Detection of Recombinant Hare Myxoma Virus in Wild Rabbits (Oryctolagus cuniculus algirus)

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample, Necropsy and Histopathology
2.2. Parasitological and Bacteriological Examination
2.3. Virological and Serological Examinations
2.4. Sequencing Analysis
2.5. Isolation
2.6. Electron Microscopy
3. Results
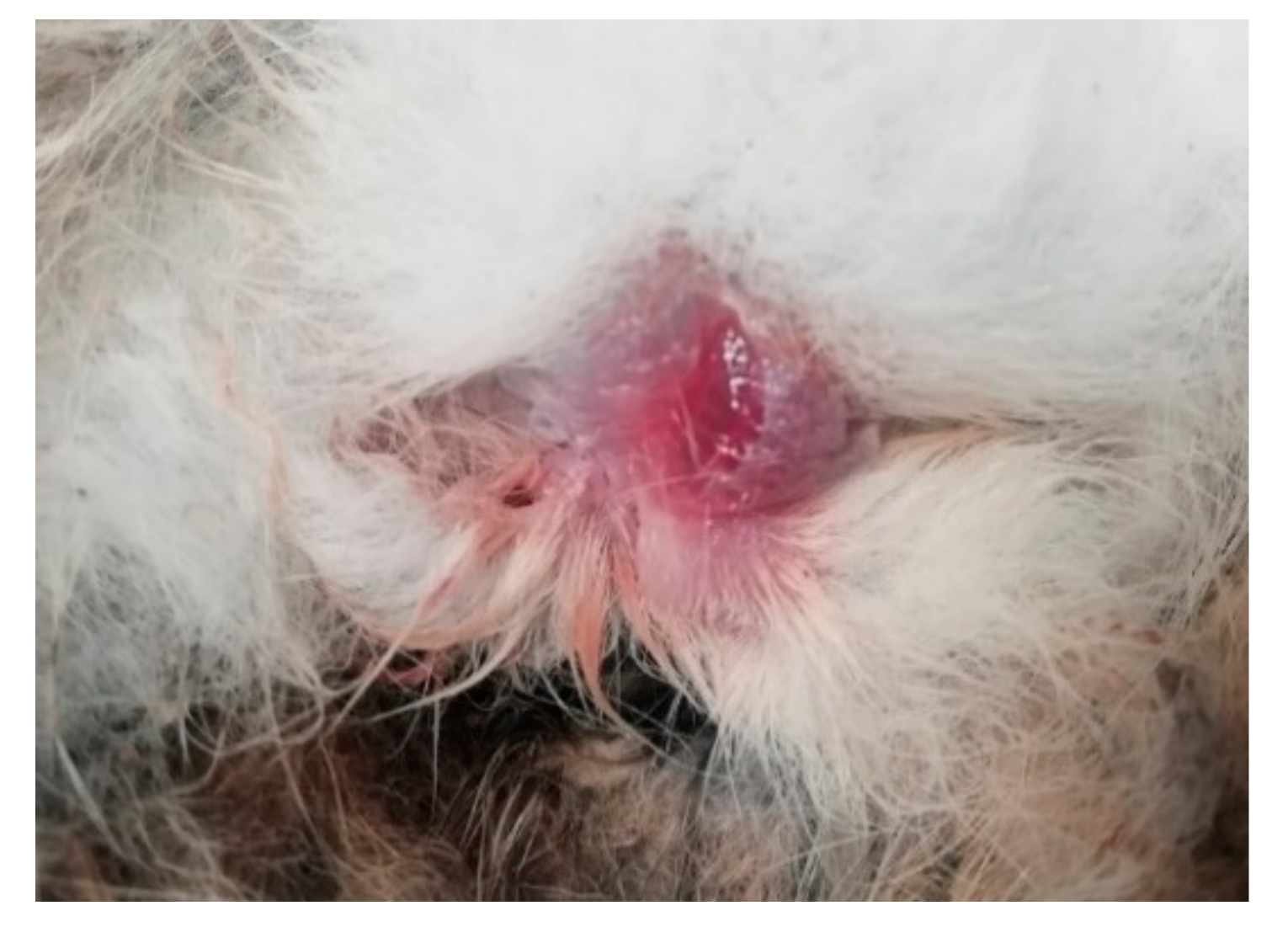
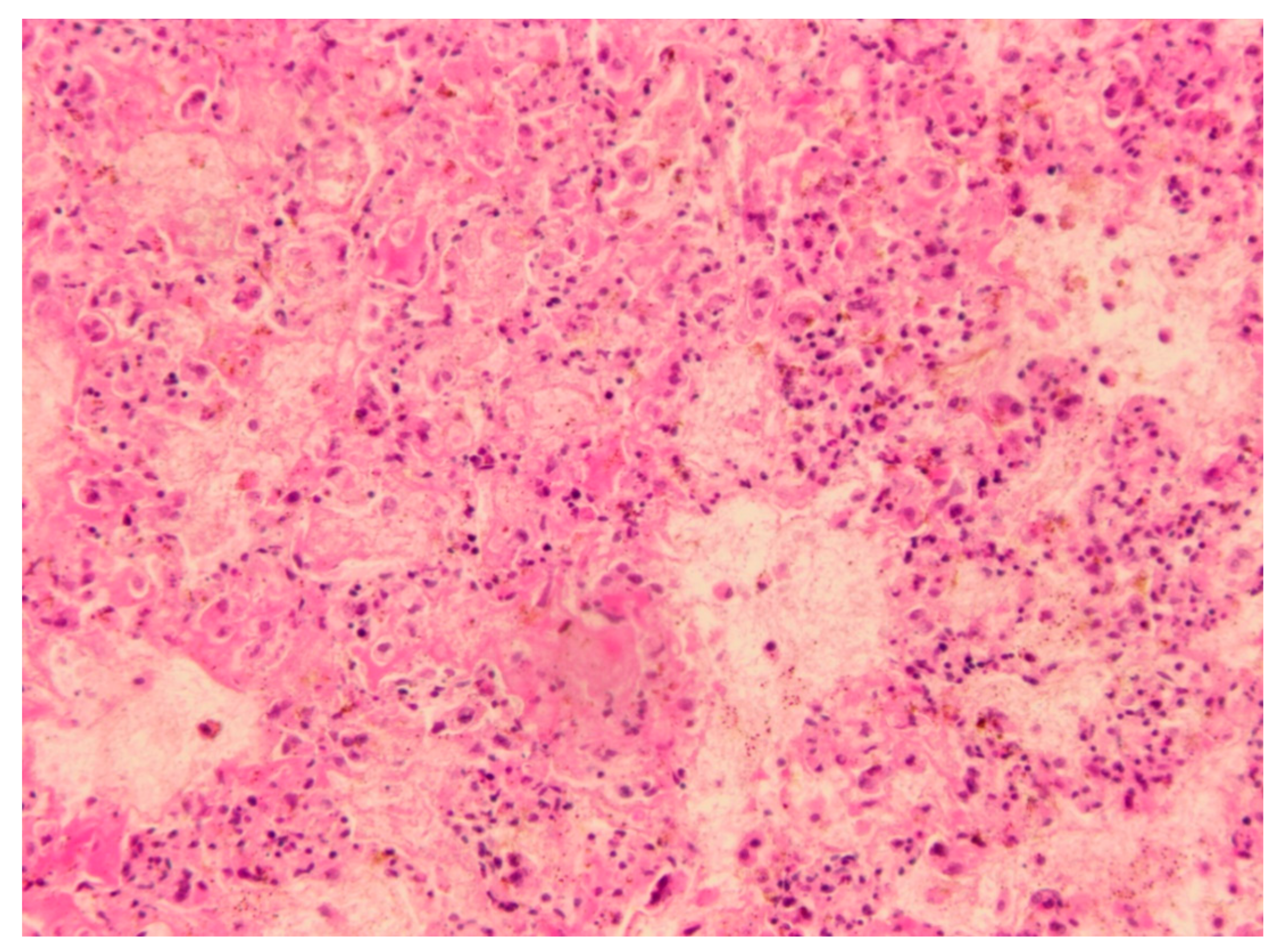
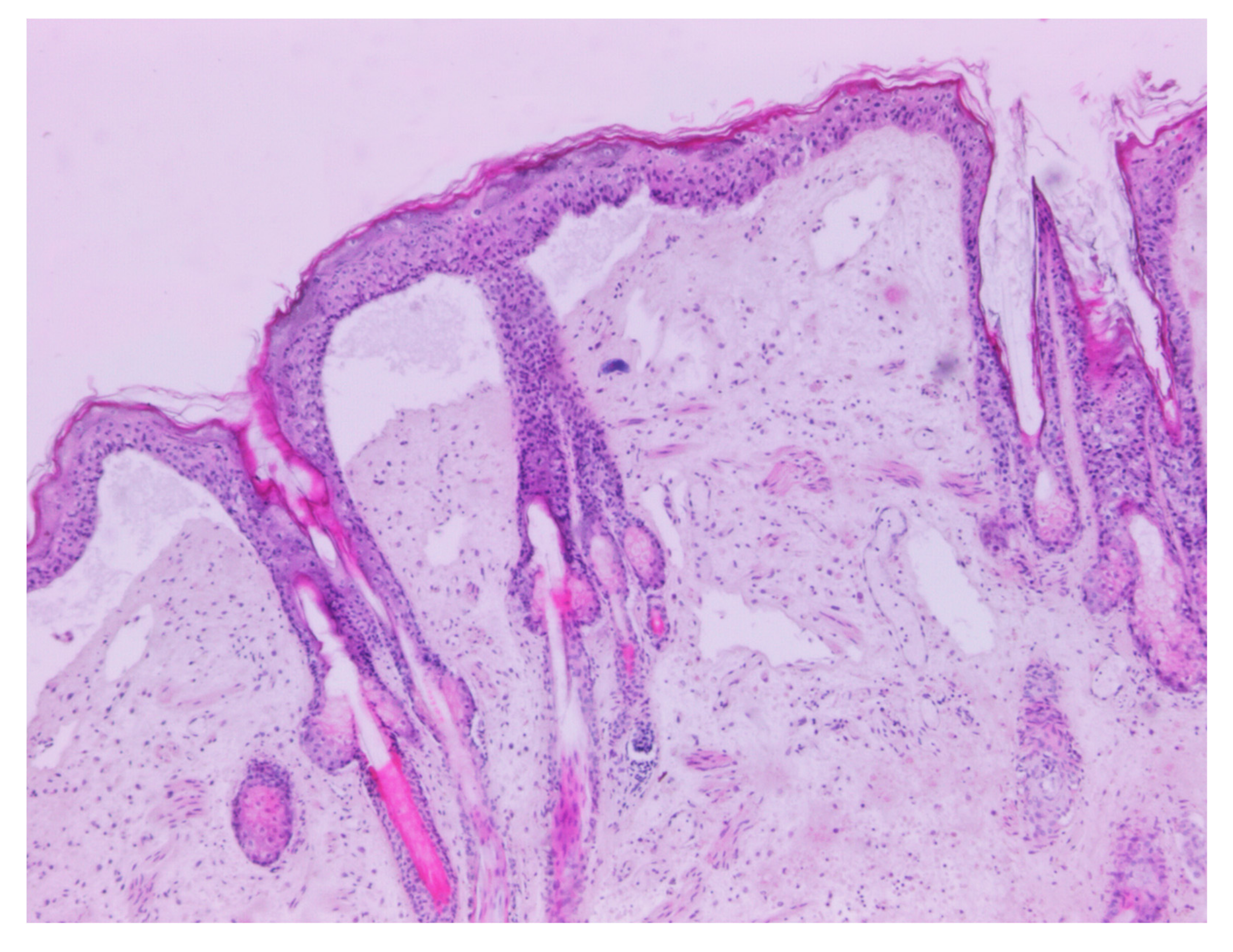
3.1. Necropsy and Histopathology
3.2. Virological, Bacteriological and Parasitological Results
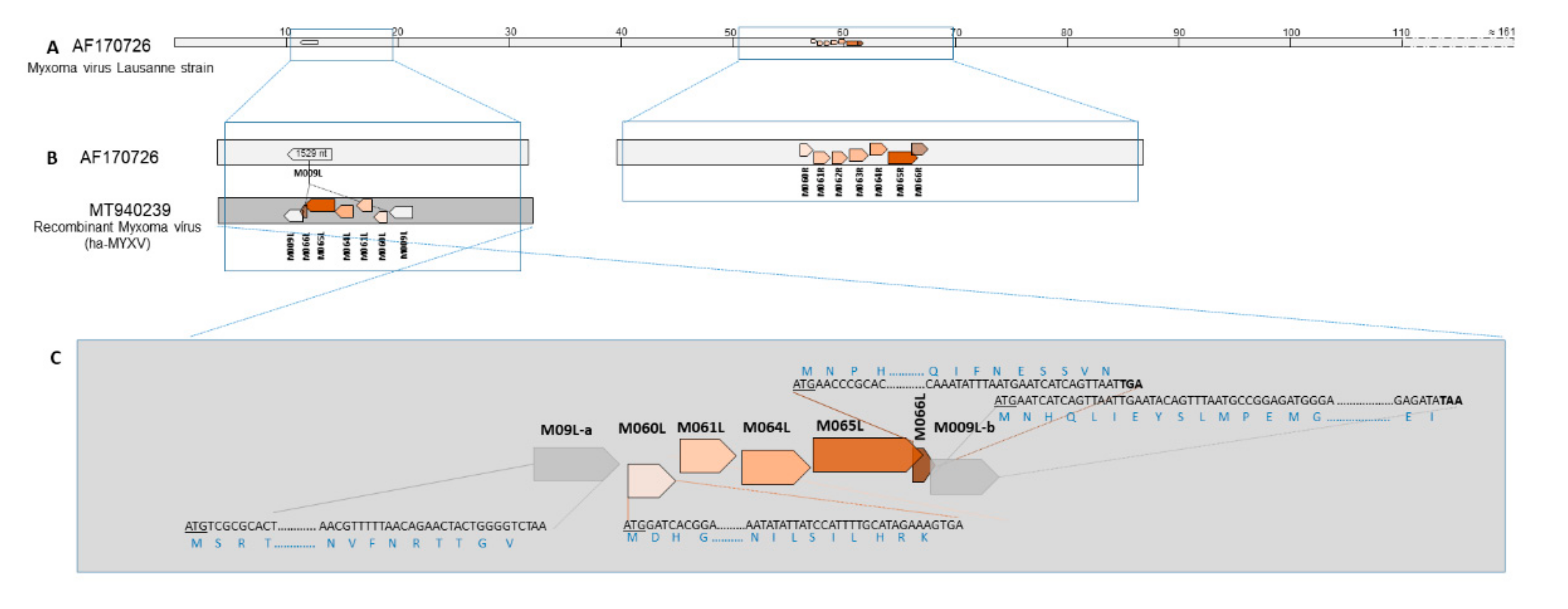
3.3. Molecular Characterisation
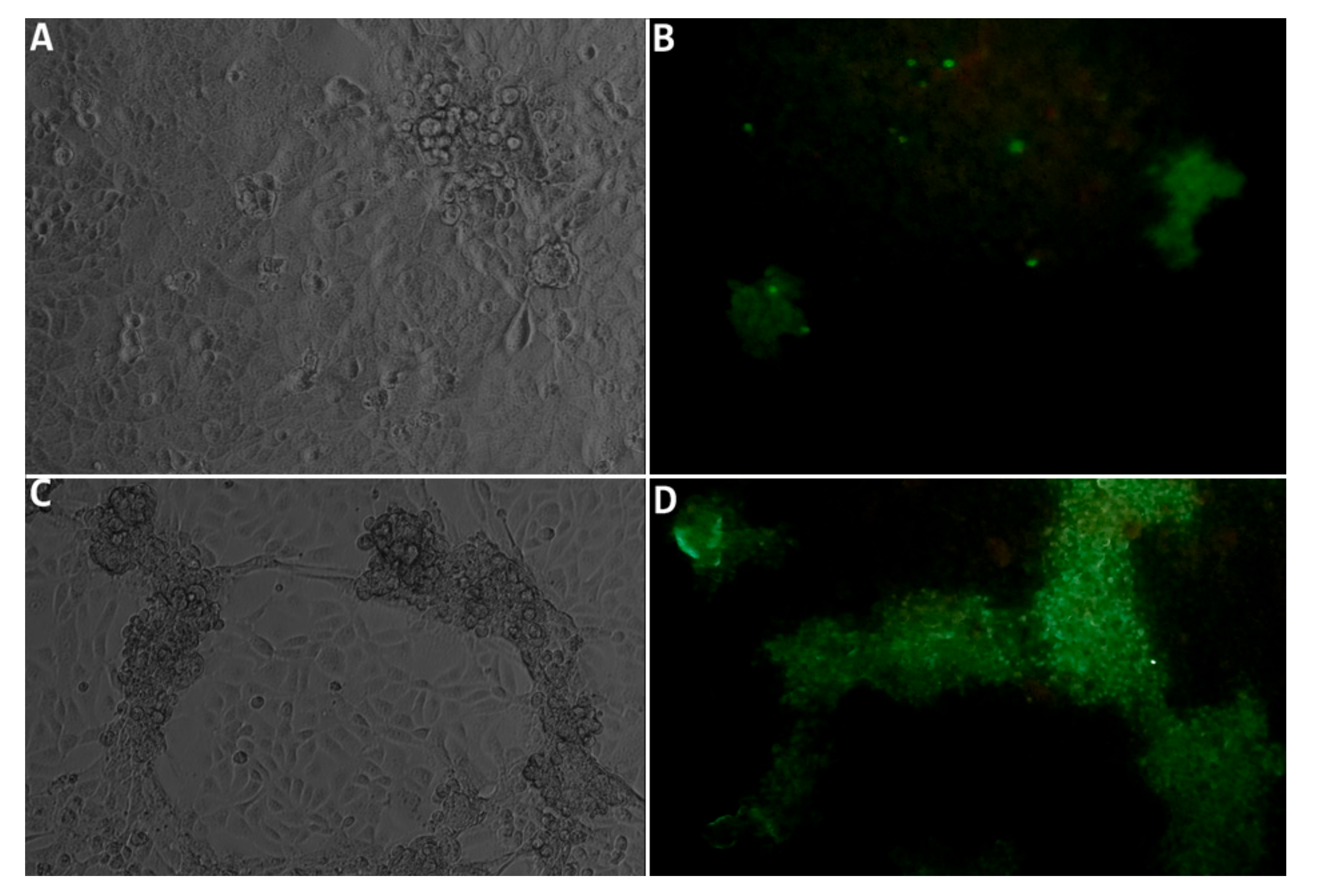
3.4. Isolation of the Virus in Cell Culture
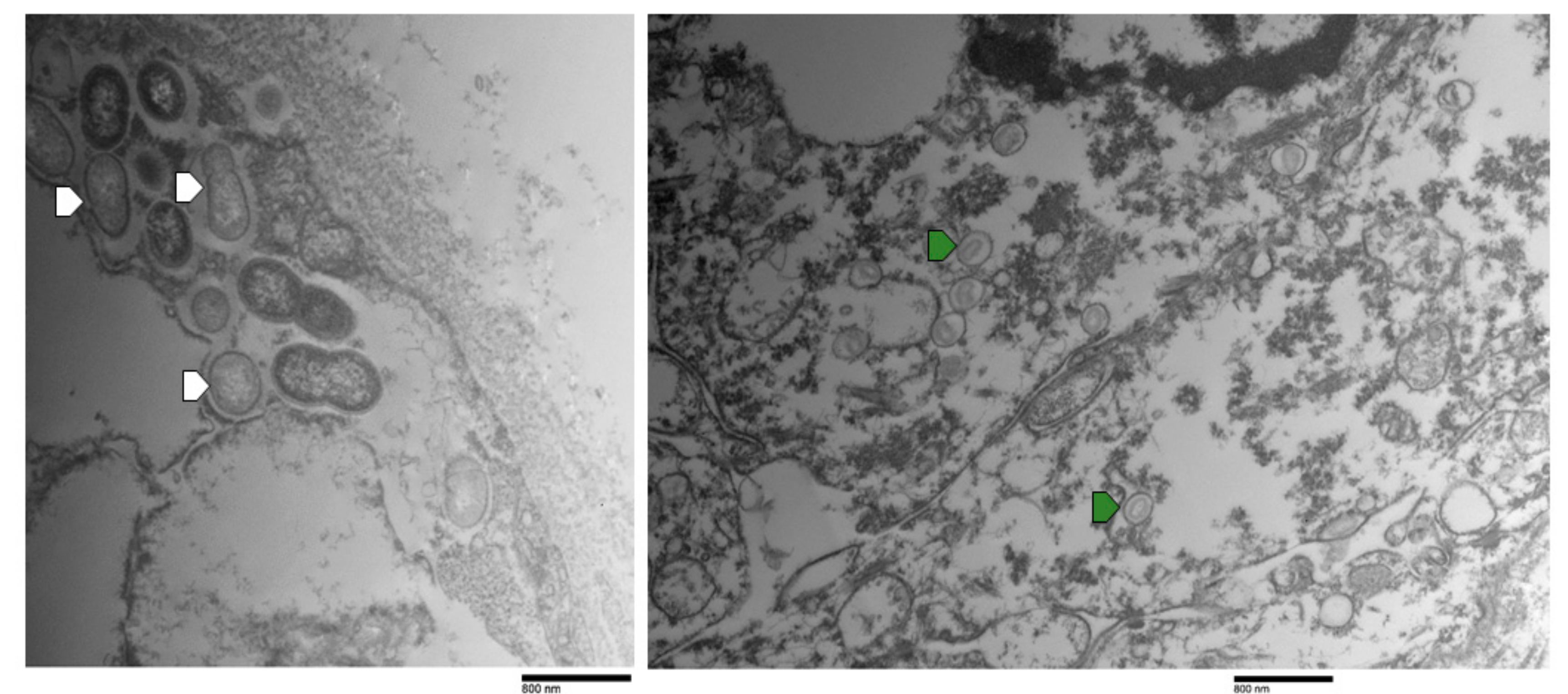
3.5. Transmission Electron Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Declaration
Conflicts of Interest
References
- Delibes-Mateos, M.; Delibes, M.; Ferreras, P.; Villafuerte, R. Key role of European rabbits in the conservation of the western Mediterranean Basin hotspot. Conserv. Biol. 2008, 22, 1106–1117. [Google Scholar] [CrossRef]
- Staniforth, R.J.; Cavers, P.B. The importance of cottontail rabbits in the dispersal of Polygonum spp. J. Appl. Ecol. 1977, 14, 261–268. [Google Scholar] [CrossRef]
- Carvalho, C.L.; Abade dos Santos, F.A.; Fagulha, T.; Carvalho, P.; Mendonça, P.; Monteiro, M.; Duarte, M.D. Myxoma virus and rabbit haemorrhagic disease virus 2 coinfection in a European wild rabbit (Oryctolagus cuniculus algirus), Portugal. Vet. Rec. Case Rep. 2020, 18–21. [Google Scholar] [CrossRef]
- Murphy, F.A.; Fauquet, C.M.; Bishop, D.H.L.; Ghabrial, S.A.; Jarvis, A.W.; Martelli, G.P.; Mayo, M.A.; Summers, M. Virus Taxonomy: Classification and Nomenclature of Viruses; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Sanarelli, G. Das myxomatogene Virus. Beitrag zum Studium der Krankheitserreger ausserhalb der Sichtbaren. (Vorliiufige Mitteilung.). Zentralbl. Bakteriol. Parasitenkd. Infektionskrankh. Abt. I. 1898, 23, 865–873. [Google Scholar]
- Rivers, T.M. Infectious myxomatosis of rabbits. J. Exp. Med. 1930, 51, 965–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlier, D.; Cassart, D.; Coignoul, F.; Vindevogel, H. Experimental Infection of Specific Pathogen-free New Zealand White Rabbits with Five Strains of Amyxomatous Myxoma Virus. J. Comp. Pathol. 1999, 121, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Moss, B. Fields virology. In Field’s Virology; Knipe, D.M., Howley, P.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 2849–2883. [Google Scholar]
- Bertagnoli, S.; Marchandeau, S. Myxomatosis. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 549–556. [Google Scholar]
- Arthur, C.P.; Louzis, C. A review of myxomatosis among rabbits in France. Rev. Sci. Tech. Off. Int. Epiz. 1988, 7, 959–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, A.; Lawrence, K.; Everest, D.; Dastjerdi, A.; Finnegan, C.; Steinbach, F. Confirmation of myxomatosis in a European brown hare in Great Britain. Vet. Rec. 2014, 175, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.J. Myxomatosis in the common hare. Ir. Vet. J. 1955, 9, 268. [Google Scholar]
- Pinto, A.; de Matos, A.L.; Abrantes, M.; Kraberger, S.; Risalde, M.A.; Gort, C.; Mcfadden, G.; Varsani, A.; Esteves, P.J. Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses 2019, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Bocanegra, I.G.; Camacho-Sillero, L.; Risalde, M.A.; Dalton, K.; Caballero-Gómez, J.; Aguero, M.; Zorrila, I.; Gómez-Guillamón, F. First outbreak of myxomatosis in Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 1–5. [Google Scholar] [CrossRef]
- Dalton, K.P.; Martín, J.M.; Nicieza, I.; Podadera, A.; Llano, D.; Casais, R.; Gimenez, S.; Badiola, I.; Agüero, M.; Duran, M.; et al. Myxoma virus jumps species to the Iberian hare. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.; Abade dos Santos, F.A.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Duarte, M.D. First cases of myxomatosis in Iberian hares (Lepus granatensis) in Portugal. Vet. Rec. Case Rep. 2020, 8. [Google Scholar] [CrossRef]
- Duarte, M.D.; Barros, S.C.; Henriques, A.M.; Fagulha, M.T.; Ramos, F.; Luís, T.; Fevereiro, M. Development and validation of a real time PCR for the detection of myxoma virus based on the diploid gene M000.5L/R. J. Virol. Methods 2013, 196, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.D.; Carvalho, C.L.; Barros, S.C.; Henriques, A.M.; Ramos, F.; Fagulha, T.; Luís, T.; Duarte, E.L.; Fevereiro, M. A real time Taqman RT-PCR for the detection of rabbit hemorrhagic disease virus 2 (RHDV2). J. Virol. Methods 2015, 219, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Abade dos Santos, F.A.; Monteiro, M.; Pinto, A.; Carvalho, C.L.; Peleteiro, M.C.; Carvalho, P.; Mendonça, P.; Carvalho, T.; Duarte, M.D. First description of a herpesvirus infection in genus Lepus. BioRxiv Prepr. 2020. [Google Scholar] [CrossRef]
- Villafuerte, R.; Delibes-Mateos, M. Oryctolagus cuniculus (errata version published in 2020). In IUCN Red List Threatened Species 2019: e.T41291A170619657; International Union for Conservation of Nature and Natural Resources: Cmbridge, UK, 2019. [Google Scholar] [CrossRef]
- Abade dos Santos, F.A.; Santos, N.; Carvalho, C.; Martinez, M.; Gortazar, C.; Garc, I.; Duarte, M.; Alves, P. Serology unveils decades-long contact of the Iberian hare, Lepus granatensis, with myxoma or antigenically-related virus. Authorea 2020. [Google Scholar] [CrossRef]
- Duarte, M.D.; Carvalho, C.L.; Abade dos Santos, F.A.; Gomes, J.; Alves, P.C.; Esteves, P.J.; Abrantes, J.; Lopes, A.M.; P., M.; Serronha, A.; et al. +Coelho: Avaliação Ecossanitária das Populações Naturais de Coelho Bravo Visando o Controlo da Doença Hemorrágica Viral. Relatório de Atividades na Época Venatória 2018, 1, 1–156. [Google Scholar]
- Kerr, P.J.; Liu, J.; Cattadori, I.; Ghedin, E.; Read, A.F.; Holmes, E.C. Myxoma virus and the leporipoxviruses: An evolutionary paradigm. Viruses 2015, 7, 1020–1061. [Google Scholar] [CrossRef]
- Hipra, L. Serological response to myxomatosis vaccination by different inoculation systems on farm rabbits. World Rabbit Sci. 2003, 11, 145–156. [Google Scholar]
- Best, S.M.; Kerr, P.J. Coevolution of host and virus: The pathogenesis of virulent and attenuated strains of myxoma virus in resistant and susceptible European rabbits. Virology 2000, 267, 36–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5’-3’) | Position in MK340973 (nt) | Reference |
|---|---|---|---|
| 9B (forward) | CGCAGGTCCACGTATAAACC | 11458–11477 and 153103–153084 | [15] |
| 9A (reverse) | CGAACGTATCATTAGACAATG | 16060–16040 | |
| 9E (forward) | CTTCGTCTACGCCTCCTACG | 12116–12135 | |
| 9F (reverse) | GCGTCGTTGTGGTCAGACAGAG | 15256–15243 | |
| 305R (reverse) | AACCCGCACAACGTAAAGTACC | 12420–12399 | This manuscript |
| 448F (forward) | GTCATATTCCTGATTTGGGTAATC | 12563–12587 | |
| 796R (reverse) | AGGAGGAAAAGAACCTATGACAC | 12911–12889 | |
| 1003F (forward) | GTGTGTACCTGGTGCAGAACC | 13118–13138 | |
| 1302R (reverse) | TGAAGATGATAATGATGATGAATATCG | 13417–13391 | |
| 1467F (forward) | TTCATCGTTTATGGGAAAATCTATG | 13582–13606 | |
| 1819R (reverse) | GAGGGGACAGTTATGGATGTAC | 13934–13913 | |
| 2028F (forward) | AAGATGCGTCTGTGTAACAATCC | 14143–14165 | |
| 2325R (reverse) | AACAATGTATACACTCATGACAGTAC | 14440–14415 | |
| 2458F (forward) | ATGGCCATCGTAAGTTGCCATG | 14573–14594 | |
| 2847R (reverse) | CAGAGTACTTAGATTTTCTGCTAG | 14962–14939 | |
| 2954F (forward) | ATCCATTGTTCGTCAGTAGATCG | 15069–15091 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abade dos Santos, F.A.; Carvalho, C.L.; Pinto, A.; Rai, R.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Peleteiro, M.C.; Parra, F.; Duarte, M.D. Detection of Recombinant Hare Myxoma Virus in Wild Rabbits (Oryctolagus cuniculus algirus). Viruses 2020, 12, 1127. https://doi.org/10.3390/v12101127
Abade dos Santos FA, Carvalho CL, Pinto A, Rai R, Monteiro M, Carvalho P, Mendonça P, Peleteiro MC, Parra F, Duarte MD. Detection of Recombinant Hare Myxoma Virus in Wild Rabbits (Oryctolagus cuniculus algirus). Viruses. 2020; 12(10):1127. https://doi.org/10.3390/v12101127
Chicago/Turabian StyleAbade dos Santos, Fábio A., Carina L. Carvalho, Andreia Pinto, Ranjit Rai, Madalena Monteiro, Paulo Carvalho, Paula Mendonça, Maria C. Peleteiro, Francisco Parra, and Margarida D. Duarte. 2020. "Detection of Recombinant Hare Myxoma Virus in Wild Rabbits (Oryctolagus cuniculus algirus)" Viruses 12, no. 10: 1127. https://doi.org/10.3390/v12101127