Differential Localization of Structural and Non-Structural Proteins during the Bluetongue Virus Replication Cycle


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
2.2. Antibodies and Reagents
2.3. Immunofluorescence Confocal and Structured Illumination Microscopy
2.4. Western Blot
2.5. Co-Immunoprecipitation Assay (Co-IP)
2.6. Differential Velocity Centrifugation
2.7. Statistical Analyses and Software
3. Results
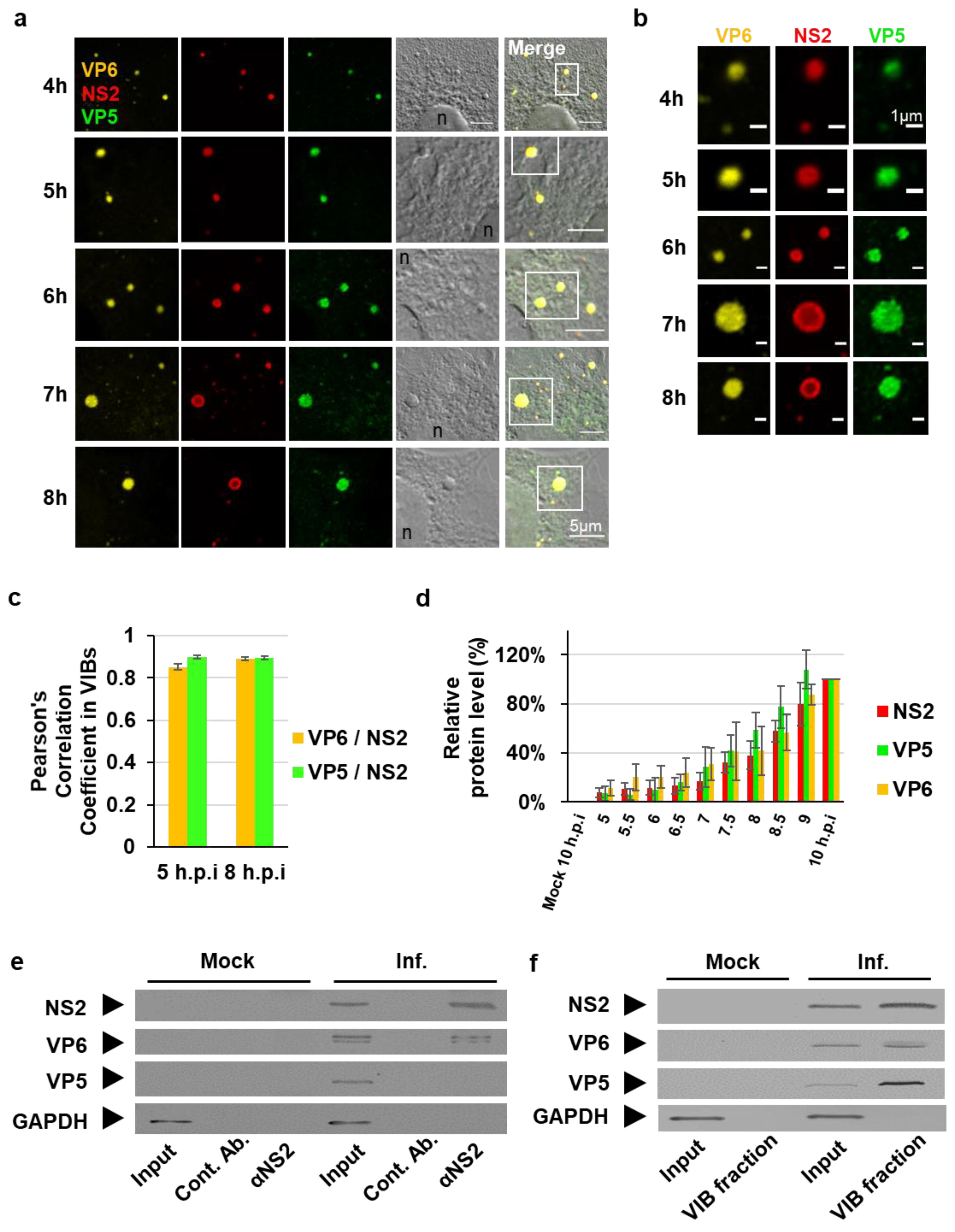
3.1. The Outer Capsid Protein VP5 Is Associated with VIBs
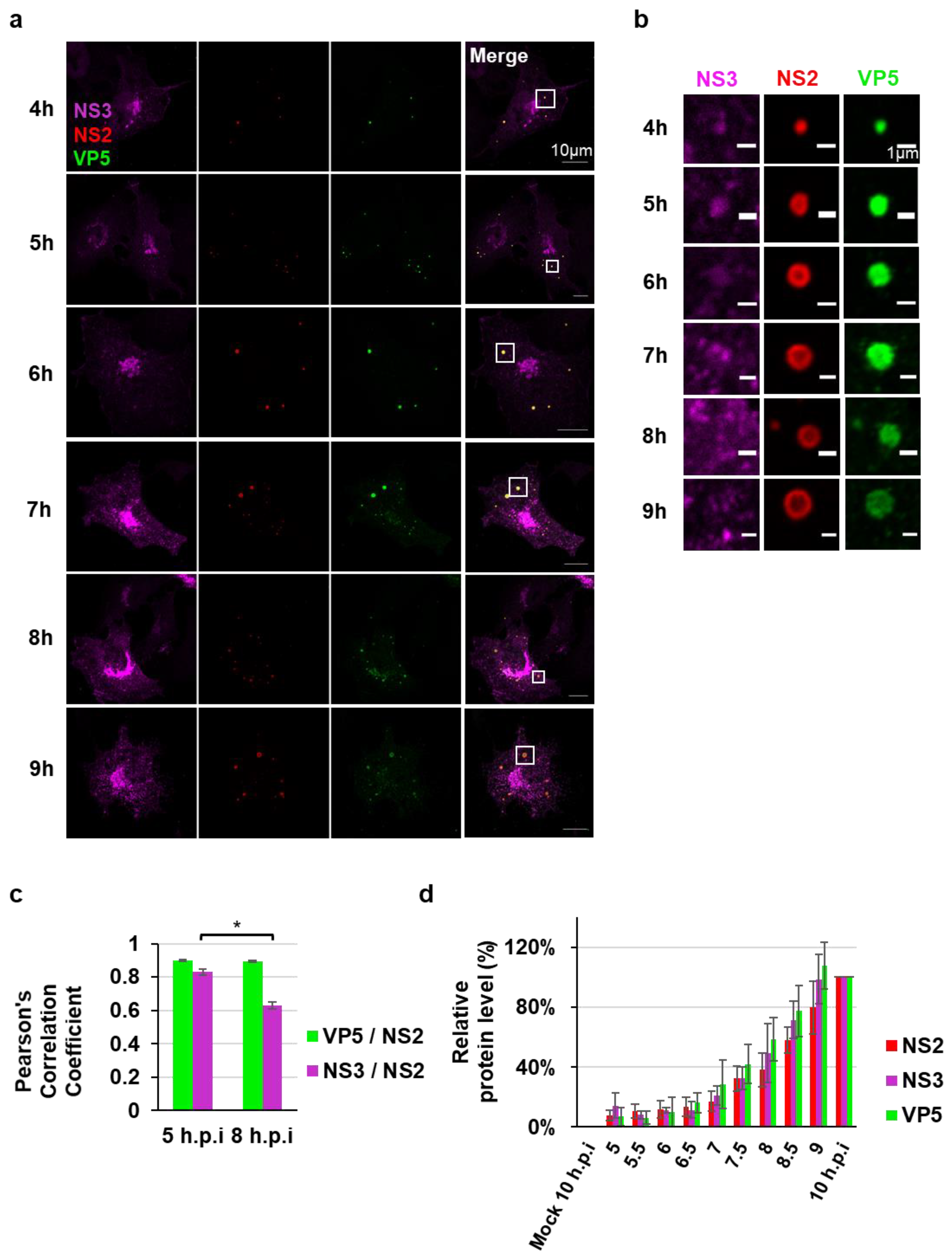
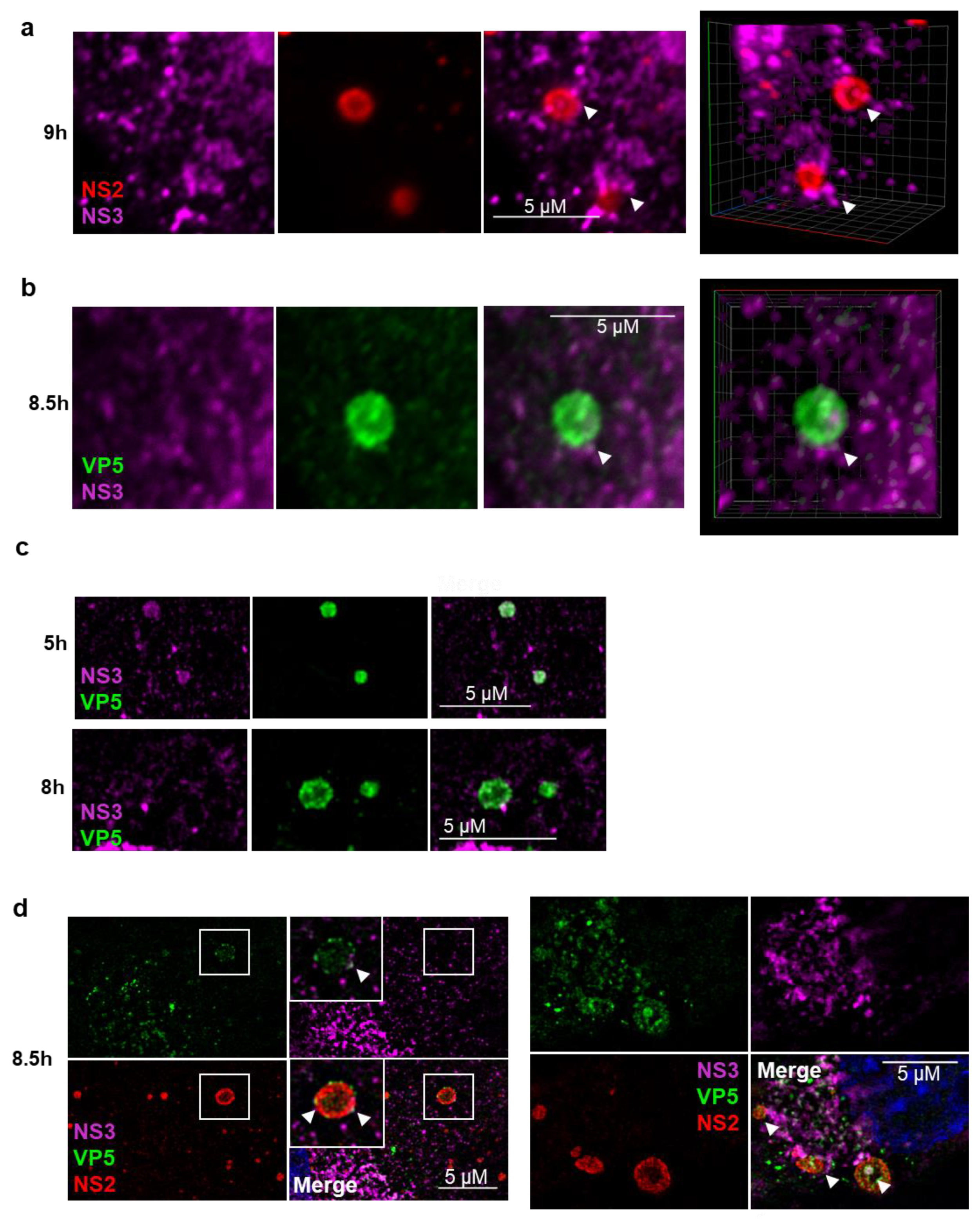
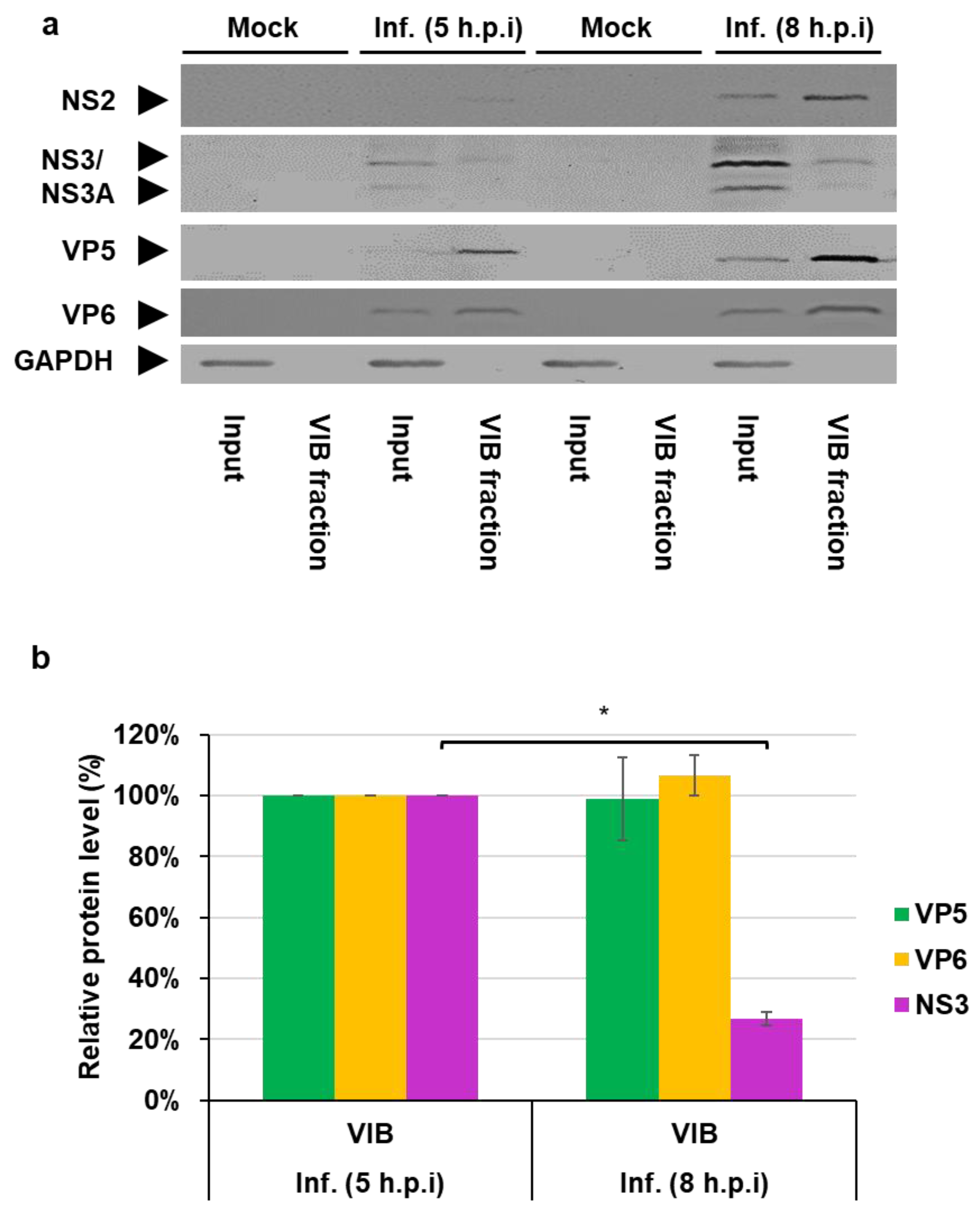
3.2. NS3 Association with VIBs During Viral Assembly
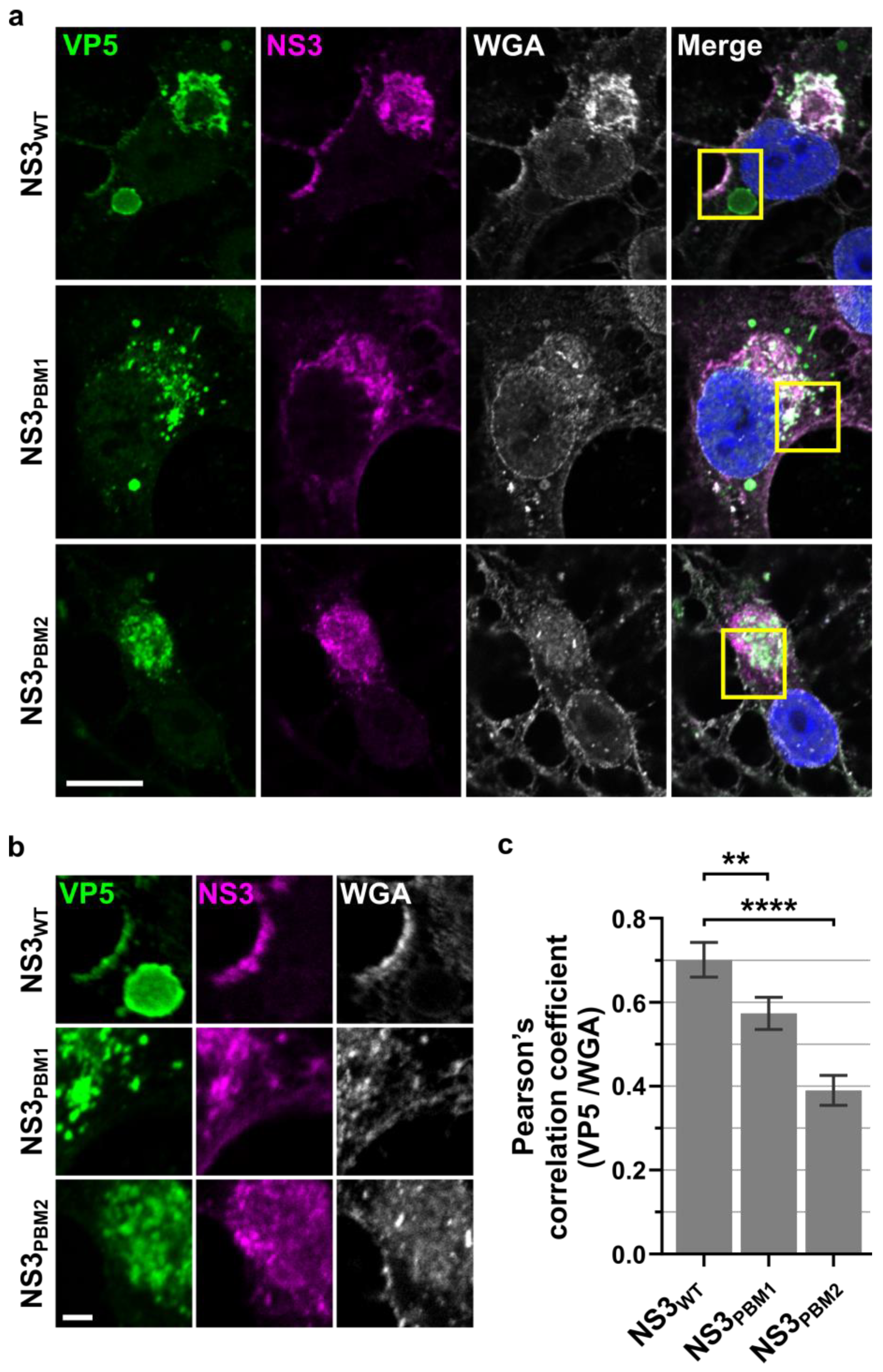
3.3. NS3 Trafficking Mutants Affect VP5 Export to the Plasma Membrane
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Verwoerd, D.W. Purification and characterization of bluetongue virus. Virology 1969, 38, 203–212. [Google Scholar] [CrossRef]
- Verwoerd, D.W.; Louw, H.; Oellermann, R.A. Characterization of bluetongue virus ribonucleic acid. J. Virol. 1970, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.A.; Zweerink, H.J. Isolation and characterization of two types of bluetongue virus particles. Virology 1972, 50, 495–506. [Google Scholar] [CrossRef]
- Mertens, P.P.; Brown, F.; Sangar, D.V. Assignment of the genome segments of bluetongue virus type 1 to the proteins which they encode. Virology 1984, 135, 207–217. [Google Scholar] [CrossRef]
- Ratinier, M.; Caporale, M.; Golder, M.; Franzoni, G.; Allan, K.; Nunes, S.F.; Armezzani, A.; Bayoumy, A.; Rixon, F.; Shaw, A.; et al. Identification and characterization of a novel non-structural protein of bluetongue virus. PLoS Pathog. 2011, 7, e1002477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forzan, M.; Wirblich, C.; Roy, P. A capsid protein of nonenveloped bluetongue virus exhibits membrane fusion activity. Proc. Natl. Acad. Sci. USA 2004, 101, 2100–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Patel, A.; Celma, C.C.; Yu, X.; Roy, P.; Zhou, Z.H. Atomic model of a nonenveloped virus reveals ph sensors for a coordinated process of cell entry. Nat. Struct. Mol. Biol. 2016, 23, 74–80. [Google Scholar] [CrossRef]
- Brookes, S.M.; Hyatt, A.D.; Eaton, B.T. Characterization of virus inclusion-bodies in bluetongue virus-infected cells. J. Gen. Virol. 1993, 74, 525–530. [Google Scholar] [CrossRef]
- Labadie, T.; Jegouic, S.; Roy, P. Bluetongue virus nonstructural protein 3 orchestrates virus maturation and drives non-lytic egress via two polybasic motifs. Viruses 2019, 11, 1107. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.P.; Booth, T.F.; Roy, P. Synthesis of bluetongue virus-encoded phosphoprotein and formation of inclusion bodies by recombinant baculovirus in insect cells: It binds the single-stranded rna species. J. Gen. Virol. 1990, 71, 2073–2083. [Google Scholar] [CrossRef]
- Huismans, H.; van Dijk, A.A.; Bauskin, A.R. In Vitro phosphorylation and purification of a nonstructural protein of bluetongue virus with affinity for single-stranded rna. J. Virol. 1987, 61, 3589–3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, A.K.; Bhattacharya, B.; Roy, P. Bluetongue virus rna binding protein ns2 is a modulator of viral replication and assembly. BMC Mol. Biol. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modrof, J.; Lymperopoulos, K.; Roy, P. Phosphorylation of bluetongue virus nonstructural protein 2 is essential for formation of viral inclusion bodies. J. Virol. 2005, 79, 10023–10031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, A.D.; Eaton, B.T.; Brookes, S.M. The release of bluetongue virus from infected cells and their superinfection by progeny virus. Virology 1989, 173, 21–34. [Google Scholar] [CrossRef]
- Matsuo, E.; Roy, P. Minimum requirements for bluetongue virus primary replication in vivo. J. Virol. 2013, 87, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Celma, C.C.; Roy, P. A viral nonstructural protein regulates bluetongue virus trafficking and release. J. Virol. 2009, 83, 6806–6816. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Chen, S.Y.; Iwata, H.; Compans, R.W.; Roy, P. Multiple glycoproteins synthesized by the smallest rna segment (s10) of bluetongue virus. J. Virol. 1992, 66, 7104–7112. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, A.D.; Gould, A.R.; Coupar, B.; Eaton, B.T. Localization of the non-structural protein ns3 in bluetongue virus-infected cells. J. Gen. Virol. 1991, 72, 2263–2267. [Google Scholar] [CrossRef]
- Celma, C.C.; Roy, P. Interaction of calpactin light chain (s100a10/p11) and a viral ns protein is essential for intracellular trafficking of nonenveloped bluetongue virus. J. Virol. 2011, 85, 4783–4791. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, B.; Celma, C.C.; Roy, P. Influence of cellular trafficking pathway on bluetongue virus infection in ovine cells. Viruses 2015, 7, 2378–2403. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, B.; Roy, P. Bluetongue virus outer capsid protein vp5 interacts with membrane lipid rafts via a snare domain. J. Virol. 2008, 82, 10600–10612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, A.K.; Iwatani, N.; Roy, P. Assembly and intracellular localization of the bluetongue virus core protein vp3. J. Virol. 2005, 79, 11487–11495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, M.; Celma, C.C.; Roy, P. Development of reverse genetics systems for bluetongue virus: Recovery of infectious virus from synthetic rna transcripts. J. Virol. 2008, 82, 8339–8348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, A.A.; Huismans, H. The effect of temperature on the in vitro transcriptase reaction of bluetongue virus, epizootic haemorrhagic disease virus and african horsesickness virus. Onderstepoort J. Vet. Res. 1982, 49, 227–232. [Google Scholar] [PubMed]
- Huismans, H.; Van Dijk, A.A.; Els, H.J. Uncoating of parental bluetongue virus to core and subcore particles in infected l cells. Virology 1987, 157, 180–188. [Google Scholar] [CrossRef]
- Ramadevi, N.; Roy, P. Bluetongue virus core protein vp4 has nucleoside triphosphate phosphohydrolase activity. J. Gen. Virol. 1998, 79, 2475–2480. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, A.A.; Huismans, H. The In Vitro activation and further characterization of the bluetongue virus-associated transcriptase. Virology 1980, 104, 347–356. [Google Scholar] [CrossRef]
- Matsuo, E.; Roy, P. Bluetongue virus vp6 acts early in the replication cycle and can form the basis of chimeric virus formation. J. Virol. 2009, 83, 8842–8848. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, B.; Roy, P. Cellular phosphoinositides and the maturation of bluetongue virus, a non-enveloped capsid virus. Virol. J. 2013, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Netherton, C.; Moffat, K.; Brooks, E.; Wileman, T. A guide to viral inclusions, membrane rearrangements, factories, and viroplasm produced during virus replication. Adv. Virus Res. 2007, 70, 101–182. [Google Scholar]
- Tenorio, R.; Fernandez de Castro, I.; Knowlton, J.J.; Zamora, P.F.; Sutherland, D.M.; Risco, C.; Dermody, T.S. Function, architecture, and biogenesis of reovirus replication neoorganelles. Viruses 2019, 11, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criglar, J.M.; Anish, R.; Hu, L.; Crawford, S.E.; Sankaran, B.; Prasad, B.V.V.; Estes, M.K. Phosphorylation cascade regulates the formation and maturation of rotaviral replication factories. Proc. Natl. Acad. Sci. USA 2018, 115, E12015–E12023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Shivakoti, S.; Ding, K.; Cui, Y.; Roy, P.; Zhou, Z.H. In Situ structures of rna-dependent rna polymerase inside bluetongue virus before and after uncoating. Proc. Natl. Acad. Sci. USA 2019, 116, 16535–16540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawton, J.A.; Estes, M.K.; Prasad, B.V.V. Three-dimensional visualization of mrna release from actively transcribing rotavirus particles [letter]. Nat. Struct. Biol. 1997, 4, 118–121. [Google Scholar] [CrossRef]
- Periz, J.; Celma, C.; Jing, B.; Pinkney, J.N.; Roy, P.; Kapanidis, A.N. Rotavirus mrnas are released by transcript-specific channels in the double-layered viral capsid. Proc. Natl. Acad. Sci. USA 2013, 110, 12042–12047. [Google Scholar] [CrossRef] [Green Version]
- Wirblich, C.; Bhattacharya, B.; Roy, P. Nonstructural protein 3 of bluetongue virus assists virus release by recruiting escrt-i protein tsg101. J. Virol. 2006, 80, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Beaton, A.R.; Rodriguez, J.; Reddy, Y.K.; Roy, P. The membrane trafficking protein calpactin forms a complex with bluetongue virus protein ns3 and mediates virus release. Proc. Natl. Acad. Sci. USA 2002, 99, 13154–13159. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohl, B.-P.; Kerviel, A.; Labadie, T.; Matsuo, E.; Roy, P. Differential Localization of Structural and Non-Structural Proteins during the Bluetongue Virus Replication Cycle. Viruses 2020, 12, 343. https://doi.org/10.3390/v12030343
Mohl B-P, Kerviel A, Labadie T, Matsuo E, Roy P. Differential Localization of Structural and Non-Structural Proteins during the Bluetongue Virus Replication Cycle. Viruses. 2020; 12(3):343. https://doi.org/10.3390/v12030343
Chicago/Turabian StyleMohl, Bjorn-Patrick, Adeline Kerviel, Thomas Labadie, Eiko Matsuo, and Polly Roy. 2020. "Differential Localization of Structural and Non-Structural Proteins during the Bluetongue Virus Replication Cycle" Viruses 12, no. 3: 343. https://doi.org/10.3390/v12030343