
Variations in BK Polyomavirus Immunodominant Large Tumor Antigen-Specific 9mer CD8 T-Cell Epitopes Predict Altered HLA-Presentation and Immune Failure


Abstract
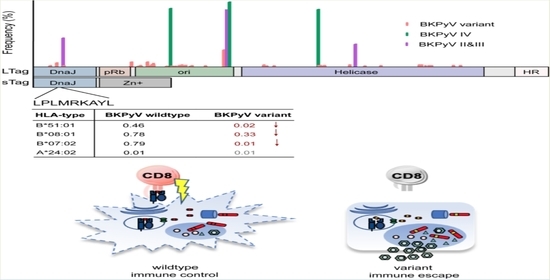
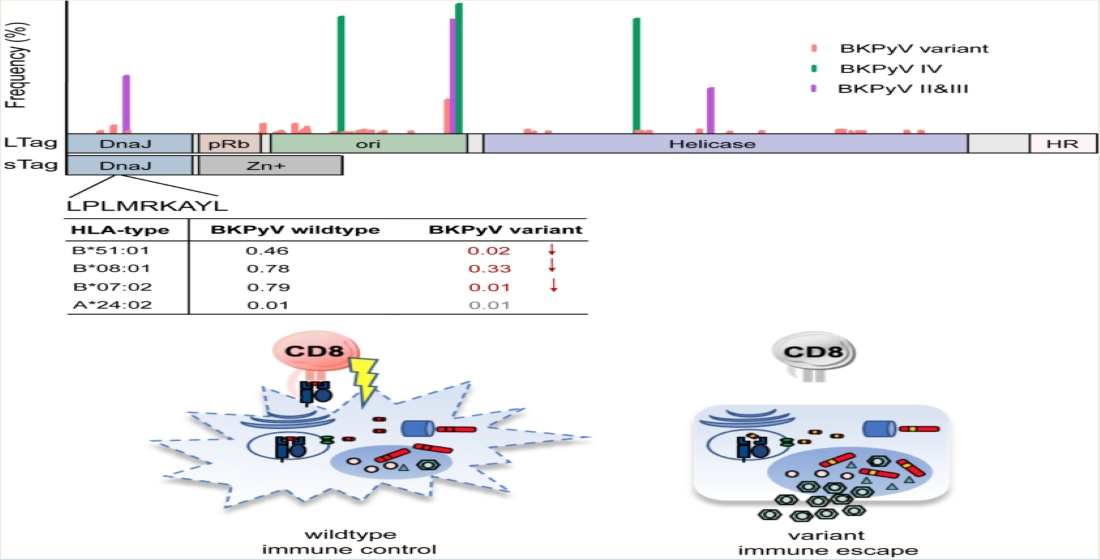
:
1. Introduction
2. Materials and Methods
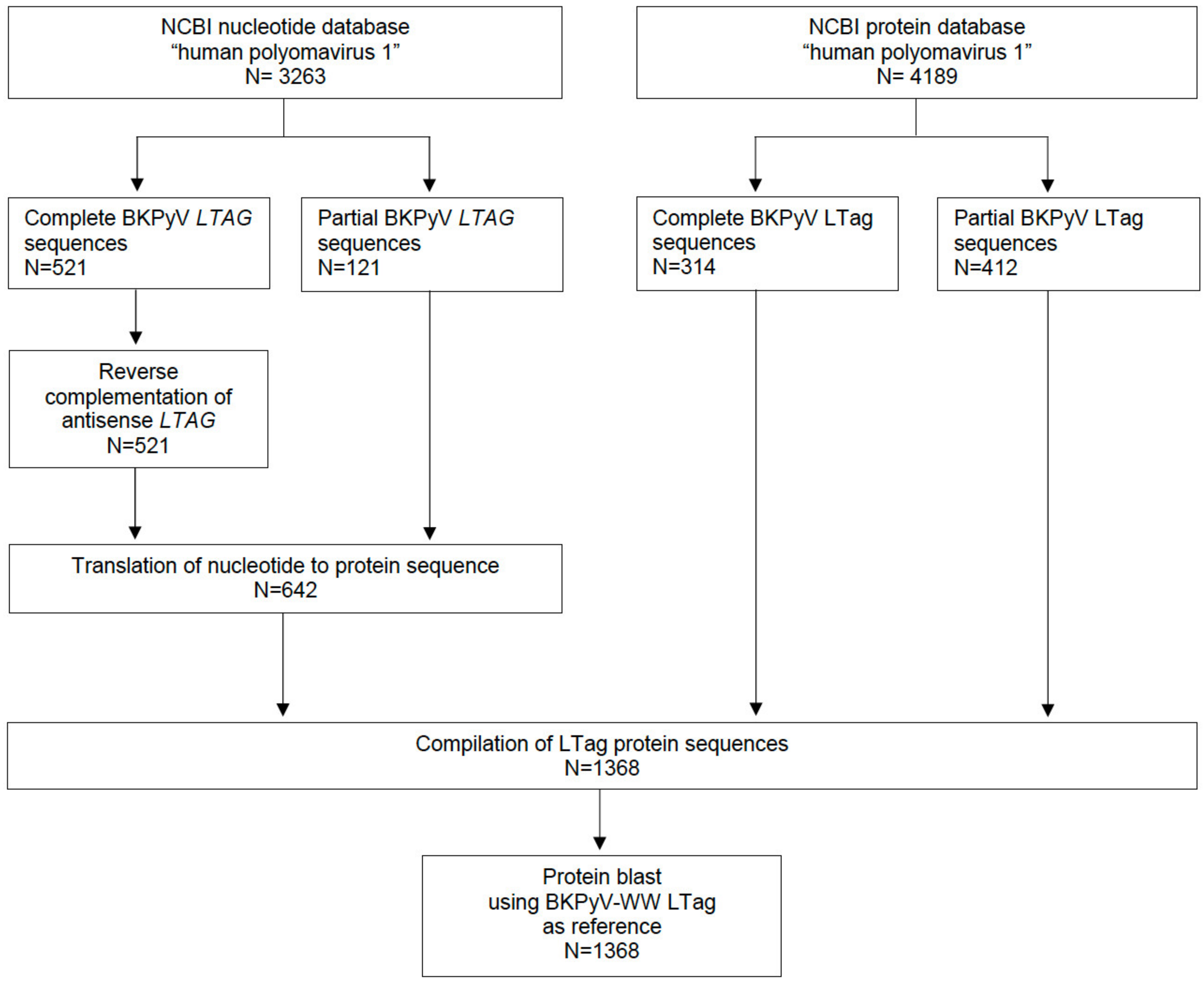
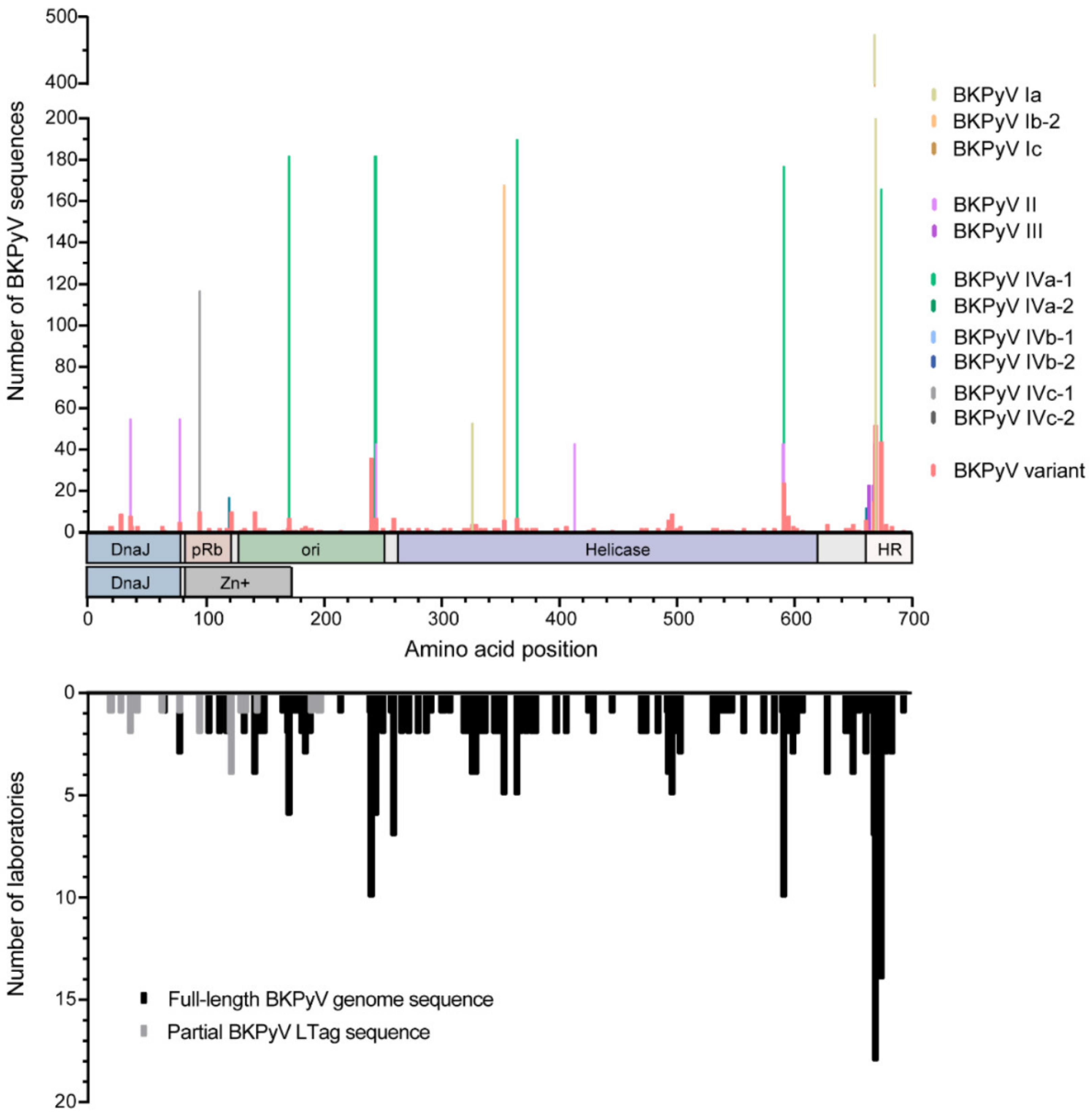
2.1. Assessing Variation in the BKPyV LTag Sequence
2.2. BKPyV Genotypes, BKPyV-Variants and LTag-9mer Variants
2.3. Prediction of HLA-A and -B Binding of BKPyV Immunodominant LTag 9mer T-Cell Epitopes
2.4. BKPyV-Specific CD8 T-Cell Responses In Vitro
2.5. Statistical Analysis
3. Results
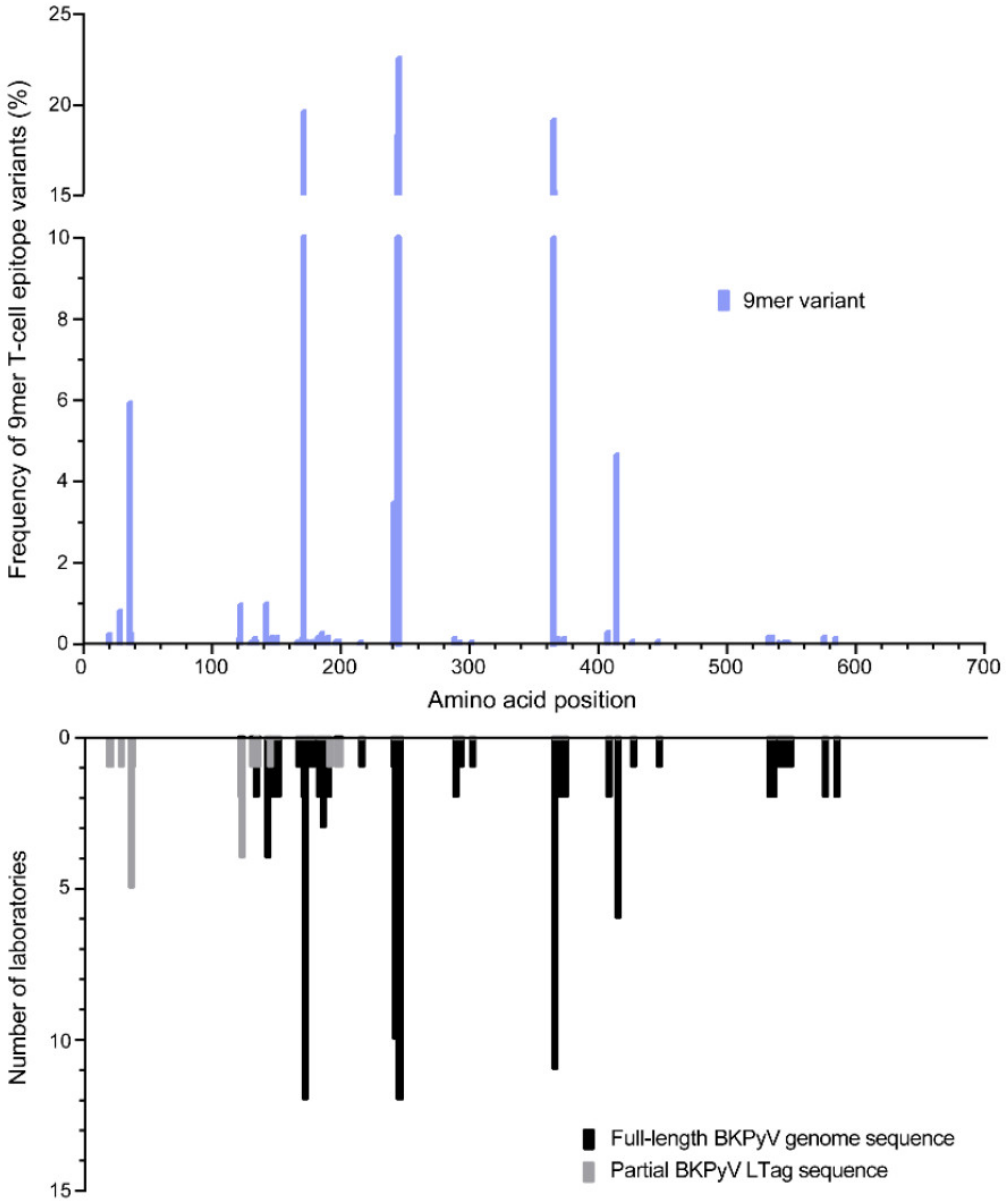
3.1. Identification of BKPyV LTag Variants
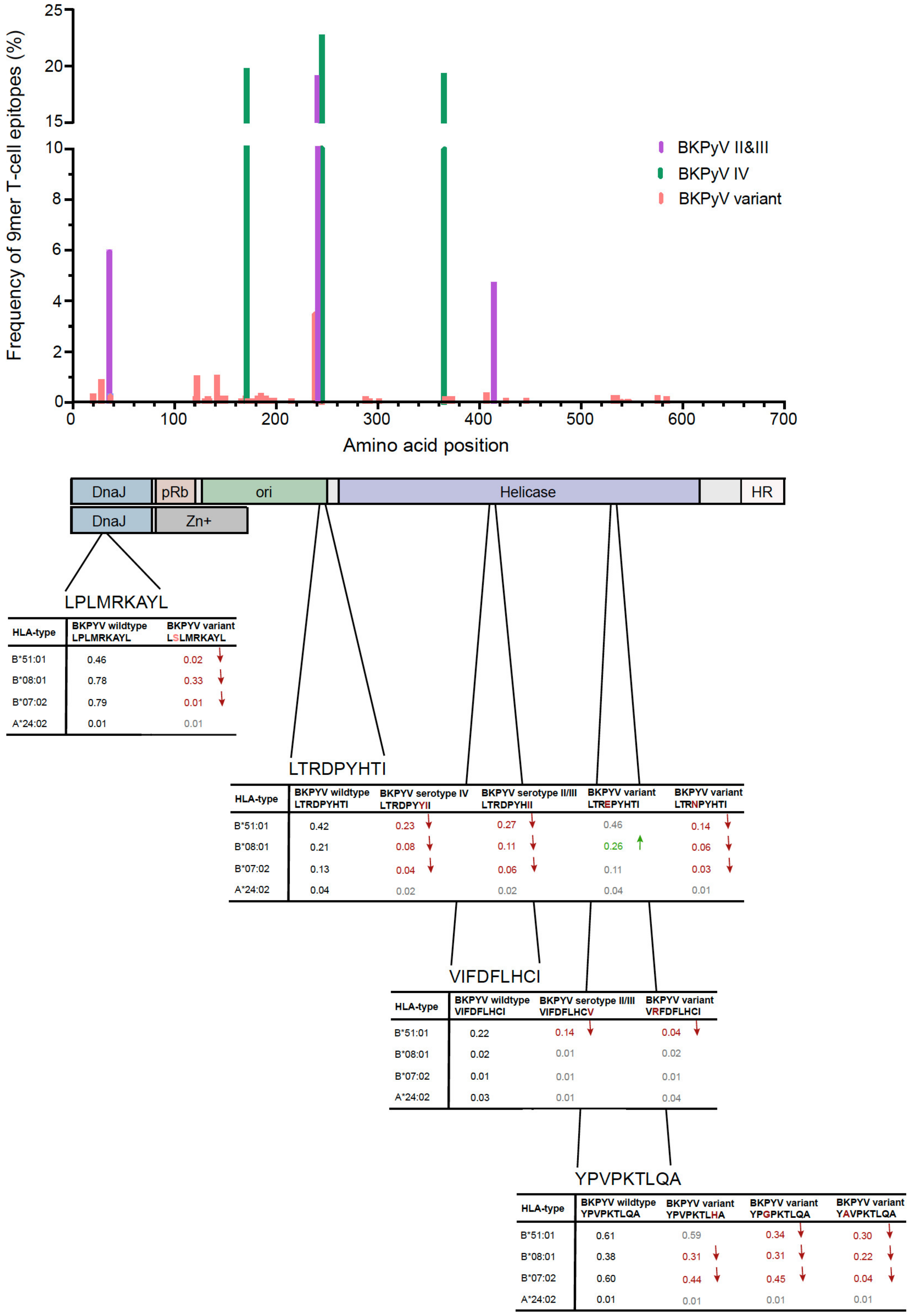
3.2. Identification of Amino Acid Exchanges in Immunodominant LTag 9mer T-Cell Epitopes
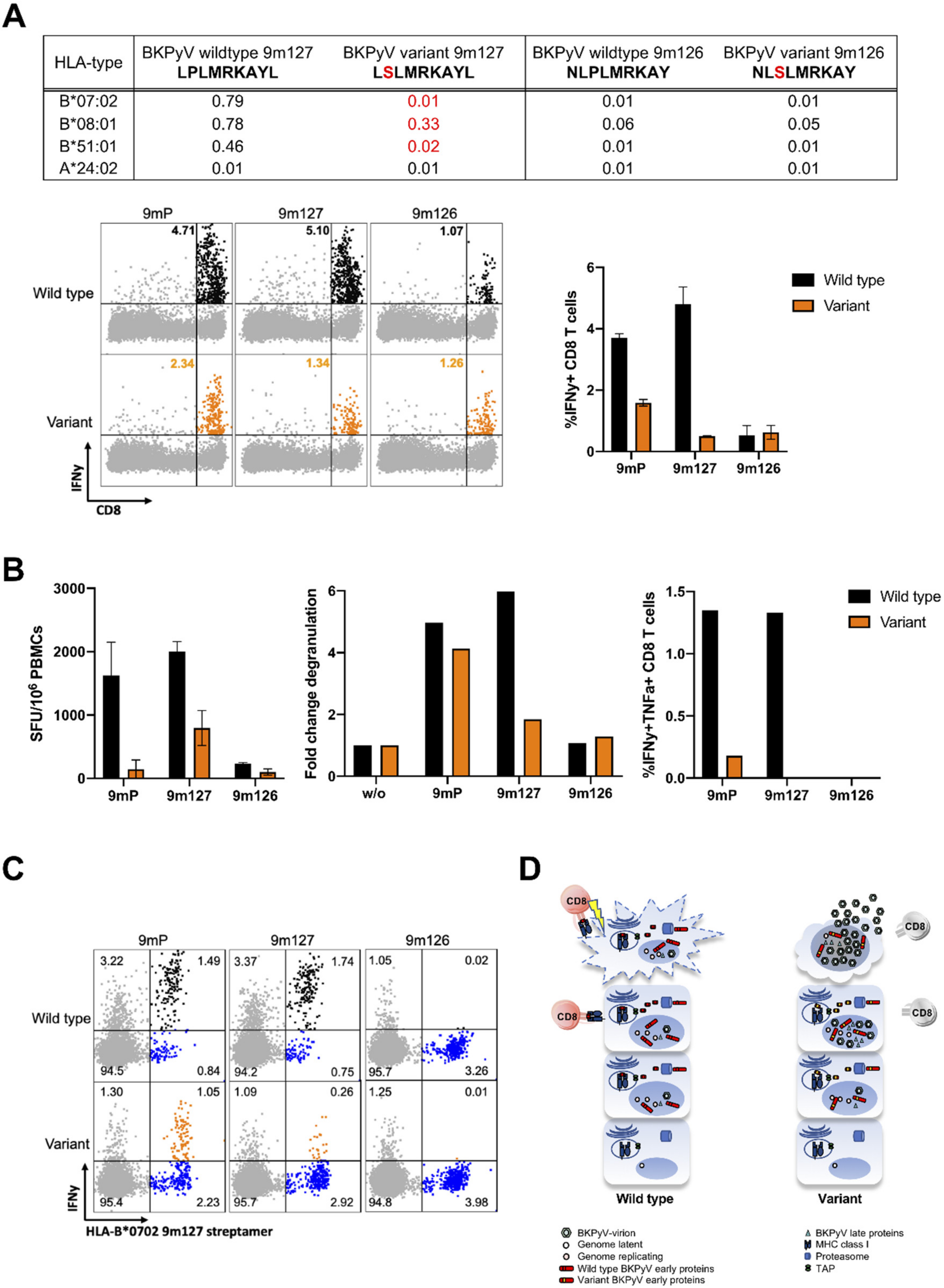
3.3. LTag-9mer Variants Affect HLA-B*07:02, HLA-B*08:01 and HLA-B*51:01 Binding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cesaro, S.; Dalianis, T.; Hanssen Rinaldo, C.; Koskenvuo, M.; Pegoraro, A.; Einsele, H.; Cordonnier, C.; Hirsch, H.H.; ECIL-6 Group. ECIL guidelines for the prevention, diagnosis and treatment of BK polyomavirus-associated haemorrhagic cystitis in haematopoietic stem cell transplant recipients. J. Antimicrob. Chemother. 2018, 73, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Koskenvuo, M.; Dumoulin, A.; Lautenschlager, I.; Auvinen, E.; Mannonen, L.; Anttila, V.J.; Jahnukainen, K.; Saarinen-Pihkala, U.M.; Hirsch, H.H. BK polyomavirus-associated hemorrhagic cystitis among pediatric allogeneic bone marrow transplant recipients: Treatment response and evidence for nosocomial transmission. J. Clin. Virol. 2013, 56, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Imlay, H.; Xie, H.; Leisenring, W.M.; Duke, E.R.; Kimball, L.E.; Huang, M.L.; Pergam, S.A.; Hill, J.A.; Jerome, K.R.; Milano, F.; et al. Presentation of BK polyomavirus-associated hemorrhagic cystitis after allogeneic hematopoietic cell transplantation. Blood Adv. 2020, 4, 617–628. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, H.H.; Knowles, W.; Dickenmann, M.; Passweg, J.; Klimkait, T.; Mihatsch, M.J.; Steiger, J. Prospective study of polyomavirus type BK replication and nephropathy in renal-transplant recipients. N. Engl. J. Med. 2002, 347, 488–496. [Google Scholar] [CrossRef]
- Hirsch, H.H.; Randhawa, P.S. BK polyomavirus in solid organ transplantation-Guidelines from the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transplant. 2019, 33, e13528. [Google Scholar] [CrossRef]
- Kenan, D.J.; Mieczkowski, P.A.; Burger-Calderon, R.; Singh, H.K.; Nickeleit, V. The oncogenic potential of BK-polyomavirus is linked to viral integration into the human genome. J. Pathol. 2015, 237, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, J.C.; Randhawa, P.; Rinaldo, C.H.; Drachenberg, C.B.; Alexiev, B.; Hirsch, H.H. BK Polyomavirus Infection and Renourinary Tumorigenesis. Am. J. Transplant. 2016, 16, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.C.; Ramo, M.; Naegele, K.; Ribi, S.; Wetterauer, C.; Perrina, V.; Quagliata, L.; Vlajnic, T.; Ruiz, C.; Balitzki, B.; et al. Donor-derived, metastatic urothelial cancer after kidney transplantation associated with a potentially oncogenic BK polyomavirus. J. Pathol. 2018, 244, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Meier, R.P.H.; Muller, Y.D.; Dietrich, P.Y.; Tille, J.C.; Nikolaev, S.; Sartori, A.; Labidi-Galy, I.; Ernandez, T.; Kaur, A.; Hirsch, H.H.; et al. Immunologic Clearance of a BK Virus-associated Metastatic Renal Allograft Carcinoma. Transplantation 2020, 2020. [Google Scholar] [CrossRef] [Green Version]
- Ginevri, F.; Comoli, P.; Fontana, I.; Botti, G.; Perfumo, F.; Azzi, A. Prevention of polyomavirus BK-associated nephropathy in pediatric kidney transplantation by prospective monitoring and preemptive immunosuppression reduction. J. Am. Soc. Nephrol. 2005, 7, 2727–2735. [Google Scholar] [CrossRef]
- Binggeli, S.; Egli, A.; Schaub, S.; Binet, I.; Mayr, M.; Steiger, J.; Hirsch, H.H. Polyomavirus BK-Specific Cellular Immune Response to VP1 and Large T-Antigen in Kidney Transplant Recipients. Am. J. Transplant. 2007, 7, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Schachtner, T.; Stein, M.; Babel, N.; Reinke, P. The Loss of BKV-specific Immunity from Pretransplantation to Posttransplantation Identifies Kidney Transplant Recipients at Increased Risk of BKV Replication. Am. J. Transplant. 2015, 15, 2159–2169. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Wilhelm, M.; Wilk, S.; Hirsch, H.H. BK polyomavirus-specific antibody and T-cell responses in kidney transplantation: Update. Curr. Opin. Infect. Dis. 2019, 32, 575–583. [Google Scholar] [CrossRef]
- Kardas, P.; Leboeuf, C.; Hirsch, H.H. Optimizing JC and BK polyomavirus IgG testing for seroepidemiology and patient counseling. J. Clin. Virol. 2015, 71, 28–33. [Google Scholar] [CrossRef]
- Gossai, A.; Waterboer, T.; Nelson, H.H.; Michel, A.; Willhauck-Fleckenstein, M.; Farzan, S.F.; Hoen, A.G.; Christensen, B.C.; Kelsey, K.T.; Marsit, C.J.; et al. Seroepidemiology of Human Polyomaviruses in a US Population. Am. J. Epidemiol. 2016, 183, 61–69. [Google Scholar] [CrossRef]
- Egli, A.; Infanti, L.; Dumoulin, A.; Buser, A.; Samaridis, J.; Stebler, C.; Gosert, R.; Hirsch, H.H. Prevalence of Polyomavirus BK and JC Infection and Replication in 400 Healthy Blood Donors. J. Infect. Dis. 2009, 199, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Manzetti, J.; Weissbach, F.H.; Graf, F.E.; Unterstab, G.; Wernli, M.; Hopfer, H.; Drachenberg, C.B.; Rinaldo, C.H.; Hirsch, H.H. BK Polyomavirus Evades Innate Immune Sensing by Disrupting the Mitochondrial Network and Promotes Mitophagy. iScience 2020, 23, 101257. [Google Scholar] [CrossRef]
- Randhawa, P.S.; Khaleel-Ur-Rehman, K.; Swalsky, P.A.; Vats, A.; Scantlebury, V.; Shapiro, R.; Finkelstein, S. DNA sequencing of viral capsid protein VP-1 region in patients with BK virus interstitial nephritis. Transplantation 2002, 73, 1090–1094. [Google Scholar] [CrossRef]
- Bohl, D.L.; Storch, G.A.; Ryschkewitsch, C.; Gaudreault-Keener, M.; Schnitzler, M.A.; Major, E.O.; Brennan, D.C. Donor origin of BK virus in renal transplantation and role of HLA C7 in susceptibility to sustained BK viremia. Am. J. Transplant. 2005, 5, 2213–2221. [Google Scholar] [CrossRef]
- Schwarz, A.; Linnenweber-Held, S.; Heim, A.; Framke, T.; Haller, H.; Schmitt, C. Viral Origin, Clinical Course, and Renal Outcomes in Patients with BK Virus Infection After Living-Donor Renal Transplantation. Transplantation 2016, 100, 844–853. [Google Scholar] [CrossRef]
- Pastrana, D.V.; Brennan, D.C.; Cuburu, N.; Storch, G.A.; Viscidi, R.P.; Randhawa, P.S.; Buck, C.B. Neutralization serotyping of BK polyomavirus infection in kidney transplant recipients. PLoS Pathog. 2012, 8, e1002650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solis, M.; Velay, A.; Porcher, R.; Domingo-Calap, P.; Soulier, E.; Joly, M.; Meddeb, M.; Kack-Kack, W.; Moulin, B.; Bahram, S.; et al. Neutralizing Antibody-Mediated Response and Risk of BK Virus-Associated Nephropathy. J. Am. Soc. Nephrol. 2018, 29, 326–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Hirsch, H.H.; Kant, J.; Randhawa, P. VP-1 quasispecies in human infection with polyomavirus BK. J. Med. Virol. 2012, 84, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosert, R.; Rinaldo, C.H.; Funk, G.A.; Egli, A.; Ramos, E.; Drachenberg, C.B.; Hirsch, H.H. Polyomavirus BK with rearranged noncoding control region emerge in vivo in renal transplant patients and increase viral replication and cytopathology. J. Exp. Med. 2008, 205, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Peretti, A.; Geoghegan, E.M.; Pastrana, D.V.; Smola, S.; Feld, P.; Sauter, M.; Lohse, S.; Ramesh, M.; Lim, E.S.; Wang, D.; et al. Characterization of BK Polyomaviruses from Kidney Transplant Recipients Suggests a Role for APOBEC3 in Driving In-Host Virus Evolution. Cell Host Microbe 2018, 23, 628–635e7. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.; Adam, C.; Hirsch, H.H.; Janssen, M.W.; Wolf, M.; Dirks, J.; Kardas, P.; Ahlenstiel-Grunow, T.; Pape, L.; Rohrer, T.; et al. BK Polyomavirus-Specific Cellular Immune Responses Are Age-Dependent and Strongly Correlate with Phases of Virus Replication. Am. J. Transplant. 2014, 14, 1334–1345. [Google Scholar] [CrossRef] [Green Version]
- Krautkramer, E.; Klein, T.M.; Sommerer, C.; Schnitzler, P.; Zeier, M. Mutations in the BC-loop of the BKV VP1 region do not influence viral load in renal transplant patients. J. Med. Virol. 2009, 81, 75–81. [Google Scholar] [CrossRef]
- Lorentzen, E.M.; Henriksen, S.; Kaur, A.; Kro, G.B.; Hammarstrom, C.; Hirsch, H.H.; Midtvedt, K.; Rinaldo, C.H. Early fulminant BK polyomavirus-associated nephropathy in two kidney transplant patients with low neutralizing antibody titers receiving allografts from the same donor. Virol. J. 2020, 17, 5. [Google Scholar] [CrossRef]
- Leuzinger, K.; Naegele, K.; Schaub, S.; Hirsch, H.H. Quantification of plasma BK polyomavirus loads is affected by sequence variability, amplicon length, and non-encapsidated viral DNA genome fragments. J. Clin. Virol. 2019, 121, 104210. [Google Scholar] [CrossRef]
- Naegele, K.; Lautenschlager, I.; Gosert, R.; Loginov, R.; Bir, K.; Helantera, I.; Schaub, S.; Khanna, N.; Hirsch, H.H. Cytomegalovirus sequence variability, amplicon length, and DNase-sensitive non-encapsidated genomes are obstacles to standardization and commutability of plasma viral load results. J. Clin. Virol. 2018, 104, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Pang, X.L.; Mabilangan, C.; Preiksaitis, J.K. Determination of the Biological Form of Human Cytomegalovirus DNA in the Plasma of Solid-Organ Transplant Recipients. J. Infect. Dis. 2017, 215, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Sester, M.; Leboeuf, C.; Schmidt, T.; Hirsch, H.H. The “ABC” of Virus-Specific T Cell Immunity in Solid Organ Transplantation. Am. J. Transplant. 2016, 16, 1697–1706. [Google Scholar] [CrossRef]
- Ahlenstiel-Grunow, T.; Sester, M.; Sester, U.; Hirsch, H.H.; Pape, L. BK Polyomavirus-specific T Cells as a Diagnostic and Prognostic Marker for BK Polyomavirus Infections After Pediatric Kidney Transplantation. Transplantation 2020, 104, 2393–2402. [Google Scholar] [CrossRef]
- Cioni, M.; Leboeuf, C.; Comoli, P.; Ginevri, F.; Hirsch, H.H. Characterization of Immunodominant BK Polyomavirus 9mer Epitope T Cell Responses. Am. J. Transplant. 2016, 16, 1193–1206. [Google Scholar] [CrossRef] [Green Version]
- Van Aalderen, M.C.; Remmerswaal, E.B.; Heutinck, K.M.; Ten Brinke, A.; Pircher, H.; van Lier, R.A.; Ten Berge, I.J. Phenotypic and Functional Characterization of Circulating Polyomavirus BK VP1-Specific CD8+ T Cells in Healthy Adults. J. Virol. 2013, 87, 10263–10272. [Google Scholar] [CrossRef] [Green Version]
- Ramaswami, B.; Popescu, I.; Macedo, C.; Metes, D.; Bueno, M.; Zeevi, A.; Shapiro, R.; Viscidi, R.; Randhawa, P.S. HLA-A01-, -A03-, and -A024-binding nanomeric epitopes in polyomavirus BK large T antigen. Hum. Immunol. 2009, 70, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Leboeuf, C.; Wilk, S.; Achermann, R.; Binet, I.; Golshayan, D.; Hadaya, K.; Hirzel, C.; Hoffmann, M.; Huynh-Do, U.; Koller, M.T.; et al. BK Polyomavirus-Specific 9mer CD8 T Cell Responses Correlate with Clearance of BK Viremia in Kidney Transplant Recipients: First Report from the Swiss Transplant Cohort Study. Am. J. Transplant. 2017, 17, 2591–2600. [Google Scholar] [CrossRef]
- Willhelm, M.; Wilk, S.; Kaur, A.; Hirsch, H.H.; Swiss Transplant Cohort, S. Can HLA-B51 Protect Against BKPyV-DNAemia? Transplantation 2019, 103, e384–e385. [Google Scholar] [CrossRef]
- Wunderink, H.F.; Haasnoot, G.W.; de Brouwer, C.S.; van Zwet, E.W.; Kroes, A.C.M.; de Fijter, J.W.; Rotmans, J.I.; Claas, F.H.J.; Feltkamp, M.C.W. Reduced Risk of BK Polyomavirus Infection in HLA-B51-positive Kidney Transplant Recipients. Transplantation 2019, 103, 604–612. [Google Scholar] [CrossRef]
- Wilhelm, M.; Kaur, A.; Wernli, M.; Hirsch, H.H. BK Polyomavirus (BKPyV)-specific CD8 T-cell expansion in vitro using 27mer peptide antigens for developing adoptive T-cell transfer and vaccination. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Jin, L.; Gibson, P.E.; Booth, J.C.; Clewley, J.P. Genomic typing of BK virus in clinical specimens by direct sequencing of polymerase chain reaction products. J. Med. Virol. 1993, 41, 11–17. [Google Scholar] [CrossRef]
- Nishimoto, Y.; Zheng, H.Y.; Zhong, S.; Ikegaya, H.; Chen, Q.; Sugimoto, C.; Kitamura, T.; Yogo, Y. An Asian origin for subtype IV BK virus based on phylogenetic analysis. J. Mol. Evol. 2007, 65, 103–111. [Google Scholar] [CrossRef]
- Torres, C. Evolution and molecular epidemiology of polyomaviruses. Infect. Genet. Evol. 2020, 79, 104150. [Google Scholar] [CrossRef]
- DeCaprio, J.A.; Garcea, R.L. A cornucopia of human polyomaviruses. Nat. Rev. Microbiol. 2013, 11, 264–276. [Google Scholar] [CrossRef]
- Ahlenstiel-Grunow, T.; Pape, L. Immunosuppression, BK polyomavirus infections, and BK polyomavirus-specific T cells after pediatric kidney transplantation. Pediatr. Nephrol. 2020, 35, 625–631. [Google Scholar] [CrossRef]
- Comoli, P.; Cioni, M.; Basso, S.; Gagliardone, C.; Potenza, L.; Verrina, E.; Luppi, M.; Zecca, M.; Ghiggeri, G.M.; Ginevri, F. Immunity to Polyomavirus BK Infection: Immune Monitoring to Regulate the Balance between Risk of BKV Nephropathy and Induction of Alloimmunity. Clin. Dev. Immunol. 2013, 2013, 256923. [Google Scholar] [CrossRef]
- Ikegaya, H.; Saukko, P.J.; Tertti, R.; Metsarinne, K.P.; Carr, M.J.; Crowley, B.; Sakurada, K.; Zheng, H.Y.; Kitamura, T.; Yogo, Y. Identification of a genomic subgroup of BK polyomavirus spread in European populations. J. Gen. Virol. 2006, 87 Pt 11, 3201–3208. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Nishimoto, Y.; Chen, Q.; Hasegawa, M.; Zhong, S.; Ikegaya, H.; Ohno, N.; Sugimoto, C.; Takasaka, T.; Kitamura, T.; et al. Relationships between BK virus lineages and human populations. Microbes Infect. 2007, 9, 204–213. [Google Scholar] [CrossRef]
- Abend, J.R.; Changala, M.; Sathe, A.; Casey, F.; Kistler, A.; Chandran, S.; Howard, A.; Wojciechowski, D. Correlation of BK Virus Neutralizing Serostatus With the Incidence of BK Viremia in Kidney Transplant Recipients. Transplantation 2017, 101, 1495–1505. [Google Scholar] [CrossRef]
- McIlroy, D.; Honemann, M.; Nguyen, N.K.; Barbier, P.; Peltier, C.; Rodallec, A.; Halary, F.; Przyrowski, E.; Liebert, U.; Hourmant, M.; et al. Persistent BK Polyomavirus Viruria is Associated with Accumulation of VP1 Mutations and Neutralization Escape. Viruses 2020, 12, 824. [Google Scholar] [CrossRef]
- Ginevri, F.; Azzi, A.; Hirsch, H.H.; Basso, S.; Fontana, I.; Cioni, M.; Bodaghi, S.; Salotti, V.; Rinieri, A.; Botti, G.; et al. Prospective monitoring of polyomavirus BK replication and impact of pre-emptive intervention in pediatric kidney recipients. Am. J. Transplant. 2007, 7, 2727–2735. [Google Scholar] [CrossRef] [PubMed]
- Schachtner, T.; Muller, K.; Stein, M.; Diezemann, C.; Sefrin, A.; Babel, N.; Reinke, P. BK virus-specific immunity kinetics: A predictor of recovery from polyomavirus BK-associated nephropathy. Am. J. Transplant. 2011, 11, 2443–2452. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequency of LTag 9mer Variants (%) | Number of LTag 9mer Variants | Number of LTag 9mer Variants with Changed HLA Binding 1 | |||
|---|---|---|---|---|---|
| ↓ | ↑ | ↓↑ | Total | ||
| <1% | 127 | 19 | 15 | 2 | 36 |
| 1–<5% | 14 | 4 | 1 | 1 | 6 |
| 5–<10% | 10 | 2 | 1 | - | 3 |
| 10–<25.0% | 8 | 2 | 1 | - | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leuzinger, K.; Kaur, A.; Wilhelm, M.; Hirsch, H.H. Variations in BK Polyomavirus Immunodominant Large Tumor Antigen-Specific 9mer CD8 T-Cell Epitopes Predict Altered HLA-Presentation and Immune Failure. Viruses 2020, 12, 1476. https://doi.org/10.3390/v12121476
Leuzinger K, Kaur A, Wilhelm M, Hirsch HH. Variations in BK Polyomavirus Immunodominant Large Tumor Antigen-Specific 9mer CD8 T-Cell Epitopes Predict Altered HLA-Presentation and Immune Failure. Viruses. 2020; 12(12):1476. https://doi.org/10.3390/v12121476
Chicago/Turabian StyleLeuzinger, Karoline, Amandeep Kaur, Maud Wilhelm, and Hans H. Hirsch. 2020. "Variations in BK Polyomavirus Immunodominant Large Tumor Antigen-Specific 9mer CD8 T-Cell Epitopes Predict Altered HLA-Presentation and Immune Failure" Viruses 12, no. 12: 1476. https://doi.org/10.3390/v12121476