Olea Europaea Geminivirus: A Novel Bipartite Geminivirid Infecting Olive Trees

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.1.1. NGS Libraries and Identification of Gemini-Like Sequences
2.1.2. Occurrence of the Novel Virus in Olive Groves
2.1.3. Assessment of the Small RNA Profiles
2.2. Bioinformatics Analysis
2.3. Genome Organization and Homology Searches for OEGV Genes
2.4. Rolling Circle Amplifications and Southern Blot Assay
3. Results
3.1. High-Throughput Sequencing Results
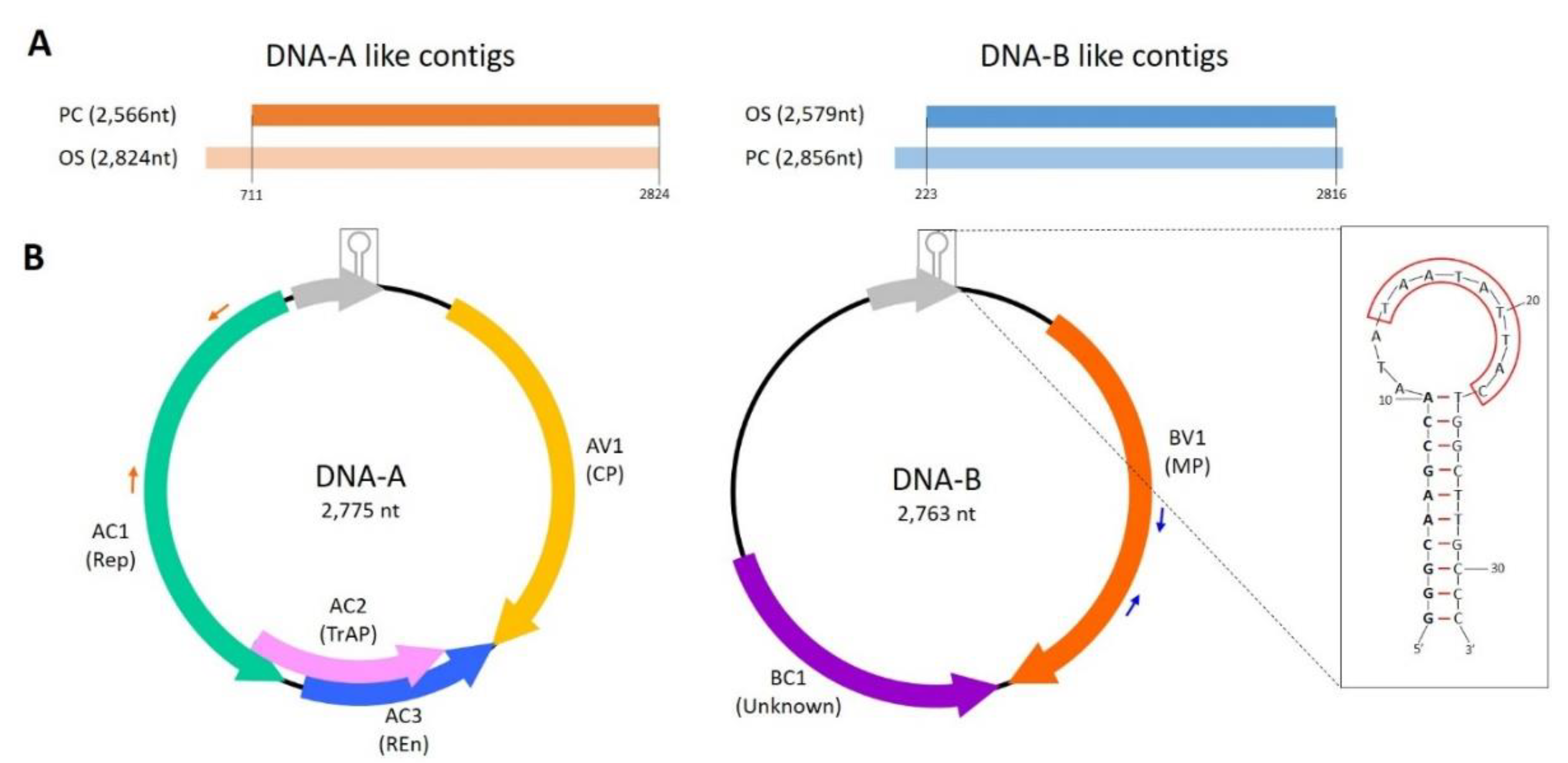
3.2. Genomic Features
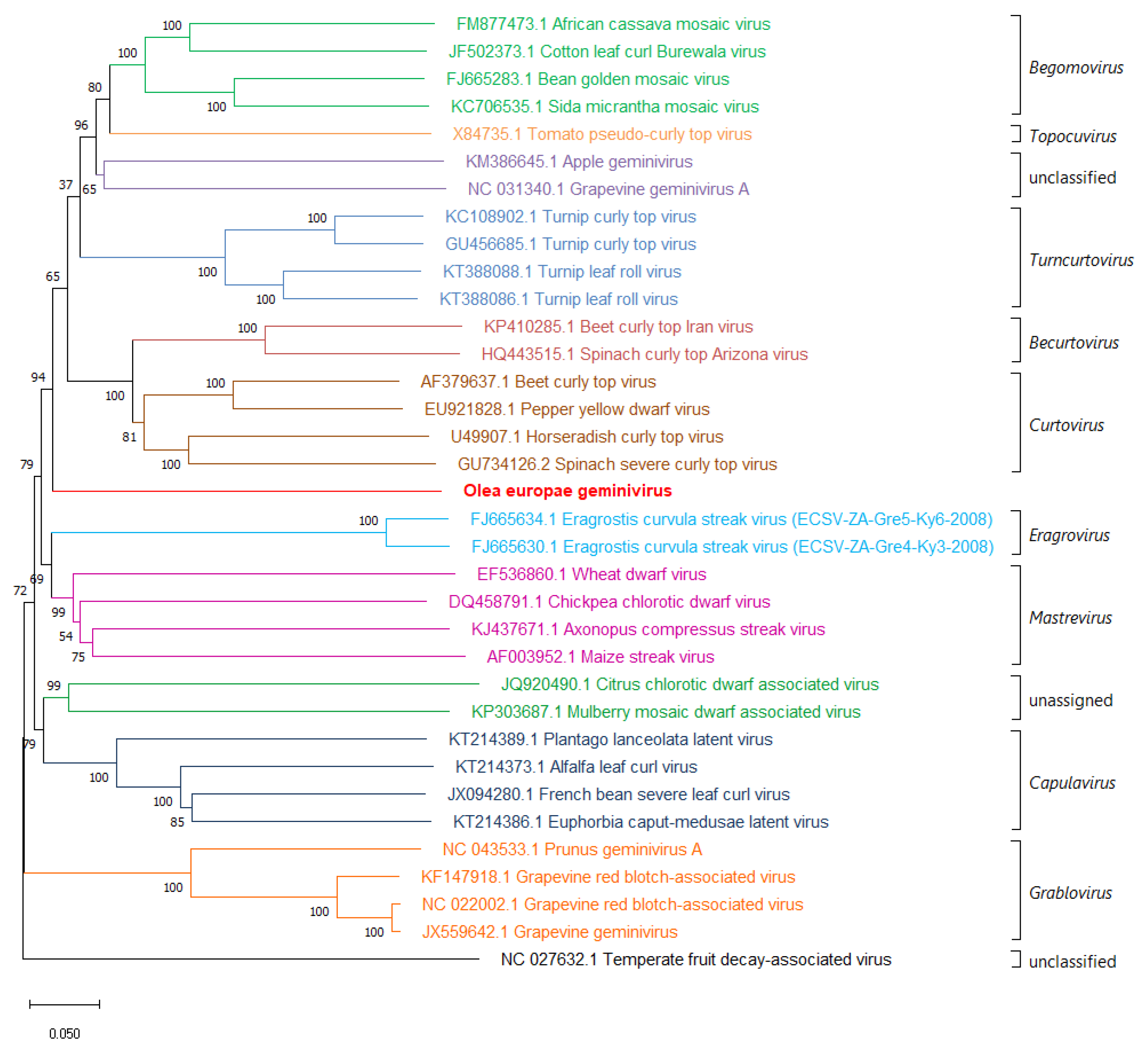
3.3. Phylogenetic Relationship with other Member of the Family Geminiviridae
3.4. A Survey for OEGV in Apulian Orchards Reveals This Virus Is Widespread
3.5. Attempts of Further Characterization by Southern Blot and Electron Microscopy
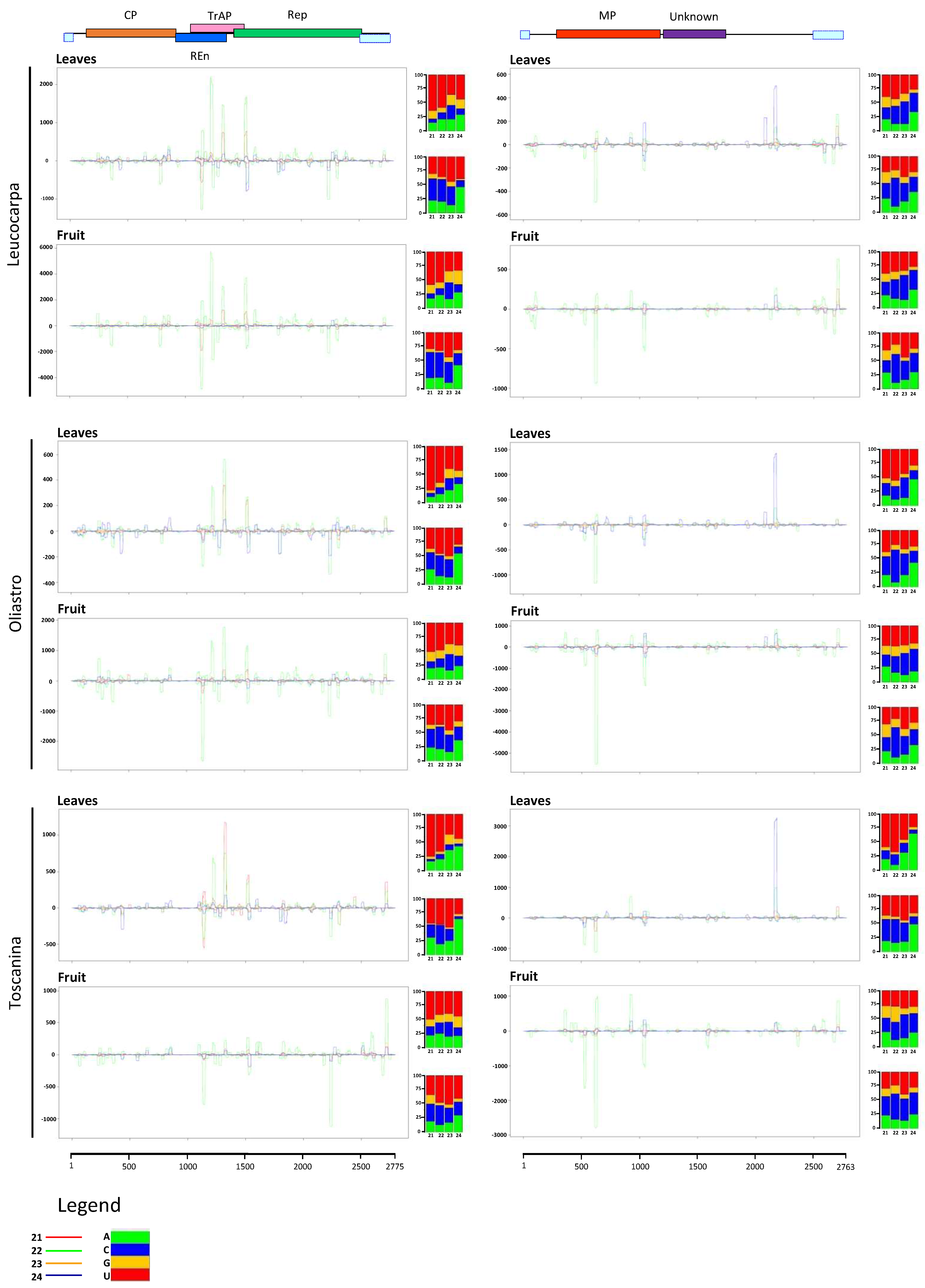
3.6. Analysis of the Virus-Derived sRNAs in OEGV-Infected Olive Trees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Çağlayan, K.; Faggioli, F.; Barba, M. Viruses, Phytoplasmas, and Diseases of Unknown Etiology of Olive Trees. Virus Virus-Like Dis. Pome Stone Fruits 2011, 289–297. [Google Scholar] [CrossRef]
- Martelli, G.P. Infectious Diseases and Certification of Olive: An Overview. EPPO Bull. 1999, 29, 127–133. [Google Scholar] [CrossRef]
- Felix, M.R.F.; Clara, M.I.E. Two necrovirus isolates with properties of olive latent virus 1 and of tobacco necrosis virus from olive in portugal. In Proceedings of the IV International Symposium on Olive Growing 586, Valenzano, Italy, 25–30 September 2000; pp. 725–728. [Google Scholar]
- Savino, V.; Barba, M.; Galitelli, G.; Martelli, G.P. Two Nepoviruses Isolated from Olive in Italy. Phytopathol. Mediterr. 1979, 18, 135–142. [Google Scholar]
- Savino, V.; Gallitelli, D. Isolation of Cucumber Mosaic Virus from Olive in Italy. Phytopathol. Mediterr. 1983, 22, 76–77. [Google Scholar]
- Faggioli, F.; Barba, M. An elongated virus isolated from olive (Olea Europea L.). In Proceedings of the XVI International Symposium on Fruit Tree Virus Diseases 386, Rome, Italy, 27 June–2 July 1994; pp. 593–600. [Google Scholar]
- Materazzi, A.; Toni, S.; Panettoni, A.; Osti, M.; Triolo, E. On the Presence of a New Isometric Virus in Olea europaea L. Proc. Convegno Annu. Soc. Ital. Patol. Veg. Udine 1996, 1, 57–59. [Google Scholar]
- Savino, V.; Sabanadzovic, S.; Scarito, G.; Laviola, C.; Martelli, G.P. Due Giallumi Dell’olivo Di Possibile Origine Virale in Sicilia. Inf. Fitopatol. 1996, 46, 55–59. [Google Scholar]
- Triolo, E.; Materazzi, A.; Toni, S. An Isolate of Tobacco Mosaic Tobamovirus from Olea europaea L. Adv. Hortic. Sci. 1996, 10, 39–45. [Google Scholar]
- Ferretti, L.; Faggioli, F.; Pasquini, G.; Sciarroni, R.; Pannelli, G.; Baldoni, L.; Barba, M. Strawberry Latent Ringspot Virus (SLRSV) Cause of Differentiation among Raggiola and Frantoio Olive Cultivars. J. Plant Pathol. 2002, 84, 182. [Google Scholar]
- Marte, M.; Gadani, F.; Savino, V.; Rugini, E. Strawberry Latent Ringspot Virus Associated with a New Disease of Olive in Central Italy. Plant Dis. 1986, 70, 171–172. [Google Scholar] [CrossRef]
- Ruiz-García, A.B.; Candresse, T.; Canales, C.; Morán, F.; Machado de Oliveira, C.; Bertolini, E.; Olmos, A. Molecular Characterization of the Complete Coding Sequence of Olive Leaf Yellowing-Associated Virus. Plants 2020, 9, 1272. [Google Scholar] [CrossRef]
- Radford, A.D.; Chapman, D.; Dixon, L.; Chantrey, J.; Darby, A.C.; Hall, N. Application of Next-Generation Sequencing Technologies in Virology. J. Gen. Virol. 2012, 93, 1853. [Google Scholar] [CrossRef] [Green Version]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent Advances on Detection and Characterization of Fruit Tree Viruses Using High-Throughput Sequencing Technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.T.; Voinnet, O.; Baulcombe, D.C. Initiation and Maintenance of Virus-Induced Gene Silencing. Plant Cell 1998, 10, 937–946. [Google Scholar] [CrossRef]
- Bologna, N.G.; Voinnet, O. The Diversity, Biogenesis, and Activities of Endogenous Silencing Small RNAs in Arabidopsis. Annu. Rev. Plant Biol. 2014, 65, 473–503. [Google Scholar] [CrossRef]
- Parent, J.-S.; Martinez de Alba, A.E.; Vaucheret, H. The Origin and Effect of Small RNA Signaling in Plants. Front. Plant Sci. 2012, 3, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumplin, N.; Voinnet, O. RNA Silencing Suppression by Plant Pathogens: Defence, Counter-Defence and Counter-Counter-Defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.; Vaucheret, H. Form, Function, and Regulation of ARGONAUTE Proteins. Plant Cell 2010, 22, 3879–3889. [Google Scholar] [CrossRef]
- Ruiz-Ferrer, V.; Voinnet, O. Roles of Plant Small RNAs in Biotic Stress Responses. Annu. Rev. Plant Biol. 2009, 60, 485–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varma, A.; Malathi, V.G. Emerging Geminivirus Problems: A Serious Threat to Crop Production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Scholthof, K.-B.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P. Top 10 Plant Viruses in Molecular Plant Pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef]
- Rybicki, E.P. A Top Ten List for Economically Important Plant Viruses. Arch. Virol. 2015, 160, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A.; Consortium, I.R. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131. [Google Scholar] [CrossRef]
- Loconsole, G.; Saldarelli, P.; Doddapaneni, H.; Savino, V.; Martelli, G.P.; Saponari, M. Identification of a Single-Stranded DNA Virus Associated with Citrus Chlorotic Dwarf Disease, a New Member in the Family Geminiviridae. Virology 2012, 432, 162–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Navarro, B.; Zhang, Z.; Lu, M.; Zhou, X.; Chi, S.; Di Serio, F.; Li, S. Identification and Molecular Characterization of a Novel Monopartite Geminivirus Associated with Mulberry Mosaic Dwarf Disease. J. Gen. Virol. 2015, 96, 2421–2434. [Google Scholar] [CrossRef]
- Liang, P.; Navarro, B.; Zhang, Z.; Wang, H.; Lu, M.; Xiao, H.; Wu, Q.; Zhou, X.; Di Serio, F.; Li, S. Identification and Characterization of a Novel Geminivirus with a Monopartite Genome Infecting Apple Trees. J. Gen. Virol. 2015, 96, 2411–2420. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Alabi, O.J.; Westrick, N.M.; Golino, D.; Rowhani, A. Description of a Novel Monopartite Geminivirus and Its Defective Subviral Genome in Grapevine. Phytopathology 2017, 107, 240–251. [Google Scholar] [CrossRef] [Green Version]
- Al Rwahnih, M.; Alabi, O.J.; Westrick, N.M.; Golino, D. Prunus Geminivirus A: A Novel Grablovirus Infecting Prunus Spp. Plant Dis. 2018, 102, 1246–1253. [Google Scholar] [CrossRef] [Green Version]
- Perry, K.L.; McLane, H.; Thompson, J.R.; Fuchs, M. A Novel Grablovirus from Non-Cultivated Grapevine (Vitis Sp.) in North America. Arch. Virol. 2018, 163, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Vaghi Medina, C.G.; Teppa, E.; Bornancini, V.A.; Flores, C.R.; Marino-Buslje, C.; López Lambertini, P.M. Tomato Apical Leaf Curl Virus: A Novel, Monopartite Geminivirus Detected in Tomatoes in Argentina. Front. Microbiol. 2018, 8, 2665. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A. Revision of Begomovirus Taxonomy Based on Pairwise Sequence Comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for Plant DNA Replication, Transcription, and Cell Cycle Regulation. Crit. Rev. Plant Sci. 1999, 18, 71–106. [Google Scholar] [CrossRef]
- Noueiry, A.O.; Lucas, W.J.; Gilbertson, R.L. Two Proteins of a Plant DNA Virus Coordinate Nuclear and Plasmodesmal Transport. Cell 1994, 76, 925–932. [Google Scholar] [CrossRef]
- Sanderfoot, A.A.; Lazarowitz, S.G. Getting It Together in Plant Virus Movement: Cooperative Interactions between Bipartite Geminivirus Movement Proteins. Trends Cell Biol. 1996, 6, 353–358. [Google Scholar] [CrossRef]
- Sánchez-Campos, S.; Martínez-Ayala, A.; Márquez-Martín, B.; Aragón-Caballero, L.; Navas-Castillo, J.; Moriones, E. Fulfilling Koch’s Postulates Confirms the Monopartite Nature of Tomato Leaf Deformation Virus: A Begomovirus Native to the New World. Virus Res. 2013, 173, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.G.; Thompson, W.F. Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giampetruzzi, A.; Chiumenti, M.; Minafra, A.; Saldarelli, P. Small RNA isolation from tissues of grapevine and woody plants. In Viral Metagenomics; Springer: New York, NY, USA, 2018; pp. 27–36. [Google Scholar]
- Hernandez, D.; François, P.; Farinelli, L.; Øster\aas, M.; Schrenzel, J. De Novo Bacterial Genome Sequencing: Millions of Very Short Reads Assembled on a Desktop Computer. Genome Res. 2008, 18, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prüfer, K.; Stenzel, U.; Dannemann, M.; Green, R.E.; Lachmann, M.; Kelso, J. PatMaN: Rapid Alignment of Short Sequences to Large Databases. Bioinformatics 2008, 24, 1530–1531. [Google Scholar] [CrossRef]
- Zuker, M. Mfold Web Server for Nucleic Acid Folding and Hybridization Prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Seguin, J.; Otten, P.; Baerlocher, L.; Farinelli, L.; Pooggin, M.M. MISIS: A Bioinformatics Tool to View and Analyze Maps of Small RNAs Derived from Viruses and Genomic Loci Generating Multiple Small RNAs. J. Virol. Methods 2014, 195, 120–122. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Letunic, I.; Bork, P. 20 Years of the SMART Protein Domain Annotation Resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I. CDD: NCBI’s Conserved Domain Database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A Virus Classification Tool Based on Pairwise Sequence Alignment and Identity Calculation. PLoS ONE 2014, 9, e108277. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Idris, A.M.; Mansoor, S.; Bedford, I.D.; Dhawan, P.; Rishi, N.; Siwatch, S.S.; Abdel-Salam, A.M. Diversity of DNA β, a Satellite Molecule Associated with Some Monopartite Begomoviruses. Virology 2003, 312, 106–121. [Google Scholar] [CrossRef] [Green Version]
- Fontes, E.P.; Eagle, P.A.; Sipe, P.S.; Luckow, V.A.; Hanley-Bowdoin, L. Interaction between a Geminivirus Replication Protein and Origin DNA Is Essential for Viral Replication. J. Biol. Chem. 1994, 269, 8459–8465. [Google Scholar] [CrossRef]
- Laufs, J.; Traut, W.; Heyraud, F.; Matzeit, V.; Rogers, S.G.; Schell, J.; Gronenborn, B. In Vitro Cleavage and Joining at the Viral Origin of Replication by the Replication Initiator Protein of Tomato Yellow Leaf Curl Virus. Proc. Natl. Acad. Sci. USA 1995, 92, 3879–3883. [Google Scholar] [CrossRef] [Green Version]
- Blevins, T.; Rajeswaran, R.; Shivaprasad, P.V.; Beknazariants, D.; Si-Ammour, A.; Park, H.-S.; Vazquez, F.; Robertson, D.; Meins Jr, F.; Hohn, T. Four Plant Dicers Mediate Viral Small RNA Biogenesis and DNA Virus Induced Silencing. Nucleic Acids Res. 2006, 34, 6233–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wang, Y.; Guo, W.; Xie, Y.; Xie, Q.; Fan, L.; Zhou, X. Characterization of Small Interfering RNAs Derived from the Geminivirus/Betasatellite Complex Using Deep Sequencing. PLoS ONE 2011, 6, e16928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingo, E.; Sheldon, J.; Perales, C. Viral Quasispecies Evolution. Microbiol. Mol. Biol. Rev. 2012, 76, 159–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andino, R.; Domingo, E. Viral Quasispecies. Virology 2015, 479, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rwahnih, M.A.; Dave, A.; Anderson, M.M.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Association of a DNA Virus with Grapevines Affected by Red Blotch Disease in California. Phytopathology 2013, 103, 1069–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenz, B.; Thompson, J.R.; Fuchs, M.; Perry, K.L. Complete Genome Sequence of a New Circular DNA Virus from Grapevine. Am. Soc. Microbiol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Polston, J.E.; Londoño, M.A.; Capobianco, H. The Complete Genome Sequence of New World Jatropha Mosaic Virus. Arch. Virol. 2014, 159, 3131–3136. [Google Scholar] [CrossRef]
- Basso, M.F.; da Silva, J.C.F.; Fajardo, T.V.M.; Fontes, E.P.B.; Zerbini, F.M. A Novel, Highly Divergent SsDNA Virus Identified in Brazil Infecting Apple, Pear and Grapevine. Virus Res. 2015, 210, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Marwal, A.; Sahu, A.K.; Gaur, R.K. First Report of Airborne Begomovirus Infection in Melia Azedarach (Pride of India), an Ornamental Tree in India. Aerobiologia 2014, 30, 211–215. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at Redirecting and Reprogramming Plant Processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Amiot, M.J.; Fleuriet, A.; Macheix, J.J. Importance and Evolution of Phenolic Compounds in Olive during Growth and Maturation. J. Agric. Food Chem. 1986, 34, 823–826. [Google Scholar] [CrossRef]
- Filloux, D.; Murrell, S.; Koohapitagtam, M.; Golden, M.; Julian, C.; Galzi, S.; Uzest, M.; Rodier-Goud, M.; D’Hont, A.; Vernerey, M.S. The Genomes of Many Yam Species Contain Transcriptionally Active Endogenous Geminiviral Sequences That May Be Functionally Expressed. Virus Evol. 2015, 1, vev002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejarano, E.R.; Khashoggi, A.; Witty, M.; Lichtenstein, C. Integration of Multiple Repeats of Geminiviral DNA into the Nuclear Genome of Tobacco during Evolution. Proc. Natl. Acad. Sci. USA 1996, 93, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, C. Geminivirus DNA Replication. Cell. Mol. Life Sci. 1999, 56, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Aregger, M.; Borah, B.K.; Seguin, J.; Rajeswaran, R.; Gubaeva, E.G.; Zvereva, A.S.; Windels, D.; Vazquez, F.; Blevins, T.; Farinelli, L. Primary and Secondary SiRNAs in Geminivirus-Induced Gene Silencing. PLoS Pathog. 2012, 8, e1002941. [Google Scholar] [CrossRef]
- Yang, Z.; Li, Y. Dissection of RNAi-Based Antiviral Immunity in Plants. Curr. Opin. Virol. 2018, 32, 88–99. [Google Scholar] [CrossRef]
- Bouché, N.; Lauressergues, D.; Gasciolli, V.; Vaucheret, H. An Antagonistic Function for Arabidopsis DCL2 in Development and a New Function for DCL4 in Generating Viral SiRNAs. EMBO J. 2006, 25, 3347–3356. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhou, H.; Zhang, Q.; Zhang, J.; Ni, F.; Liu, C.; Qi, Y. DNA Methylation Mediated by a MicroRNA Pathway. Mol. Cell 2010, 38, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Blevins, T.; Rajeswaran, R.; Aregger, M.; Borah, B.K.; Schepetilnikov, M.; Baerlocher, L.; Farinelli, L.; Meins, F., Jr.; Hohn, T.; Pooggin, M.M. Massive Production of Small RNAs from a Non-Coding Region of Cauliflower Mosaic Virus in Plant Defense and Viral Counter-Defense. Nucleic Acids Res. 2011, 39, 5003–5014. [Google Scholar] [CrossRef] [Green Version]
- Raja, P.; Wolf, J.N.; Bisaro, D.M. RNA Silencing Directed against Geminiviruses: Post-Transcriptional and Epigenetic Components. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2010, 1799, 337–351. [Google Scholar] [CrossRef]
- Rogans, S.J.; Allie, F.; Tirant, J.E.; Rey, M.E.C. Small RNA and Methylation Responses in Susceptible and Tolerant Landraces of Cassava Infected with South African Cassava Mosaic Virus. Virus Res. 2016, 225, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Chiumenti, M.; Morelli, M.; De Stradis, A.; Elbeaino, T.; Stavolone, L.; Minafra, A. Unusual Genomic Features of a Badnavirus Infecting Mulberry. J. Gen. Virol. 2016, 97, 3073–3087. [Google Scholar] [CrossRef]
- Pooggin, M.M. How Can Plant DNA Viruses Evade SiRNA-Directed DNA Methylation and Silencing? Int. J. Mol. Sci. 2013, 14, 15233–15259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinks, D.; Rajeswaran, R.; Shivaprasad, P.V.; Akbergenov, R.; Oakeley, E.J.; Veluthambi, K.; Hohn, T.; Pooggin, M.M. Suppression of RNA Silencing by a Geminivirus Nuclear Protein, AC2, Correlates with Transactivation of Host Genes. J. Virol. 2005, 79, 2517–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soitamo, A.J.; Jada, B.; Lehto, K. Expression of Geminiviral AC2 RNA Silencing Suppressor Changes Sugar and Jasmonate Responsive Gene Expression in Transgenic Tobacco Plants. BMC Plant Biol. 2012, 12, 204. [Google Scholar] [CrossRef] [Green Version]
- Mi, S.; Cai, T.; Hu, Y.; Chen, Y.; Hodges, E.; Ni, F.; Wu, L.; Li, S.; Zhou, H.; Long, C. Sorting of Small RNAs into Arabidopsis Argonaute Complexes Is Directed by the 5′ Terminal Nucleotide. Cell 2008, 133, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Iwasaki, S.; Watanabe, T.; Utsumi, M.; Watanabe, Y. The Mechanism Selecting the Guide Strand from Small RNA Duplexes Is Different among Argonaute Proteins. Plant Cell Physiol. 2008, 49, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, V.N. Sorting out Small RNAs. Cell 2008, 133, 25–26. [Google Scholar] [CrossRef] [Green Version]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 Participate in a DCL4-Initiated Antiviral RNA Silencing Pathway Negatively Regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Liu, H.-L.; Daxinger, L.; Pontes, O.; He, X.; Qian, W.; Lin, H.; Xie, M.; Lorkovic, Z.J.; Zhang, S. An RNA Polymerase II-and AGO4-Associated Protein Acts in RNA-Directed DNA Methylation. Nature 2010, 465, 106–109. [Google Scholar] [CrossRef] [Green Version]
- Hamera, S.; Song, X.; Su, L.; Chen, X.; Fang, R. Cucumber Mosaic Virus Suppressor 2b Binds to AGO4-Related Small RNAs and Impairs AGO4 Activities. Plant J. 2012, 69, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, C.; El Oirdi, M.; Adurogbangba, A.; Ma, X.; Moffett, P. Antiviral Defense Involves AGO4 in an Arabidopsis–Potexvirus Interaction. Mol. Plant-Microbe Interact. 2016, 29, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosseau, C.; Moffett, P. Functional and Genetic Analysis Identify a Role for Arabidopsis ARGONAUTE5 in Antiviral RNA Silencing. Plant Cell 2015, 27, 1742–1754. [Google Scholar] [CrossRef] [Green Version]
- Minoia, S.; Carbonell, A.; Di Serio, F.; Gisel, A.; Carrington, J.C.; Navarro, B.; Flores, R. Specific Argonautes Selectively Bind Small RNAs Derived from Potato Spindle Tuber Viroid and Attenuate Viroid Accumulation In Vivo. J. Virol. 2014, 88, 11933–11945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiumenti, M.; Greco, C.; Antelmi, I.; Sion, V.; Altamura, G.; Nigro, F.; Saldarelli, P. Molecular Characterisation of a Novel Gemycircularvirus Associated with Olive Trees in Italy. Virus Res. 2019, 263, 169–172. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leucocarpa (LC) | Oliastro (OL) | Toscanina (TS) | ||||
|---|---|---|---|---|---|---|
| Leaves | Fruit | Leaves | Fruit | Leaves | Fruit | |
| total | 12,340,147 | 7,813,730 | 8,218,400 | 6,545,789 | 9,488,906 | 2,968,628 |
| OEGV mapping reads (raw counts) | ||||||
| aligned exactly 1 time | 46,810 | 97,311 | 23,718 | 74,586 | 36,416 | 35,022 |
| aligned >1 time | 2178 | 4310 | 1045 | 5126 | 1999 | 3857 |
| total aligned reads | 48,988 | 101,621 | 24,763 | 79,712 | 38,415 | 38,879 |
| % aligned reads | 0.4 | 1.3 | 0.3 | 1.2 | 0.4 | 1.3 |
| DNA-A | 41,548 | 91,820 | 10,655 | 40,442 | 17,522 | 13,942 |
| DNA-B | 7440 | 9801 | 14,108 | 39,270 | 20,893 | 24,937 |
| Normalized OEGV mapping reads (RPKM) | ||||||
| DNA-A | 1213.30 | 4234.63 | 467.20 | 2226.42 | 665.43 | 1692.41 |
| DNA-B | 218.21 | 453.97 | 621.29 | 2171.29 | 796.90 | 3040.24 |
| Mean coverage of OEGV genomic components (X) | ||||||
| DNA-A | 324.0 | 710.9 | 85.0 | 318.1 | 137.4 | 110.4 |
| DNA-B | 59.8 | 76.6 | 114.7 | 311.4 | 169.1 | 197.8 |
| Coverage of OEGV genomic components (% of reference sequence) | ||||||
| DNA-A | 99.5 | 99.4 | 98.2 | 99.3 | 99.4 | 98.5 |
| DNA-B | 97.3 | 98.2 | 97.9 | 98.6 | 98.2 | 97.3 |
| Sequence | Position | Reference | SNP | Quality Score (Phred) | Genomic Region * |
|---|---|---|---|---|---|
| DNA-A | 175 | T | C | 27.08 | IR |
| 239 | C | A | 109 | CP | |
| 673 | G | T | 51 | CP | |
| 725 | T | G | 30.91 | CP | |
| 936 | A | C | 231 | CP | |
| 1021 | C | T | 258 | REn | |
| 1124 | C | T | 27.71 | TrAP/REn | |
| 1160 | G | A | 59 | TrAP/REn | |
| 1259 | G | C | 69 | TrAP/REn | |
| 1350 | T | C | 20.02 | TrAP/REn | |
| 1429 | C | T | 43.26 | TrAP | |
| 1462 | G | T | 329 | TrAP/Rep | |
| 1481 | T | C | 591 | TrAP/Rep | |
| 1761 | T | C | 106 | Rep | |
| 1771 | T | C | 20.03 | Rep | |
| 1915 | A | G | 273 | Rep | |
| 1954 | T | G | 55 | Rep | |
| DNA-B | 118 | A | T | 88 | IR |
| 169 | G | A | 245 | IR | |
| 181 | G | A | 33.64 | IR | |
| 988 | C | T | 999 | MP | |
| 1278 | A | T | 32.01 | Unknown | |
| 2084 | G | C | 397 | IR | |
| 2117 | T | C | 477 | IR | |
| 2178 | G | T | 24.69 | IR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiumenti, M.; Greco, C.; De Stradis, A.; Loconsole, G.; Cavalieri, V.; Altamura, G.; Zicca, S.; Saldarelli, P.; Saponari, M. Olea Europaea Geminivirus: A Novel Bipartite Geminivirid Infecting Olive Trees. Viruses 2021, 13, 481. https://doi.org/10.3390/v13030481
Chiumenti M, Greco C, De Stradis A, Loconsole G, Cavalieri V, Altamura G, Zicca S, Saldarelli P, Saponari M. Olea Europaea Geminivirus: A Novel Bipartite Geminivirid Infecting Olive Trees. Viruses. 2021; 13(3):481. https://doi.org/10.3390/v13030481
Chicago/Turabian StyleChiumenti, Michela, Claudia Greco, Angelo De Stradis, Giuliana Loconsole, Vincenzo Cavalieri, Giuseppe Altamura, Stefania Zicca, Pasquale Saldarelli, and Maria Saponari. 2021. "Olea Europaea Geminivirus: A Novel Bipartite Geminivirid Infecting Olive Trees" Viruses 13, no. 3: 481. https://doi.org/10.3390/v13030481