Molecular Detection of Rabies Lyssaviruses from Dogs in Southeastern Nigeria: Evidence of TransboundaryTransmission of Rabies in West Africa

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
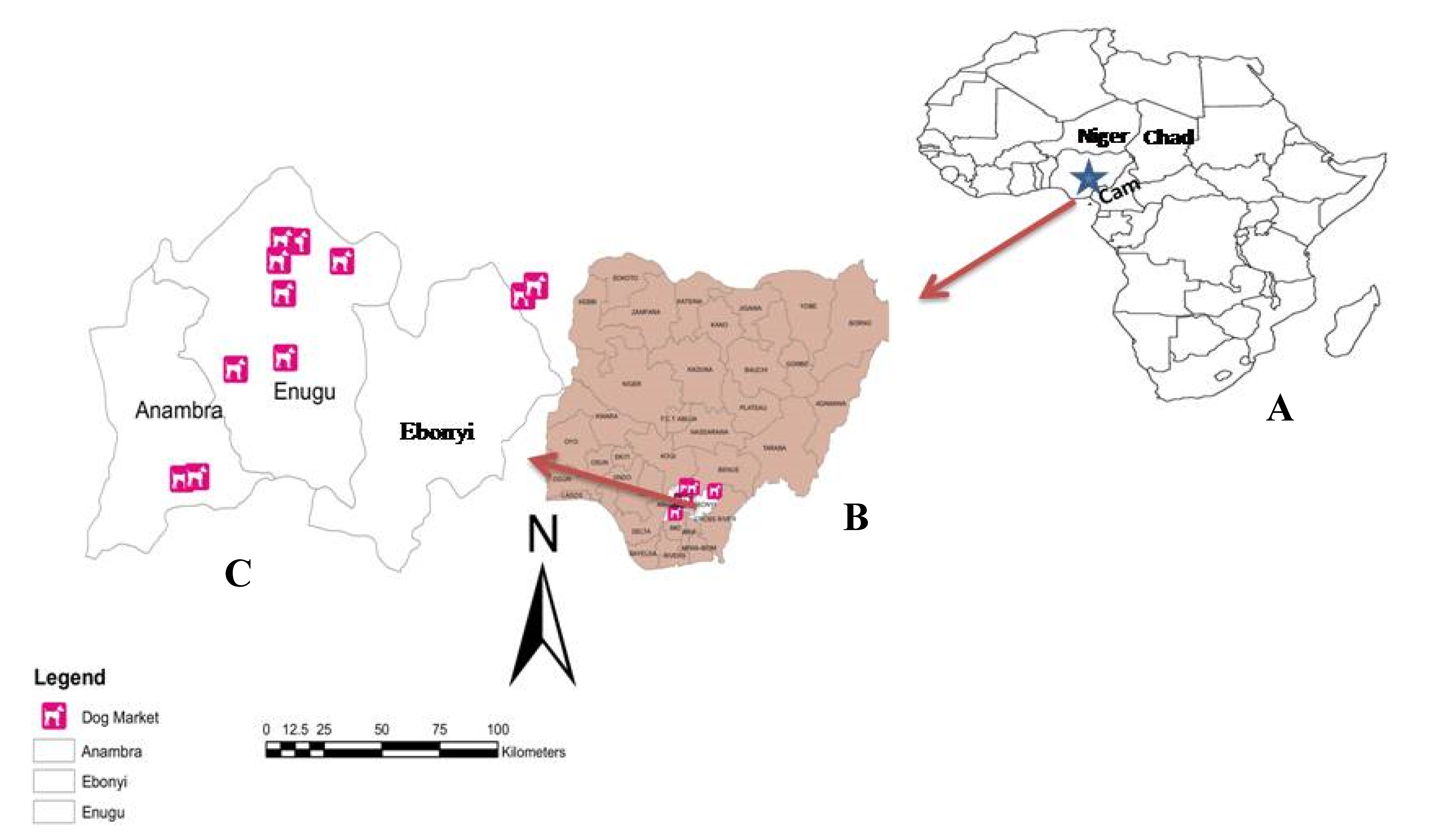
2.2. Study Locations
2.3. Specimen Collection
2.4. Direct Fluorescent Antibody Test for Detection of Rabies Lyssavirus Antigen
2.5. RNA Extraction
2.6. Detection of Nucleoprotein (N) Gene of Lyssaviruses Using Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.7. Reverse Transcription (RT)
2.8. Polymerase Chain Reaction (PCR) Assay
2.9. Electrophoresis and Gel Image Documentation
2.10. PCR Product Purification and Sequencing
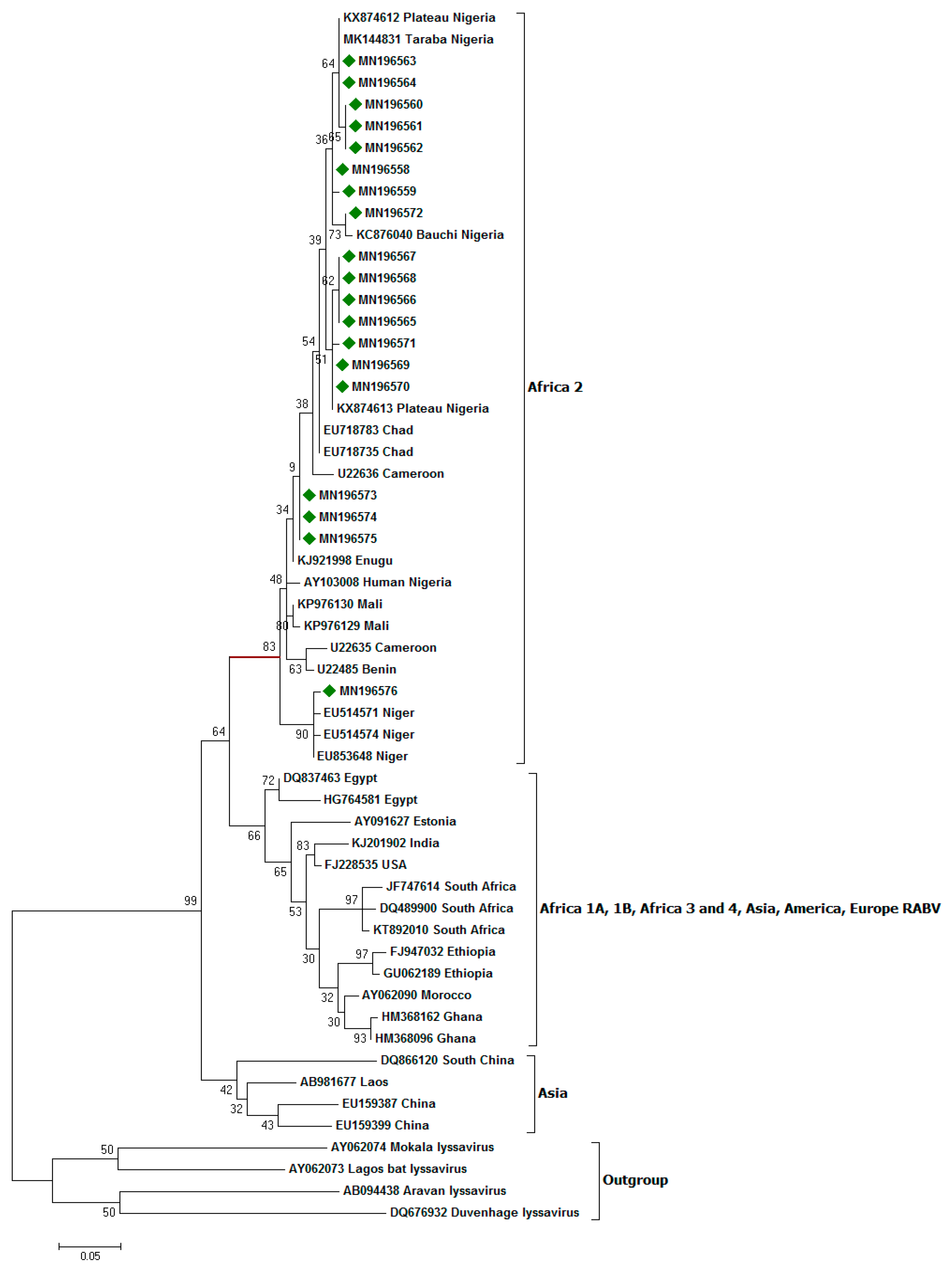
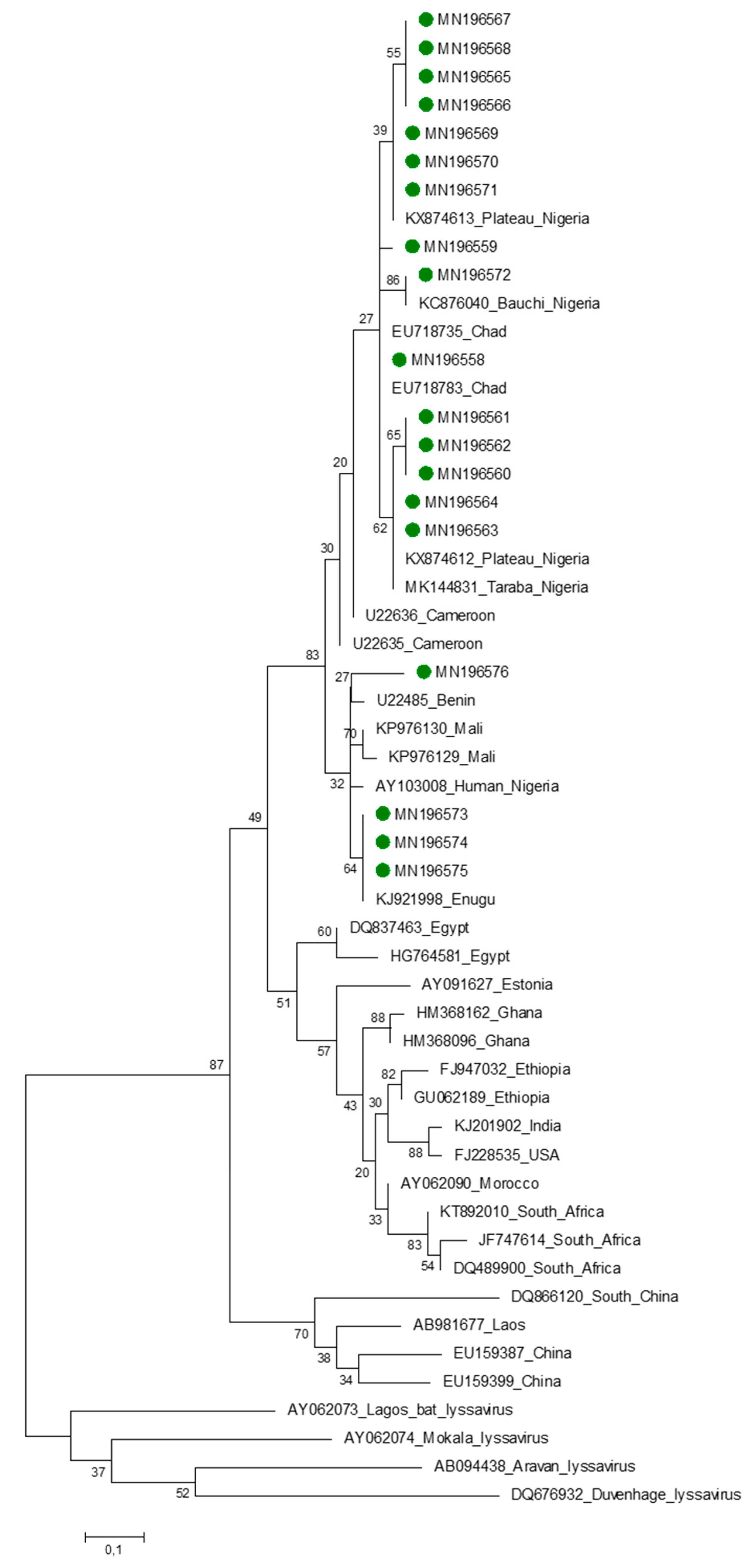
2.11. Phylogenetic Analysis
3. Results
3.1. Quality of the Extracted RNA
3.2. Gel Electrophoresis Result
3.3. Nucleoprotein Gene Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Umoh, J.U.; Belino, E.D. Rabiesin Nigeria: Ahistorical review. Int. J. Zoonosis 1978, 6, 41–48. [Google Scholar]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the global burden of endemic canine rabies. PLoS Negl. Trop. Dis. 2015, 9, e0003709. [Google Scholar]
- Fooks, A.R. Investigation of a human case of rabies in the United Kingdom. J. Clin. Virol. 2002, 25, 351–356. [Google Scholar]
- International Committeeon Taxonomy of Viruses (ICTV). Online 2018b Release EC50, Washington, DC.10th Report of the International Committee on Taxonomy of Viruses. 2018. Available online: http://www.ictvonline.org/virusTaxonomy.asp (accessed on 19 May 2019).
- Kemp, G.E.; Causey, O.R.; Moore, D.L.; Odeola, A.; Fabiyi, A. Mokola Virus. Further studies on IbAn 27377, a new etiologic agent of zoonosis in Nigeria. Am. J. Trop. Med. Hyg. 1972, 21, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Kemp, G.E.; Moore, D.L.; Isoun, T.T.; Fabiyi, A. Mokola virus: Experimental infection and transmission studies with the shrew, a natural host. Arch. Virusforsch. 1973, 43, 242–250. [Google Scholar] [CrossRef] [PubMed]
- USAID. Strategic Review, Feed the Future, United States Agency for International Development/West Africa. 2010. Available online: http://feedthefuture.gov/resource/feed-future-guide (accessed on 6 April 2019).
- United Nations Department of Economic and Social Affairs (UNDES), Population Division. World Population Prospects: The 2017 Revision, Custom Data Acquired via Website; UNDES: New York, NY, USA, 2017. [Google Scholar]
- Silberfein, M.; Conteh, A.H. Boundaries and Conflict in the Mano River Region of West Africa. Confl. Manag. Peace Sci. 2006, 23, 343–361. [Google Scholar] [CrossRef]
- Sunday, O.V.O.; Okechukwu, O.; Okechukwu, R. Cross Border Crimes in West African Sub-Region: Implications for Nigeria’s National Security and External Relations. Glob. J. Hum. Soc. Sci. F Political Sci. 2014, 14, 45–57. [Google Scholar]
- Ogunkoya, A.B. Review of rabies and problems of rabies in Nigeria. In Proceedings of the National Conference/Work on Rabies; Ahmadu Bello University: Zaria, Nigeria, 2008; pp. 62–70. [Google Scholar]
- Ogo, M.F.; Nel, L.; Sabeta, C.T. Phylogenetic Evidence of the Public and Veterinary Health Threat of Dog Rabies in Nigeria. Niger. Vet. J. 2011, 32, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Hayman, D.T.S.; Johnson, N.; Horton, D.L.; Hedge, J.; Wakeley, P.R.; Banyard, A.C.; Zhang, S.; Alhassan, A.; Fooks, A.R. Evolutionary History of Rabies in Ghana. PLoS Negl. Trop. Dis. 2011, 5, e1001. [Google Scholar] [CrossRef]
- Nadin-Davis, S.A.; Turner, G.; Paul, J.P.V.; Madhusudana, S.N.; Wandeler, A.I. Emergence of Arctic-like Rabies Lineage in India. Emerg. Infect. Dis. J. 2007, 31, 111–116. [Google Scholar] [CrossRef]
- Hyun, B.H.; Lee, K.K.; Kim, I.J.; Lee, K.W.; Park, H.J.; Lee, O.S. Molecular epidemiology of rabies virus isolates from South Korea. Virus Res. 2005, 114, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Botvinkin, A.D.; McElhinney, L.M.; Smith, J.S.; Orciari, L.A.; Hughes, G.J. Molecular epidemiology of terrestrial rabies in the former Soviet Union. J. Wildl. Dis. 2004, 40, 617–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansfield, K.L.; Racloz, V.; McElhinney, L.M.; Marston, D.A.; Johnson, N.; Ronsholt, L. Molecular epidemiological study of Arctic rabies virus isolates from Greenland and comparison with isolates from throughout the Arctic and Baltic regions. Virus Res. 2006, 116, 1–10. [Google Scholar] [CrossRef]
- Kissi, B.; Tordo, N.; Bourhy, H. Genetic polymorphism in the rabies virus nucleoprotein gene. Virology 1995, 209, 526–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbi, C.; Holmes, E.C.; de Benedictis, P.; Faye, O.; Gamatie, E.N.D.; Diarra, A.; Elmamy, B.O.; Sow, A.; Adjogoua, E.V.; Sangare, O.; et al. Evolutionary history and dynamics of dog rabies virus in western and central Africa. J. Gen. Virol. 2009, 90, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Olarinmoye, A.O.; Kamara, V.; Jomah, N.D.; Olugasa, B.O.; Ishola, O.O.; Kamara, A.; Luka, P.D. Molecular detection of rabies virus strain with N-gene that clustered with China lineage 2 co-circulating with Africa lineages in Monrovia, Liberia: First reported case in Africa. Epidemiol. Infect. 2019, 147, e85. [Google Scholar] [CrossRef] [Green Version]
- Sabeta, C.T.; Mansfield, K.L.; McElhiney, L.M.; Fooks, A.R.; Nel, L.H. Molecular epidemiology of rabies in bat-eared foxes (Otocyonmegalotis) in South Africa. Virus Res. 2007, 129, 1–10. [Google Scholar] [CrossRef]
- David, D.; Osinubi, M.O.V.; Nok, A.J.; Ogunkoya, A.B.; Umoh, J.U.; Balogun, E.; Yakobson, B.A. Molecular epidemiology of canine rabies isolates from Plateau State Nigeria. In Proceedings of the National Conference/Workshop on Rabies; IDR-ABU: Zaria, Nigeria, 2008; pp. 84–87. [Google Scholar]
- Garba, A. Epidemiology of Rabies in Niger State, Nigeria. Ph.D. Thesis, Ahmadu Bello University, Zaria, Nigeria, 2015. [Google Scholar]
- Kia, G.S.N.; Huang, Y.; Zhou, M.; Zhou, Z.; Gnanadurai, C.W.; Leysona, C.M.; Umoh, J.U.; Kazeem, H.M.; Ehizibolo, D.O.; Kwaga, J.K.P.; et al. Molecular characterization of a rabies virus isolated from trade dogs in Plateau State, Nigeria. Sokoto J. Vet. Sci. 2018, 16, 54–62. [Google Scholar] [CrossRef]
- Eze, U.U.; Anene, B.M.; Chukwu, C.C.; Ogunkoya, A.B.; George, S.O.; Eze, J.I.; Animoke, P.C.; Nwosuh, C.I. Risk of typical rabies in dog meat-eating human population, in Enugu, Nigeria. Int. J. Public Health Epidemiol. 2015, 4, 107–109. [Google Scholar]
- Chigere, N.H. Foreign Missionary Background and Indigenious Evangelization in Igboland: Igboland and the Igbo People of Nigeria; Transaction Publishers: Piscataway, NJ, USA, 2000; p. 17. [Google Scholar]
- Dean, D.J.; Abelseth, M.K.; Atanasiu, P. The fluorescent antibody test. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.-X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 88–95. [Google Scholar]
- Heaton, P.R.; Johnstone, P.; McElhinney, L.M.; Cowley, R.; O’Sullivan, E.; Whitby, J.E. Heminested PCR assay for the detection of six genotypes of rabies and rabies related viruses. J. Clin. Microbiol. 1997, 35, 2762–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markotter, W.; Kuzmin, I.; Rupprecht, C.E.; Randles, J.; Sabeta, C.T.; Wandeler, A.I.; Nel, L.H. Isolation of Lagos bat virus from water mongoose. Emerg. Infect. Dis. 2006, 12, 1913–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tordo, N.; Poch, O.; Ermine, A.; Keith, G.; Rougeon, F. Walking along the rabies genome: Is the large G-Lintergenic region are mnantgene? Proc. Natl. Acad. Sci. USA 1986, 83, 3914–3918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, S.A.; Higuchi, R. Avoiding false positives with PCR. Nature 1989, 339, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: Auser-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symp. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Strecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetic analysis version 7.0 for bigger dataset. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.1. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Hills, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- De Benedictis, P.; Sow, A.; Fusaro, A.; Veggiato, C.; Talbi, C.; Kabore, A.; Dundon, W.G.; Bourhy, H.; Capua, I. Phylogenetic analysis of rabies viruses from Burkina Faso, 2007. Zoonoses Public Health 2009, 57, 42–46. [Google Scholar] [CrossRef]
- Ogunkoya, A.B.; Beran, G.W.; Wiktor, T.J. Antigenic analysis of some Nigerian street rabies virus using monoclonal antibodies. Niger. J. 2000, 27, 241–253. [Google Scholar]
- Smith, J.S.; Pamela, A.; Yager, P.A.; Bigler, W.J.; Eldert, C.; Hartwig, E.C. Surveillance and epidemiologic mapping of monoclonal antibody-defined rabies variants in Florida. J. Wildl. Dis. 1990, 26, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Paez, A.; Nunez, C.; Grarcia, C.; Bushel, J. Molecular epizootics in Columbia: Evidence for human and dog rabies associated with bats. J. Gen. Virol. 2003, 84, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.; Letshwenyo, M.; Baipoledi, E.K.; Thebokwe, G.; Fooks, A.R. Molecular epidemiology of rabies in Botswana: A comparism between antibody typing and nucleotide sequence phylogeny. Vet. Microbiol. 2004, 101, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Nel, L.H.; Sabeta, C.T.; Von Teichman, B.; Jaftha, J.B.; Rupprecht, C.E.; Bingham, J. Mongoose rabies in southern Africa: Are-evaluation based on molecular epidemiology. Virus Res. 2005, 109, 165–173. [Google Scholar] [CrossRef]
- Van Zyl, N.; Markotter, W.; Nel, L.H. Evolutionary history of African Mongoose rabies virus. Virus Res. 2010, 150, 93–102. [Google Scholar] [CrossRef]
- Wiktor, T.J.; Marcfalan, R.I.; Foggin, C.M.; Koprowski, H. Antigenic analysis of rabies and Mokola virus from Zimbab we using monoclonal antibodies. Dev. Biol. Stand. 1984, 57, 199–211. [Google Scholar]
- Sabo, G.; Umoh, J.U.; Sackey, A.K.B.; Ahamd, A.; Okolocha, E.C. The role of dog trade in the epidemiology of rabies: A review. Vet. Clin. Pract. Bull. 2008, 1, 63–72. [Google Scholar]
- Fevre, E.M.; Kaboyo, R.W.; Persson, V.; Edelsten, M.; Coleman, P.G.; Cleaveland, S. The epidemiology of animal bite injuries in Uganda and projections of the burden of rabies. Trop. Med. Int. Health 2005, 10, 790–798. [Google Scholar] [CrossRef]
- Bourhy, H.; Reynes, J.M.; Dunham, E.J.; Dacheux, L.; Larrous, F.; Huong, V.T.Q.; Xu, G.; Yan, J.; Miranda, M.E.G.; Holmes, E.C. The origin and phylogeography of dog rabies virus. J. Gen. Virol. 2008, 89, 2673–2681. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Sense | Nucleotide Sequence 5′-3′ | Uses | Position on Genome |
|---|---|---|---|
| JW12 (+) | ATGTAACACC(C/T)CTACAATTG | cDNA synthesis, PCR, hnPCR | 55–74 |
| 001lys (+) | ACGCTTAACGAMAAA | cDNA synthesis | 1–15 |
| 304 (-) | TTGACAAAGATCTTGCTCAT | PCR and sequencing | 1514–1533 |
| 550B (-) | GTRCTCCARTTAGCRCACAT | hnPCR | 647–666 |
| Genbank Accession Number | Year of Collection | Country | Host Species | Lineage |
|---|---|---|---|---|
| MK144831 | 2014 | Taraba, Nigeria | Dog | Africa 2 |
| KX874612 | 2010 | Jos, Nigeria | Dog | Africa 2 |
| KX874613 | 2010 | Jos, Nigeria | Dog | Africa 2 |
| KC876040 | 2012 | Bauchi, Nigeria | Cattle | Africa 2 |
| KJ 921998 | 2012 | Enugu, Nigeria | Dog | Africa 2 |
| EU514571 | 2007 | Niger | Dog | Africa 2 |
| EU514574 | 2007 | Niger | Dog | Africa 2 |
| EU853648 | 1990 | Niger | Dog | Africa 2 |
| EU718783 | 2006 | Chad | Dog | Africa 2 |
| EU718735 | 2006 | Chad | Dog | Africa 2 |
| KP976130 | 2007 | Mali | Canis lupus familiaris | Africa 2 |
| KP976129 | 2007 | Mali | Canis lupus familiaris | Africa 2 |
| AY103008 | 1996 | Nigeria | Human | Africa 2 |
| U22636 | 1995 | Cameroon | Dog | Africa 2 |
| U22635 | 1995 | Cameroon | Dog | Africa 2 |
| U22485 | 1995 | Benin | Dog | Africa 2 |
| DQ837463 | 1999 | Egypt | Dog | Africa 4 |
| HG764581 | 2013 | Egypt | Cattle | Africa 4 |
| HM368162 | 2007 | Ghana | Dog | Africa 1A |
| HM368096 | 2008 | Ghana | Dog | Africa 1A |
| AY062090 | 2001 | Morocco | Fox | Africa 1B |
| FJ947032 | 2009 | Ethiopia | Dog | Africa 1A |
| GU062189 | 2009 | Ethiopia | Canissimensis | Africa 1A |
| AY091627 | 2002 | Estonia | Raccoon dog | Europe |
| KJ201902 | 2010 | India | Musmusculus | Asia |
| FJ228535 | 1950 | USA | Dog | America |
| JF747614 | 2008 | South Africa | Canine | Africa 3 |
| KT892010 | 1997 | South Africa | African wild dog | Africa3 |
| DQ194892 | 1997 | South Africa | African wild dog | Africa 3 |
| DQ 866120 | 2006 | South China | Dog | Asia (China) |
| AB981677 | 2012 | Laos | Canis lupus familiaris | Asia |
| EU159399 | 1994 | China | Dog | Asia |
| AB094438 | 2002 | Kyrgyzstan | Myotisblythii | Aravanlyssairus (Outgroup) |
| DQ676932 | 2006 | South Africa | Human | Duvenhagelyssavirus (Outgroup) |
| AY062073 | 2002 | South Africa | Bat | Lagos bat Lyssavirus (Outgroup) |
| AY062074 | 2001 | Nigeria | Shrew | Mokolalyssavirus (Outgroup) |
| S/N | Lab ID | Date of Collection | Source Location | Accession Number |
|---|---|---|---|---|
| 1 | 14NG | 11 October 2015 | OrieOrba, Enugu State, Nigeria | MN196558 |
| 2 | 11NG | 26 October 2015 | OrieOrba, Enugu State, Nigeria | MN196559 |
| 3 | 18NG | 7 November 2015 | OrieOrba, Enugu State, Nigeria | MN196560 |
| 4 | 12NG | 7 November 2015 | OrieOrba, Enugu State Nigeria | MN196561 |
| 5 | 7NG | 6 December 2015 | OrieOrba, Enugu State Nigeria | MN196562 |
| 6 | 17NG | 10 December 2015 | OrieOrba, Enugu State Nigeria | MN196563 |
| 7 | 1NG | 19 December 2015 | OrieOrba, Enugu State Nigeria | MN196564 |
| 8 | 20NG | 9 January 2016 | NkwoOgbede Enugu State, Nigeria | MN196565 |
| 9 | 26NG | 10 January 206 | IheNze, Enugu State, Nigeria | MN196566 |
| 10 | 28NG | 17 January 2016 | IheNze, Enugu State, Nigeria | MN196567 |
| 11 | 29NG | 22 February 2016 | Umuna, Enugu State, Nigeria | MN196568 |
| 12 | 23NG | 18 April 2016 | Iboko, Ebonyi State, Nigeria | MN196569 |
| 13 | 13NG | 28 April 2016 | Nkwoogbede, Enugu State | MN196570 |
| 14 | 19NG | 16 May 2016 | OrieOrba, Enugu State Nigeria | MN196571 |
| 15 | 22NG | 16 May 2016 | OrieOrba, Enugu State Nigeria | MN196572 |
| 16 | 21NG | 29 May 2016 | NkwoIgboukwuAnambra State, Nigeria | MN196573 |
| 17 | 27NG | 7 June 2016 | NkwoIgboukwuAnambra State, Nigeria | MN196574 |
| 18 | 25NG | 7 June 2016 | NkwoIgboukwuAnambra State, Nigeria | MN196575 |
| 19 | 16NG | 20 July 2016 | Vet Clinic Uwani Enugu, Nigeria | MN196576 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eze, U.U.; Ngoepe, E.C.; Anene, B.M.; Ezeokonkwo, R.C.; Nwosuh, C.I.; Sabeta, C.T. Molecular Detection of Rabies Lyssaviruses from Dogs in Southeastern Nigeria: Evidence of TransboundaryTransmission of Rabies in West Africa. Viruses 2020, 12, 134. https://doi.org/10.3390/v12020134
Eze UU, Ngoepe EC, Anene BM, Ezeokonkwo RC, Nwosuh CI, Sabeta CT. Molecular Detection of Rabies Lyssaviruses from Dogs in Southeastern Nigeria: Evidence of TransboundaryTransmission of Rabies in West Africa. Viruses. 2020; 12(2):134. https://doi.org/10.3390/v12020134
Chicago/Turabian StyleEze, Ukamaka U, Ernest C Ngoepe, Boniface M Anene, Romanus C Ezeokonkwo, Chika I Nwosuh, and Claude T Sabeta. 2020. "Molecular Detection of Rabies Lyssaviruses from Dogs in Southeastern Nigeria: Evidence of TransboundaryTransmission of Rabies in West Africa" Viruses 12, no. 2: 134. https://doi.org/10.3390/v12020134