigan, T.L.; Letendre, S.L.; Notestine, R.J.; Taylor, M.J.; Theilmann, R.J.; Julaton, M.D.; Croteau, D.J.; et al. Increases in brain white matter abnormalities and subcortical gray matter are linked to CD4 recovery in HIV infection. J. NeuroVirol. 2013, 19, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, L.H.; Sacktor, N.; Creighton, J.; Du, Y.; Endres, C.J.; Pomper, M.G.; Coughlin, J.M. Microglial activation is inversely associated with cognition in individuals living with HIV on effective antiretroviral therapy. AIDS 2018, 32, 1661–1667. [Google Scholar] [CrossRef]
- Thompson, K.A.; Cherry, C.L.; Bell, J.E.; McLean, C.A. Brain cell reservoirs of latent virus in presymptomatic HIV-infected individuals. Am. J. Pathol. 2011, 179, 1623–1629. [Google Scholar] [CrossRef] [Green Version]
- Teo, I.; Veryard, C.; Barnes, H.; An, S.F.; Jones, M.; Lantos, P.L.; Luthert, P.; Shaunak, S. Circular forms of unintegrated human immunodeficiency virus type 1 DNA and high levels of viral protein expression: Association with dementia and multinucleated giant cells in the brains of patients with AIDS. J. Virol. 1997, 71, 2928–2933. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; Song, K.; Vande Stouwe, C.; Hollenbach, A.; Amedee, A.; Mohan, M.; Winsauer, P.; Molina, P. Delta9-Tetrahydrocannabinol (Delta9-THC) Promotes Neuroimmune-Modulatory MicroRNA Profile in Striatum of Simian Immunodeficiency Virus (SIV)-Infected Macaques. J. Neuroimmune Pharmacol. 2016, 11, 192–213. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.J.; Peterson, S.; Cherner, M.; Morgan, E.; Schrier, R.; Tang, B.; Hoenigl, M.; Letendre, S.; Iudicello, J. Beneficial Effects of Cannabis on Blood Brain Barrier Function in HIV. Clin. Infect Dis. 2020. [Google Scholar] [CrossRef]
- Marsicano, G.; Lutz, B. Neuromodulatory functions of the endocannabinoid system. J. Endocrinol. Investig. 2006, 29 (Suppl. 3), 27–46. [Google Scholar]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef]
- Pryce, G.; Giovannoni, G.; Baker, D. Mifepristone or inhibition of 11beta-hydroxylase activity potentiates the sedating effects of the cannabinoid receptor-1 agonist Delta (9)-tetrahydrocannabinol in mice. Neurosci. Lett. 2003, 341, 164–166. [Google Scholar] [CrossRef]
- Klein, T.W. Cannabinoid-based drugs as anti-inflammatory therapeutics. Nat. Rev. Immunol. 2005, 5, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Bilkei-Gorzo, A. The endocannabinoid system in normal and pathological brain ageing. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3326–3341. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Lutz, B. Expression of the cannabinoid receptor CB1 in distinct neuronal subpopulations in the adult mouse forebrain. Eur. J. Neurosci. 1999, 11, 4213–4225. [Google Scholar] [CrossRef] [PubMed]
- Bedse, G.; Romano, A.; Cianci, S.; Lavecchia, A.M.; Lorenzo, P.; Elphick, M.R.; Laferla, F.M.; Vendemiale, G.; Grillo, C.; Altieri, F.; et al. Altered expression of the CB1 cannabinoid receptor in the triple transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2014, 40, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Silverdale, M.A.; McGuire, S.; McInnes, A.; Crossman, A.R.; Brotchie, J.M. Striatal cannabinoid CB1 receptor mRNA expression is decreased in the reserpine-treated rat model of Parkinson’s disease. Exp. Neurol. 2001, 169, 400–406. [Google Scholar] [CrossRef]
- Esposito, G.; Ligresti, A.; Izzo, A.; Bisogno, T.; Ruvo, M.; Di Rosa, M.; Di Marzo, V.; Iuvone, T. The Endocannabinoid System Protects Rat Glioma Cells Against HIV-1 Tat Protein-induced Cytotoxicity. J. Biol. Chem. 2002, 277, 50348–50354. [Google Scholar] [CrossRef] [Green Version]
- Roloff, A.M.; Thayer, S.A. Modulation of excitatory synaptic transmission by Delta 9-tetrahydrocannabinol switches from agonist to antagonist depending on firing rate. Mol. Pharmacol. 2009, 75, 892–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brailoiu, G.C.; Brailoiu, E.; Chang, J.K.; Dun, N.J. Excitatory effects of human immunodeficiency virus 1 Tat on cultured rat cerebral cortical neurons. Neurosciences 2008, 151, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Green, M.V.; Thayer, S.A. NMDARs Adapt to Neurotoxic HIV Protein Tat Downstream of a GluN2A–Ubiquitin Ligase Signaling Pathway. J. Neurosci. 2016, 36, 12640–12649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musante, V.; Summa, M.; Neri, E.; Puliti, A.; Godowicz, T.T.; Severi, P.; Battaglia, G.; Raiteri, M.; Pittaluga, A. The HIV-1 Viral Protein Tat Increases Glutamate and Decreases GABA Exocytosis from Human and Mouse Neocortical Nerve Endings. Cereb. Cortex 2009, 20, 1974–1984. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Hermes, D.J.; Nwanguma, B.; Jacobs, I.R.; Mackie, K.; Mukhopadhyay, S.; Lichtman, A.H.; Ignatowska-Jankowska, B.; Fitting, S. Endocannabinoids exert CB1 receptor-mediated neuroprotective effects in models of neuronal damage induced by HIV-1 Tat protein. Mol. Cell Neurosci. 2017, 83, 92–102. [Google Scholar] [CrossRef]
- Eugenin, E.A.; King, J.E.; Nath, A.; Calderon, T.M.; Zukin, R.S.; Bennett, M.V.; Berman, J.W. HIV-tat induces formation of an LRP-PSD-95- NMDAR-nNOS complex that promotes apoptosis in neurons and astrocytes. Proc. Natl. Acad. Sci. USA 2007, 104, 3438–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, A.; Psooy, K.; Martin, C.; Knudsen, B.; Magnuson, D.S.; Haughey, N.; Geiger, J.D. Identification of a human immunodeficiency virus type 1 Tat epitope that is neuroexcitatory and neurotoxic. J. Virol. 1996, 70, 1475–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, M.; Garden, G.A.; Lipton, S.A. Pathways to neuronal injury and apoptosis in HIV-associated dementia. Nat. Cell Biol. 2001, 410, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Furlan, R.; De Chiara, V.; Muzio, L.; Musella, A.; Motta, C.; Studer, V.; Cavasinni, F.; Bernardi, G.; Martino, G.; et al. Cannabinoid CB1 receptors regulate neuronal TNF-α effects in experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2011, 25, 1242–1248. [Google Scholar] [CrossRef]
- Chiarlone, A.; Bellocchio, L.; Blázquez, C.; Resel, E.; Soria-Gómez, E.; Cannich, A.; Ferrero, J.J.; Sagredo, O.; Benito, C.; Romero, J.; et al. A restricted population of CB1 cannabinoid receptors with neuroprotective activity. Proc. Natl. Acad. Sci. USA 2014, 111, 8257–8262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermes, D.J.; Xu, C.; Poklis, J.L.; Niphakis, M.J.; Cravatt, B.F.; Mackie, K.; Lichtman, A.H.; Ignatowska-Jankowska, B.M.; Fitting, S. Neuroprotective effects of fatty acid amide hydrolase catabolic enzyme inhibition in a HIV-1 Tat model of neuroAIDS. Neuropharmacology 2018, 141, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Thayer, S.A. Monoacylglycerol lipase inhibitor JZL184 prevents HIV-1 gp120-induced synapse loss by altering endo-cannabinoid signaling. Neuropharmacology 2018, 128, 269–281. [Google Scholar] [CrossRef]
- Gorantla, S.; Makarov, E.; Finke-Dwyer, J.; Castanedo, A.; Holguin, A.; Gebhart, C.L.; Gendelman, H.E.; Poluektova, L. Links between Progressive HIV-1 Infection of Humanized Mice and Viral Neuropathogenesis. Am. J. Pathol. 2010, 177, 2938–2949. [Google Scholar] [CrossRef]
- Avraham, H.K.; Jiang, S.; Fu, Y.; Rockenstein, E.; Makriyannis, A.; Wood, J.; Wang, L.; Masliah, E.; Avraham, S. Impaired neurogenesis by HIV-1-Gp120 is rescued by genetic deletion of fatty acid amide hydrolase enzyme. Br. J. Pharmacol. 2015, 172, 4603–4614. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Hermes, D.J.; Mackie, K.; Lichtman, A.H.; Ignatowska-Jankowska, B.M.; Fitting, S. Cannabinoids Occlude the HIV-1 Tat-Induced Decrease in GABAergic Neurotransmission in Prefrontal Cortex Slices. J. Neuroimmune Pharmacol. 2016, 11, 316–331. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, C. Endocannabinoid 2-Arachidonoylglycerol Protects Neurons by Limiting COX-2 Elevation. J. Biol. Chem. 2008, 283, 22601–22611. [Google Scholar] [CrossRef] [Green Version]
- Di Marzo, V. Targeting the endocannabinoid system: To enhance or reduce? Nat. Rev. Drug Discov. 2008, 7, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Costantino, C.M.; Gupta, A.; Yewdall, A.W.; Dale, B.M.; Devi, L.A.; Chen, B.K. Cannabinoid Receptor 2-Mediated Attenuation of CXCR4-Tropic HIV Infection in Primary CD4+ T Cells. PLoS ONE 2012, 7, e33961. [Google Scholar] [CrossRef] [Green Version]
- Peterson, P.; Gekker, G.; Hu, S.; Cabral, G.; Lokensgard, J. Cannabinoids and morphine differentially affect HIV-1 expression in CD4(+) lymphocyte and microglial cell cultures. J. Neuroimmunol. 2004, 147, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Duffhues, G.; Calzado, M.A.; de Vinuesa, A.G.; Caballero, F.J.; Ech-Chahad, A.; Appendino, G.; Krohn, K.; Fiebich, B.L.; Muñoz, E. Denbinobin, a naturally occurring 1,4-phenanthrenequinone, inhibits HIV-1 replication through an NF-kappaB-dependent pathway. Biochem. Pharmacol. 2008, 76, 1240–1250. [Google Scholar] [CrossRef]
- Sancho, R.; de la Vega, L.; Macho, A.; Appendino, G.; Di Marzo, V.; Muñoz, E. Mechanisms of HIV-1 Inhibition by the Lipid MediatorN-Arachidonoyldopamine. J. Immunol. 2005, 175, 3990–3999. [Google Scholar] [CrossRef]
- Ghorpade, A.; Nukuna, A.; Che, M.; Haggerty, S.; Persidsky, Y.; Carter, E.; Carhart, L.; Shafer, L.; Gendelman, H.E. Human immunodeficiency virus neurotropism: An analysis of viral replication and cytopathicity for divergent strains in monocytes and microglia. J. Virol. 1998, 72, 3340–3350. [Google Scholar] [CrossRef] [Green Version]
- Ghorpade, A.; Xia, M.Q.; Hyman, B.T.; Persidsky, Y.; Nukuna, A.; Bock, P.; Che, M.; Limoges, J.; Gendelman, H.E.; Mackay, C.R. Role of the beta-chemokine receptors CCR3 and CCR5 in human immunodeficiency virus type 1 infection of monocytes and microglia. J. Virol. 1998, 72, 3351–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, S.H.; Reichenbach, N.L.; Fan, S.; Rom, S.; Merkel, S.F.; Wang, X.; Ho, W.-Z.; Persidsky, Y. Attenuation of HIV-1 replication in macrophages by cannabinoid receptor 2 agonists. J. Leukoc. Biol. 2013, 93, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Rock, R.B.; Gekker, G.; Hu, S.; Sheng, W.S.; Cabral, G.A.; Martin, B.R.; Peterson, P.K. WIN55,212-2-Mediated Inhibition of HIV-1 Expression in Microglial Cells: Involvement of Cannabinoid Receptors. J. Neuroimmune Pharmacol. 2006, 2, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Fur, G.; Casellas, P. Expression of Central and Peripheral Cannabinoid Receptors in Human Immune Tissues and Leukocyte Subpopulations. J. Biol. Inorg. Chem. 1995, 232, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Lim, S.; Ryu, J.; Lee, C.-W.; Kim, Y.; Kang, J.-H.; Kang, S.-S.; Ahn, Y.K.; Park, C.-S.; Kim, J.J. CB1 and CB2 cannabinoid receptors differentially regulate the production of reactive oxygen species by macrophages. Cardiovasc. Res. 2009, 84, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.C.; Appelberg, S.; Goldberger, B.A.; Klein, T.W.; Sleasman, J.W.; Goodenow, M.M. Delta (9)-Tetrahydrocannabinol treatment during human monocyte differentiation reduces macrophage susceptibility to HIV-1 infection. J. Neuroimmune Pharmacol. 2014, 9, 369–379. [Google Scholar] [CrossRef]
- Agudelo, M.; Figueroa, G.; Yndart, A.; Casteleiro, G.; Muñoz, K.; Samikkannu, T.; Atluri, V.; Nair, M.P. Alcohol and Cannabinoids Differentially Affect HIV Infection and Function of Human Monocyte-Derived Dendritic Cells (MDDC). Front. Microbiol. 2015, 6, 1452. [Google Scholar] [CrossRef]
- Chen, W.; Crawford, R.B.; Kaplan, B.L.; Kaminski, N.E. Modulation of HIVGP120 Antigen-Specific Immune Responses In Vivo by Delta9-Tetrahydrocannabinol. J. Neuroimmune Pharmacol. 2015, 10, 344–355. [Google Scholar] [CrossRef]
- Chen, W.; Kaplan, B.; Pike, S.T.; Topper, L.A.; Lichorobiec, N.R.; Simmons, S.; Ramabhadran, R.; Kaminski, N.E. Magnitude of stimulation dictates the cannabinoid-mediated differential T cell response to HIVgp120. J. Leukoc. Biol. 2012, 92, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Lombard, C.; Hegde, V.L.; Nagarkatti, M.; Nagarkatti, P.S. Perinatal exposure to Delta9-tetrahydrocannabinol triggers profound defects in T cell differentiation and function in fetal and postnatal stages of life, including decreased responsiveness to HIV antigens. J. Pharmacol. Exp. Ther. 2011, 339, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Roth, M.D.; Tashkin, D.P.; Whittaker, K.M.; Choi, R.; Baldwin, G.C. Tetrahydrocannabinol suppresses immune function and enhances HIV replication in the huPBL-SCID mouse. Life Sci. 2005, 77, 1711–1722. [Google Scholar] [CrossRef]
- Correa, F.; Docagne, F.; Mestre, L.; Clemente, D.; Hernangómez, M.; Loria, F.; Guaza, C. A role for CB2 receptors in anandamide signalling pathways involved in the regulation of IL-12 and IL-23 in microglial cells. Biochem. Pharmacol. 2009, 77, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, F.M.; Mestre, L.; Loría, F.; Spagnolo, A.; Docagne, F.; Di Marzo, V.; Guaza, C. Anandamide enhances IL-10 production in activated microglia by targeting CB (2) receptors: Roles of ERK1/2, JNK, and NF-kappaB. Glia 2010, 58, 135–147. [Google Scholar] [CrossRef]
- Gabrielli, M.; Battista, N.; Riganti, L.; Prada, I.; Antonucci, F.; Cantone, L.; Matteoli, M.; Maccarrone, M.; Verderio, C. Active endocannabinoids are secreted on extracellular membrane vesicles. EMBO Rep. 2015, 16, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer-Stenger, K.; Pettit, D.A.D.; Cabral, G.A. Delta 9-tetrahydrocannabinol inhibition of tumor necrosis factor-alpha: Sup-pression of post-translational events. J. Pharmacol. Exp. Ther. 1993, 267, 1558. [Google Scholar]
- Ehrhart, J.; Obregon, D.; Mori, T.; Hou, H.; Sun, N.; Bai, Y.; Klein, T.; Fernandez, F.; Tan, J.; Shytle, R.D. Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation. J. Neuroinflamm. 2005, 2, 29. [Google Scholar] [CrossRef] [Green Version]
- Romero-Sandoval, E.A.; Horvath, R.; Landry, R.P.; De Leo, J.A. Cannabinoid Receptor Type 2 Activation Induces a Microglial Anti-Inflammatory Phenotype and Reduces Migration via MKP Induction and ERK Dephosphorylation. Mol. Pain 2009, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Racz, I.; Nadal, X.; Alferink, J.; Baños, J.E.; Rehnelt, J.; Martín, M.; Pintado, B.; Gutierrez-Adan, A.; Sanguino, E.; Bellora, N.; et al. Interferon-gamma is a critical modulator of CB (2) cannabinoid receptor signaling during neuropathic pain. J. Neurosci. 2008, 28, 12136–12145. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.D.; Crawford, R.B.; Bach, A.; Sermet, S.; Amalfitano, A.; Kaminski, N.E. Delta (9)-Tetrahydrocannabinol Suppresses Monocyte-Mediated Astrocyte Production of Monocyte Chemoattractant Protein 1 and Interleukin-6 in a Toll-Like Receptor 7-Stimulated Human Coculture. J. Pharmacol. Exp. Ther. 2019, 371, 191–201. [Google Scholar] [CrossRef]
- Sheng, W.S.; Hu, S.; Ni, H.T.; Rock, R.B.; Peterson, P.K. WIN55,212-2 Inhibits Production of CX3CL1 by Human Astrocytes: Involvement of p38 MAP Kinase. J. Neuroimmune Pharmacol. 2009, 4, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Thangaraj, A.; Periyasamy, P.; Liao, K.; Bendi, V.S.; Callen, S.; Pendyala, G.; Buch, S. HIV-1 TAT-mediated microglial activation: Role of mitochondrial dysfunction and defective mitophagy. Autophagy 2018, 14, 1596–1619. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.; Hu, S.; Hegg, C.; Thayer, S.; Peterson, P. Activation of Human Microglial Cells by HIV-1 gp41 and Tat Proteins. Clin. Immunol. 2000, 96, 243–251. [Google Scholar] [CrossRef]
- Swinton, M.K.; Carson, A.; Telese, F.; Sanchez, A.B.; Soontornniyomkij, B.; Rad, L.; Batki, I.; Quintanilla, B.; Pérez-Santiago, J.; Achim, C.L.; et al. Mitochondrial biogenesis is altered in HIV+ brains exposed to ART: Implications for therapeutic targeting of astroglia. Neurobiol. Dis. 2019, 130, 104502. [Google Scholar] [CrossRef]
- Krishnan, G.; Chatterjee, N. Endocannabinoids affect innate immunity of Muller glia during HIV-1 Tat cytotoxicity. Mol. Cell. Neurosci. 2014, 59, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Raborn, E.S.; Marciano-Cabral, F.; Buckley, N.E.; Martin, B.R.; Cabral, G.A. The Cannabinoid Delta-9-tetrahydrocannabinol Mediates Inhibition of Macrophage Chemotaxis to RANTES/CCL5: Linkage to the CB2 Receptor. J. Neuroimmune Pharmacol. 2008, 3, 117–129. [Google Scholar] [CrossRef]
- Raborn, E.S.; Cabral, G.A. Cannabinoid Inhibition of Macrophage Migration to the Trans-Activating (Tat) Protein of HIV-1 Is Linked to the CB2 Cannabinoid Receptor. J. Pharmacol. Exp. Ther. 2010, 333, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Fraga, D.; Raborn, E.S.; Ferreira, G.A.; Cabral, G.A. Cannabinoids Inhibit Migration of Microglial-like Cells to the HIV Protein Tat. J. Neuroimmune Pharmacol. 2011, 6, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic Cannabinoid Receptors Regulate Microglial Cell Migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar] [CrossRef] [Green Version]
- Gorantla, S.; Makarov, E.; Roy, D.; Finke-Dwyer, J.; Murrin, L.C.; Gendelman, H.E.; Poluektova, L. Immunoregulation of a CB2 Receptor Agonist in a Murine Model of NeuroAIDS. J. Neuroimmune Pharmacol. 2010, 5, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.G.; Reinke, S.N.; Mamik, M.K.; McKenzie, B.; Maingat, F.; Branton, W.G.; Broadhurst, D.; Power, C. Rapid inflammasome activation in microglia contributes to brain disease in HIV/AIDS. Retrovirology 2014, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Gao, J.; Taxman, D.J.; Ting, J.P.; Su, L. HIV-1 infection induces interleukin-1beta production via TLR8 protein-dependent and NLRP3 inflammasome mechanisms in human monocytes. J. Biol. Chem. 2014, 289, 21716–21726. [Google Scholar] [CrossRef] [Green Version]
- Bandera, A.; Masetti, M.; Fabbiani, M.; Biasin, M.; Muscatello, A.; Squillace, N.; Clerici, M.; Gori, A.; Trabattoni, D. The NLRP3 Inflammasome Is Upregulated in HIV-Infected Antiretroviral Therapy-Treated Individuals with Defective Immune Recovery. Front. Immunol. 2018, 9, 214. [Google Scholar] [CrossRef] [Green Version]
- Mazaheri-Tehrani, E.; Mohraz, M.; Nasi, M.; Chester, J.; De Gaetano, A.; Lo Tartaro, D.; SeyedAlinaghi, S.; Gholami, M.; De Biasi, S.; Gibellini, L.; et al. NLRP3 and IL-1β Gene Expression Is Elevated in Monocytes From HIV-Treated Patients With Neu-rocognitive Disorders. J. Acquir. Immune Defic. Syndr. 2021, 86, 496–499. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yang, W.; Zeng, Z.; Wei, Y.; Gao, J.; Zhang, B.; Li, L.; Liu, L.; Wan, Y.; Zeng, Q.; et al. NLRP3-dependent pyroptosis is required for HIV-1 gp120-induced neuropathology. Cell. Mol. Immunol. 2020, 17, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef]
- Rawat, P.; Teodorof-Diedrich, C.; Spector, S.A. Human immunodeficiency virus Type-1 single-stranded RNA activates the NLRP3 inflammasome and impairs autophagic clearance of damaged mitochondria in human microglia. Glia 2019, 67, 802–824. [Google Scholar] [CrossRef] [PubMed]
- Chivero, E.T.; Guo, M.-L.; Periyasamy, P.; Liao, K.; Callen, S.E.; Buch, S. HIV-1 Tat Primes and Activates Microglial NLRP3 Inflammasome-Mediated Neuroinflammation. J. Neurosci. 2017, 37, 3599–3609. [Google Scholar] [CrossRef] [Green Version]
- Mamik, M.K.; Hui, E.; Branton, W.G.; McKenzie, B.A.; Chisholm, J.; Cohen, E.A.; Power, C. HIV-1 Viral Protein R Activates NLRP3 Inflammasome in Microglia: Implications for HIV-1 Associated Neuroin-flammation. J. Neuroimmune Pharmacol. 2017, 12, 233–248. [Google Scholar] [CrossRef]
- Shao, B.-Z.; Wei, W.; Ke, P.; Xu, Z.-Q.; Zhou, J.-X.; Liu, C. Activating Cannabinoid Receptor 2 Alleviates Pathogenesis of Experimental Autoimmune Encephalomyelitis Via Activation of Autophagy and Inhibiting NLRP3 Inflammasome. CNS Neurosci. Ther. 2014, 20, 1021–1028. [Google Scholar] [CrossRef]
- Hu, S.; Sheng, W.S.; Rock, R.B. CB2 Receptor Agonists Protect Human Dopaminergic Neurons against Damage from HIV-1 gp120. PLoS ONE 2013, 8, e77577. [Google Scholar] [CrossRef] [PubMed]
- Avraham, H.K.; Jiang, S.; Fu, Y.; Rockenstein, E.; Makriyannis, A.; Zvonok, A.; Masliah, E.; Avraham, S. The cannabinoid CB2receptor agonist AM1241 enhances neurogenesis in GFAP/Gp120 transgenic mice displaying deficits in neurogenesis. Br. J. Pharmacol. 2014, 171, 468–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racz, I.; Nadal, X.; Alferink, J.; Baños, J.E.; Rehnelt, J.; Martín, M.; Pintado, B.; Gutierrez-Adan, A.; Sanguino, E.; Manzanares, J.; et al. Crucial Role of CB2 Cannabinoid Receptor in the Regulation of Central Immune Responses during Neuropathic Pain. J. Neurosci. 2008, 28, 12125–12135. [Google Scholar] [CrossRef] [Green Version]
- Wallace, V.C.J.; Blackbeard, J.; Segerdahl, A.R.; Hasnie, F.; Pheby, T.; McMahon, S.B.; Rice, A.S.C. Characterization of rodent models of HIV-gp120 and anti-retroviral-associated neuropathic pain. Brain 2007, 130, 2688–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, V.C.; Blackbeard, J.; Pheby, T.; Segerdahl, A.R.; Davies, M.; Hasnie, F.; Hall, S.; McMahon, S.B.; Rice, A.S. Pharmacological, behavioural and mechanistic analysis of HIV-1 gp120 induced painful neuropathy. Pain 2007, 133, 47–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, W.S.; Chauhan, P.; Hu, S.; Prasad, S.; Lokensgard, J.R. Antiallodynic Effects of Cannabinoid Receptor 2 (CB2R) Agonists on Retrovirus Infection-Induced Neuropathic Pain. Pain Res. Manag. 2019, 2019, 1260353-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, E.; Khajah, M.A.; Masocha, W. β-Caryophyllene, a CB2-Receptor-Selective Phytocannabinoid, Suppresses Mechanical Allodynia in a Mouse Model of Antiretroviral-Induced Neuropathic Pain. Molecules 2020, 25, 106. [Google Scholar] [CrossRef] [Green Version]
- Toborek, M.; Lee, Y.W.; Flora, G.; Pu, H.; András, I.E.; Wylegala, E.; Hennig, B.; Nath, A. Mechanisms of the blood-brain barrier disruption in HIV-1 infection. Cell Mol. Neurobiol. 2005, 25, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Persidsky, Y.; Ho, W.; Ramirez, S.H.; Potula, R.; Abood, M.E.; Unterwald, E.; Tuma, R. HIV-1 infection and alcohol abuse: Neurocognitive impairment, mechanisms of neurodegeneration and therapeutic interventions. Brain Behav. Immun. 2011, 25, S61–S70. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, A.; Duan, F.; Morsey, B.; Persidsky, Y.; Kanmogne, G.D. HIV-1 Activates Proinflammatory and Interferon-Inducible Genes in Human Brain Microvascular Endothelial Cells: Putative Mechanisms of Blood—Brain Barrier Dysfunction. Br. J. Pharmacol. 2007, 28, 697–711. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Yang, B.; Gendelman, H.E.; Persidsky, Y.; Kanmogne, G.D. STAT1 signaling modulates HIV-1–induced inflammatory responses and leukocyte transmigration across the blood-brain barrier. Blood 2008, 111, 2062–2072. [Google Scholar] [CrossRef] [PubMed]
- Persidsky, Y.; Ghorpade, A.; Rasmussen, J.; Limoges, J.; Liu, X.J.; Stins, M.; Fiala, M.; Way, D.; Kim, K.S.; Witte, M.H.; et al. Microglial and Astrocyte Chemokines Regulate Monocyte Migration through the Blood-Brain Barrier in Human Immunodeficiency Virus-1 Encephalitis. Am. J. Pathol. 1999, 155, 1599–1611. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, Y.; Wang, X.; Ye, L.; Zhou, Y.; Persidsky, Y.; Ho, W. Immune activation of human brain microvascular endothelial cells inhibits HIV replication in macrophages. Blood 2013, 121, 2934–2942. [Google Scholar] [CrossRef] [Green Version]
- Persidsky, Y.; Fan, S.; Dykstra, H.; Reichenbach, N.L.; Rom, S.; Ramirez, S.H. Activation of Cannabinoid Type Two Receptors (CB2) Diminish Inflammatory Responses in Macrophages and Brain Endothelium. J. Neuroimmune Pharmacol. 2015, 10, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Raborn, E.S.; Jamerson, M.; Marciano-Cabral, F.; Cabral, G.A. Cannabinoid inhibits HIV-1 Tat-stimulated adhesion of human monocyte-like cells to extracellular matrix proteins. Life Sci. 2014, 104, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Rom, S.; Zuluaga-Ramirez, V.; Dykstra, H.; Reichenbach, N.L.; Pacher, P.; Persidsky, Y. Selective Activation of Cannabinoid Receptor 2 in Leukocytes Suppresses Their Engagement of the Brain Endothelium and Protects the Blood-Brain Barrier. Am. J. Pathol. 2013, 183, 1548–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, S.H.; Haskó, J.; Skuba, A.; Fan, S.; Dykstra, H.; McCormick, R.; Reichenbach, N.; Krizbai, I.; Mahadevan, A.; Zhang, M.; et al. Activation of Cannabinoid Receptor 2 Attenuates Leukocyte-Endothelial Cell Interactions and Blood-Brain Barrier Dysfunction under Inflammatory Conditions. J. Neurosci. 2012, 32, 4004–4016. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.S.; Avraham, H.K.; Seng, S.; Tachado, S.D.; Koziel, H.; Makriyannis, A.; Avraham, S. Cannabinoids inhibit HIV-1 Gp120-mediated insults in brain microvascular endothelial cells. J. Immunol. 2008, 181, 6406–6416. [Google Scholar] [CrossRef] [PubMed]
- Pietr, M.; Kozela, E.; Levy, R.; Rimmerman, N.; Lin, Y.H.; Stella, N.; Vogel, Z.; Juknat, A. Differential changes in GPR55 during microglial cell activation. FEBS Lett. 2009, 583, 2071–2076. [Google Scholar] [CrossRef] [Green Version]
- Saliba, S.W.; Jauch, H.; Gargouri, B.; Keil, A.; Hurrle, T.; Volz, N.; Mohr, F.; Van Der Stelt, M.; Bräse, S.; Fiebich, B.L. Anti-neuroinflammatory effects of GPR55 antagonists in LPS-activated primary microglial cells. J. Neuroinflammation 2018, 15, 322. [Google Scholar] [CrossRef]
- Hill, J.D.; Zuluaga-Ramirez, V.; Gajghate, S.; Winfield, M.; Persidsky, Y. Activation of GPR55 increases neural stem cell proliferation and promotes early adult hippocampal neurogenesis. Br. J. Pharmacol. 2018, 175, 3407–3421. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.D.; Zuluaga-Ramirez, V.; Gajghate, S.; Winfield, M.; Sriram, U.; Rom, S.; Persidsky, Y. Activation of GPR55 induces neuroprotection of hippocampal neurogenesis and immune responses of neural stem cells following chronic, systemic inflammation. Brain Behav. Immun. 2019, 76, 165–181. [Google Scholar] [CrossRef]
- Reyes-Resina, I.; Navarro, G.; Aguinaga, D.; Canela, E.I.; Schoeder, C.T.; Załuski, M.; Kiec-Kononowicz, K.; Saura, C.A.; Müller, C.E.; Franco, R. Molecular and functional interaction between GPR18 and cannabinoid CB2 G-protein-coupled receptors. Relevance in neurodegenerative diseases. Biochem. Pharmacol. 2018, 157, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Munawar, N.; Oriowo, M.A.; Masocha, W. Antihyperalgesic Activities of Endocannabinoids in a Mouse Model of Antiretrovi-ral-Induced Neuropathic Pain. Front. Pharmacol. 2017, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Nasirinezhad, F.; Jergova, S.; Pearson, J.P.; Sagen, J. Attenuation of persistent pain-related behavior by fatty acid amide hydrolase (FAAH) inhibitors in a rat model of HIV sensory neuropathy. Neuropharmacology 2015, 95, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Yagyu, K.; Sackett, S.; Zhang, Y. Anti-Inflammatory Effects by Pharmacological Inhibition or Knockdown of Fatty Acid Amide Hydrolase in BV2 Microglial Cells. Cells 2019, 8, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccarrone, M.; Piccirilli, S.; Battista, N.; Del Duca, C.; Nappi, G.; Corasaniti, M.T.; Finazzi-Agrò, A.; Bagetta, G. Enhanced anandamide degradation is associated with neuronal apoptosis induced by the HIV-1 coat glycoprotein gp120 in the rat neocortex. J. Neurochem. 2004, 89, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Grabner, G.F.; Eichmann, T.O.; Wagner, B.; Gao, Y.; Farzi, A.; Taschler, U.; Radner, F.; Schweiger, M.; Lass, A.; Holzer, P.; et al. Deletion of Monoglyceride Lipase in Astrocytes Attenuates Lipopolysaccharide-induced Neuroinflammation. J. Biol. Chem. 2016, 291, 913–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojo-Bustamante, E.; Íñigo-Marco, I.; Abellanas, M.A.; Vinueza-Gavilanes, R.; Baltanás, A.; Luquin, E.; Arrasate, M.; Aymerich, M.S. CB2 Receptors and Neuron–Glia Interactions Modulate Neurotoxicity Generated by MAGL Inhibition. Biomoleciles 2020, 10, 1198. [Google Scholar] [CrossRef]
- Crews, L.; Lentz, M.R.; González, R.G.; Fox, H.S.; Masliah, E. Neuronal injury in simian immunodeficiency virus and other animal models of neuroAIDS. J. NeuroVirol. 2008, 14, 327–339. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engle, S.J.; Blaha, L.; Kleiman, R.J. Best Practices for Translational Disease Modeling Using Human iPSC-Derived Neurons. Neuron 2018, 100, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Ming, G.-L.; Brüstle, O.; Muotri, A.; Studer, L.; Wernig, M.; Christian, K.M. Cellular Reprogramming: Recent Advances in Modeling Neurological Diseases. J. Neurosci. 2011, 31, 16070–16075. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C.; et al. Brain-Region-Specific Organoids Using Mini-bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, K.-J.; Song, G.; Qian, X.; Pan, J.; Xu, D.; Rho, H.-S.; Kim, N.-S.; Habela, C.; Zheng, L.; Jacob, F.; et al. Zika-Virus-Encoded NS2A Disrupts Mammalian Cortical Neurogenesis by Degrading Adherens Junction Proteins. Cell Stem Cell 2017, 21, 349–358.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oddo, A.; Peng, B.; Tong, Z.; Wei, Y.; Tong, W.Y.; Thissen, H.; Voelcker, N.H. Advances in Microfluidic Blood–Brain Barrier (BBB) Models. Trends Biotechnol. 2019, 37, 1295–1314. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.L.; England, T.J.; O’Sullivan, S.E. A Novel Transwell Blood Brain Barrier Model Using Primary Human Cells. Front. Cell. Neurosci. 2019, 13, 230. [Google Scholar] [CrossRef] [Green Version]
- Yue, H.; Xie, K.; Ji, X.; Xu, B.; Wang, C.; Shi, P. Vascularized neural constructs for ex-vivo reconstitution of blood-brain barrier function. Biomaterials 2020, 245, 119980. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starr, A.; Jordan-Sciutto, K.L.; Mironets, E. Confound, Cause, or Cure: The Effect of Cannabinoids on HIV-Associated Neurological Sequelae. Viruses 2021, 13, 1242. https://doi.org/10.3390/v13071242
Starr A, Jordan-Sciutto KL, Mironets E. Confound, Cause, or Cure: The Effect of Cannabinoids on HIV-Associated Neurological Sequelae. Viruses. 2021; 13(7):1242. https://doi.org/10.3390/v13071242
Chicago/Turabian StyleStarr, Alexander, Kelly L. Jordan-Sciutto, and Eugene Mironets. 2021. "Confound, Cause, or Cure: The Effect of Cannabinoids on HIV-Associated Neurological Sequelae" Viruses 13, no. 7: 1242. https://doi.org/10.3390/v13071242