Systematic Identification of Host Immune Key Factors Influencing Viral Infection in PBL of ALV-J Infected SPF Chicken

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus and Experimental Animals
2.3. ALV-J Shedding, ALV-J Viremia, ALV-J Antibody Detection
2.4. Lymphocyte Isolation
2.5. Flow Cytometry
2.6. Expression Analysis of Immune-Related Gene by qRT-PCR
2.7. Statistical Analyses
3. Results
3.1. Dynamic Detection of ALV-J Shedding, ALV-J Viremia, and ALV-J Antibody
3.2. Dynamic Detection of T Lymphocyte Percentage
3.3. Kinetic Detection of Immune-related Genes in PBL
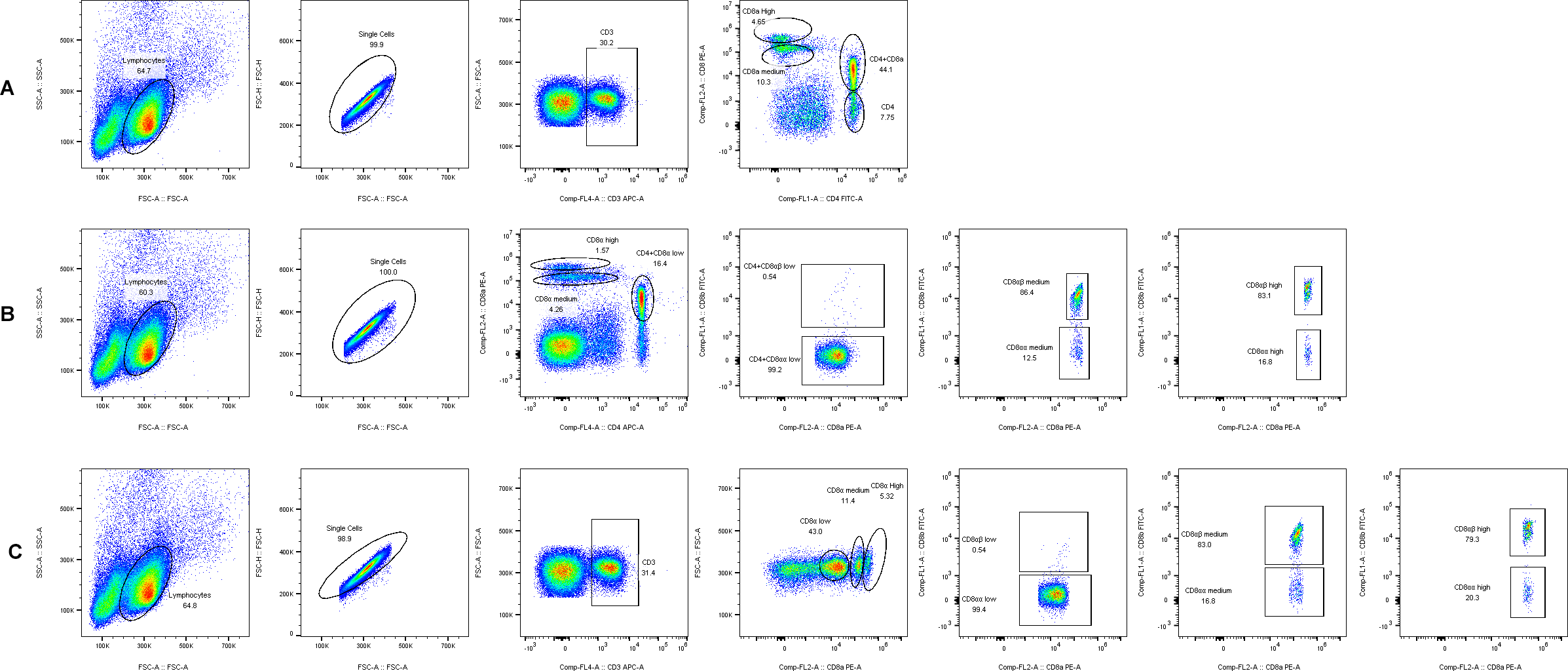
3.4. CD8+ T Cell Phenotype Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Payne, L.N.; Nair, V. The long view: 40 years of avian leukosis research. Avian Pathol. 2012, 41, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.M.; Feng, M.; Xie, T.T.; Li, Y.F.; Zhang, X.Q. Fluctuations in luteinizing hormone, follicle stimulating hormone, and progesterone might affect the disappearance of avian leukosis virus subgroup J viremia in chickens with intermittent viremia. Poult. Sci. 2019, 98, 3533–3538. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.M.; Cai, D.J.; Zhao, X.N.; Cheng, Z.Q.; Guo, H.J.; Qi, C.H.; Liu, J.Z.; Xu, R.X.; Zhao, P.; Cui, Z.Z. Liposomes containing recombinant gp85 protein vaccine against ALV-J in chickens. Vaccine 2014, 32, 2452–2456. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.W.; Li, H.M.; Cheng, Z.Q.; Zhao, P.; Liu, J.X.; Cui, Z.Z.; Liu, H.G.; Jing, W.F.; Guo, H.J. Maternal antibody induced by recombinant gp85 protein vaccine adjuvanted with CpG-ODN protects against ALV-J early infection in chickens. Vaccine 2013, 31, 6144–6149. [Google Scholar] [CrossRef]
- Xu, Q.Q.; Ma, X.J.; Wang, F.K.; Li, H.M.; Zhao, X.M. Evaluation of a multi-epitope subunit vaccine against avian leukosis virus subgroup J in chickens. Virus Res. 2015, 210, 62–68. [Google Scholar] [CrossRef]
- Xu, Q.Q.; Cui, N.; Ma, X.J.; Wang, F.K.; Li, H.M.; Shen, Z.Q.; Zhao, X.M. Evaluation of a chimeric multi-epitope-based DNA vaccine against subgroup J avian leukosis virus in chickens. Vaccine 2016, 34, 3751–3756. [Google Scholar] [CrossRef]
- Dai, M.M.; Feng, M.; Ye, Y.; Wu, X.C.; Liu, D.; Liao, M.; Cao, W.S. Exogenous avian leukosis virus-induced activation of the ERK/AP1 pathway is required for virus replication and correlates with virus-induced tumorigenesis. Sci. Rep. 2016, 6, 19226. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Liu, X.M.; Liu, H.X.; Xu, C.G.; Liao, Y.L.; Wu, X.C.; Cao, W.S.; Liao, M. Isolation, identification, and phylogenetic analysis of two avian leukosis virus subgroup J strains associated with hemangioma and myeloid leukosis. Vet. Microbiol. 2013, 166, 356–364. [Google Scholar] [CrossRef]
- Zhang, H.N.; Lai, H.Z.; Qi, Y.; Zhang, X.T.; Ning, Z.Y.; Luo, K.J.; Xin, C.A.; Cao, W.S.; Liao, M. An ALV-J isolate is responsible for spontaneous haemangiomas in layer chickens in China. Avian Pathol. 2011, 40, 261–267. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.W.; Chen, H.B.; Liu, J.Z.; Cheng, Z.Q. The critical time of avian leukosis virus subgroup J-mediated immunosuppression during early stage infection in specific pathogen-free chickens. J. Vet. Sci. 2011, 12, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Qualtiere, L.F.; Meyers, P. A reexamination of humoral tolerance in chickens congenitally infected with an avian leukosis virus. J. Immunol. 1979, 122, 825–829. [Google Scholar] [PubMed]
- Feng, M.; Dai, M.M.; Xie, T.T.; Li, Z.H.; Shi, M.Q.; Zhang, X.Q. Innate Immune Responses in ALV-J Infected Chicks and Chickens with Hemangioma In Vivo. Front. Microbiol. 2016, 7, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Zhang, X.Q. Immunity to Avian Leukosis Virus: Where Are We Now and What Should We Do? Front. Immunol. 2016, 7, 624. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.M.; Xu, C.G.; Chen, W.S.; Liao, M. Progress on chicken T cell immunity to viruses. Cell. Mol. Life Sci. 2019, 76, 2779–2788. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.M.; Wu, S.Y.; Feng, M.; Feng, S.X.; Sun, C.; Bai, D.Y.; Gu, M.Z.; Liao, M.; Cao, W.S. Recombinant chicken interferon-alpha inhibits the replication of exogenous avian leukosis virus (ALV) in DF-1 cells. Mol. Immunol. 2016, 76, 62–69. [Google Scholar] [CrossRef]
- Liu, D.; Qiu, Q.Q.; Zhang, X.; Dai, M.M.; Qin, J.R.; Hao, J.J.; Liao, M.; Cao, W.S. Infection of chicken bone marrow mononuclear cells with subgroup J avian leukosis virus inhibits dendritic cell differentiation and alters cytokine expression. Infect. Genet. Evol. 2016, 44, 130–136. [Google Scholar] [CrossRef]
- Liu, H.W.; Zhang, M.F.; Han, H.T.; Yuan, J.H.; Li, Z.D. Comparison of the expression of cytokine genes in the bursal tissues of the chickens following challenge with infectious bursal disease viruses of varying virulence. Virol. J. 2010, 7, 364. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.C.; Kim, B.S.; Wu, C.C.; Lin, T.L. Bursal transcriptome of chickens protected by DNA vaccination versus those challenged with infectious bursal disease virus. Arch. Virol. 2015, 160, 69–80. [Google Scholar] [CrossRef]
- Humphries, F.; Yang, S.; Wang, B.; Moynagh, P.N. RIP kinases: Key decision makers in cell death and innate immunity. Cell Death Differ. 2015, 22, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.F.; Liu, C.H.; Li, R.Q.; Zhang, H.Z.; Xu, X.G.; Wang, J.Y. Modulation of the innate immune-related genes expression in H9N2 avian influenza virus-infected chicken macrophage-like cells (HD11) in response to Escherichia coli LPS stimulation. Res. Vet. Sci. 2017, 111, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.L.; Li, Y.F.; Qi, W.; Ma, X.L.; Yu, K.X.; Huang, B.; Liao, M.; Li, F.; Pan, J.; Song, M.X. Comparative analysis of selected innate immune-related genes following infection of immortal DF-1 cells with highly pathogenic (H5N1) and low pathogenic (H9N2) avian influenza viruses. Virus Genes 2015, 50, 189–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarson, A.J.; Abdul-Careem, M.F.; Read, L.R.; Brisbin, J.T.; Sharif, S. Expression of Cytotoxicity-Associated Genes in Marek’s Disease Virus—Infected Chickens. Viral Immunol. 2008, 21, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Dunon, D.; Allioli, N.; Vainio, O.; Ody, C.; Imhof, B.A. Quantification of T-cell progenitors during ontogeny: Thymus colonization depends on blood delivery of progenitors. Blood 1999, 93, 2234–2243. [Google Scholar] [CrossRef] [PubMed]
- Luhtala, M. Chicken CD4, CD8alphabeta, and CD8alphaalpha T cell co-receptor molecules. Poult. Sci. 1998, 77, 1858–1873. [Google Scholar] [CrossRef]
- Dai, M.M.; Feng, M.; Xie, T.T.; Zhang, X.Q. Long non-coding RNA and MicroRNA profiling provides comprehensive insight into non-coding RNA involved host immune responses in ALV-J-infected chicken primary macrophage. Dev. Comp. Immunol. 2019, 100, 103414. [Google Scholar] [CrossRef]
- Dai, M.M.; Feng, M.; Xie, T.T.; Li, Y.F.; Ruan, Z.H.; Shi, M.Q.; Liao, M.; Zhang, X.Q. ALV-J infection induces chicken monocyte death accompanied with the production of IL-1beta and IL-18. Oncotarget 2017, 8, 99889–99900. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.S.; Wang, X.Y.; Zhai, J.; Qi, W.; Jing, L.; Ge, Y.Y.; Gao, X.L.; Liu, C.N.; Lv, X.P.; Zheng, S.M. Changes in apoptosis, proliferation and T lymphocyte subtype on thymic cells of SPF chickens infected with reticuloendotheliosis virus. Mol. Immunol. 2019, 111, 87–94. [Google Scholar] [CrossRef]
- Xue, M.; Shi, X.M.; Zhao, Y.; Cui, H.Y.; Hu, S.L.; Cui, X.L.; Wang, Y.F. Effects of Reticuloendotheliosis Virus Infection on Cytokine Production in SPF Chickens. PLoS ONE 2013, 8, e83918. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Q.; Rosa, A.J.M.; Oliverira, H.N.; Rosa, G.J.M.; Guo, X.S.; Travnicek, M.; Girshick, T. Transcriptome of Local Innate and Adaptive Immunity during Early Phase of Infectious Bronchitis Viral Infection. Viral Immunol. 2006, 19, 768–774. [Google Scholar] [CrossRef]
- Rauf, A.; Khatri, M.; Murgia, M.V.; Saif, Y.M. Expression of perforin–granzyme pathway genes in the bursa of infectious bursal disease virus-infected chickens. Dev. Comp. Immunol. 2011, 35, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Khatri, M.; Murgia, M.V.; Saif, Y.M. Fas/FasL and perforin-granzyme pathways mediated T cell cytotoxic responses in infectious bursal disease virus infected chickens. Results Immunol. 2012, 2, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, S.; Rothwell, L.; Degen, W.D.; Schijns, V.E.; Young, J.; Kaufman, J.; Kaiser, P. Characterization of the first nonmammalian T2 cytokine gene cluster: The cluster contains functional single-copy genes for IL-3, IL-4, IL-13, and GM-CSF, a gene for IL-5 that appears to be a pseudogene, and a gene encoding another cytokinelike transcript, KK34. J. Interferon Cytokine Res. 2004, 24, 600–610. [Google Scholar] [PubMed]
- Schoggins, J.W. Interferon-stimulated genes: Roles in viral pathogenesis. Curr. Opin. Virol. 2014, 6, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Yoneda, A.; Ninomiya, A.; Kawahara, M.; Watanabe, T. Both antiviral activity and intracellular localization of chicken Mx protein depend on a polymorphism at amino acid position 631. Biochem. Biophys. Res. Commun. 2013, 430, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santhakumar, D.; Rohaim, M.A.M.S.; Hussein, H.A.; Hawes, P.; Ferreira, H.L.; Behboudi, S.; Iqbal, M.; Nair, V.; Arns, C.W.; Munir, M. Chicken Interferon-induced Protein with Tetratricopeptide Repeats 5 Antagonizes Replication of RNA Viruses. Sci. Rep. 2018, 8, 6794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shini, S.; Shini, A.; Huff, G. Effects of chronic and repeated corticosterone administration in rearing chickens on physiology, the onset of lay and egg production of hens. Physiol. Behav. 2009, 98, 73–77. [Google Scholar] [CrossRef]
- Chappell, L.; Kaiser, P.; Barrow, P.; Jones, M.A.; Johnston, C.; Wigley, P. The immunobiology of avian systemic salmonellosis. Vet. Immunol. Immunopathol. 2009, 128, 53–59. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Sequence (5′–3′) | Gen Bank Accession No. | Reference |
|---|---|---|---|---|
| GAPDH | Forward | GAACATCATCCCAGCGTCCA | NM_204305.1 | [16] |
| Reverse | CGGCAGGTCAGGTCAACAAC | |||
| IFN-α | Forward | GACAGCCAACGCCAAAGC | GU119896.1 | [16] |
| Reverse | GTCGCTGCTGTCCAAGCATT | |||
| IFN-β | Forward | GCCCACACACTCCAAAACACTG | NM_001024836.1 | [16] |
| Reverse | TTGATGCTGAGGTGAGCGTTG | |||
| IL-1β | Forward | GGTCAACATCGCCACCTACA | NM_204524.1 | [16] |
| Reverse | CATACGAGATGGAAACCAGCAA | |||
| TNF-α | Forward | GCTGTTCTATGACCGCCCAGTT | NM_204267.1 | [16] |
| Reverse | AACAACCAGCTATGCACCCCA | |||
| IL-2 | Forward | GCTAATGACTACAGCTTATGGAGCA | AF000631.1 | [16] |
| Reverse | TGGGTCTCAGTTGGTGTGTAGAG | |||
| IL-6 | Forward | AAATCCCTCCTCGCCAATCT | AJ309540.1 | [16] |
| Reverse | CCCTCACGGTCTTCTCCATAAA | |||
| IL-10 | Forward | AGCAGATCAAGGAGACGTTC | NM_001004414.2 | [16] |
| Reverse | ATCAGCAGGTACTCCTCGAT | |||
| TLR3 | Forward | ACAATGGCAGATTGTAGTCACCT | NM_001011691.3 | [16] |
| Reverse | GCACAATCCTGGTTTCAGTTTAG | |||
| TLR7 | Forward | TCTGGACTTCTCTAACAACA | NM_001011688.2 | [16] |
| Reverse | AATCTCATTCTCATTCATCATCA | |||
| MHC-I | Forward | AAGAAGGGGAAGGGCTACAA | NM_001031338.1 | [16] |
| Reverse | AAGCAGTGCAGGCAAAGAAT | |||
| MHC-II | Forward | CTCGAGGTCATGATCAGCAA | DQ008588.2 | [16] |
| Reverse | TGTAAACGTCTCCCCTTTGG | |||
| IFN-γ | Forward | CCTCCAACACCTCTTCAACATG | X92479 | [17] |
| Reverse | TGGCGTGCGGTCAAT | |||
| IL-4 | Forward | TCGAGGAGTGACGGGTG | AJ621249.1 | [17] |
| Reverse | ACTATCCGGATGCTCTCCATC | |||
| IL-5 | Forward | GGAACGGCACTGTTGAAAAATAA | AJ621252.1 | [18] |
| Reverse | TTCTCCCTCTCCTGTCAGTTGTG | |||
| IL-13 | Forward | CTGCCCTTGCTCTCCTCTGT | AJ621250.1 | [18] |
| Reverse | CCTGCACTCCTCTGTTGAGCTT | |||
| Granzyme A | Forward | ACTCATGTCGAGGGGATTCA | NM_204457.1 | [19] |
| Reverse | TGTAGACACCAGGACCACCA | |||
| Granzyme K | Forward | CGGGAAGCAACTGTTGAAAT | XM_423832 | [19] |
| Reverse | GAGTCTCCCTTGCAAGCATC | |||
| MDA5 | Forward | GGACGACCACGATCTCTGTGT | NM_001193638.1 | [20] |
| Reverse | CACCTGTCTGGTCTGCATGTTATC | |||
| CXCLi1 | Forward | AACTCCGATGCCAGTG | NM_205018.1 | [21] |
| Reverse | TTGGTGTCTGCCTTGT | |||
| CXCLi2 | Forward | CATCATGAAGCATTCCATCT | NM_205498.1 | [21] |
| Reverse | CTTCCAAGGGATCTTCATTT | |||
| TGF-β3 | Forward | TCTTTACATTGACTTCCGAC | NM_205454.1 | [21] |
| Reverse | TCCTCCCAACATAGTACAAG | |||
| MX1 | Forward | AAGCCTGAGCATGAGCAGAA | NM_204609.1 | [22] |
| Reverse | TCTCAGGCTGTCAACAAGATCAA | |||
| OASL | Forward | AGATGTTGAAGCCGAAGTACCC | NM_205041.1 | [22] |
| Reverse | CTGAAGTCCTCCCTGCCTGT | |||
| ISG12-2 | Forward | TCAATGGGTGGCAAAGGAG | NM_001001296.5 | [22] |
| Reverse | TACAGGGAGAGCAAAGAAGAGAAGA | |||
| IFIT5 | Forward | CAGAATTTAATGCCGGCTATGC | XM_421662.4 | [22] |
| Reverse | TGCAAGTAAAGCCAAAAGATAAGTGT | |||
| USP18 | Forward | CAACGTGGGAAGAGGAGAAA | XM_416398.3 | [22] |
| Reverse | ACTTCATGAGCGGAGAAGGA | |||
| IRF3/7 | Forward | ACTGACCAGCCCAGGAACTCT | NM_205372.1 | [22] |
| Reverse | AAGGCTTTCCCAACCACAAA | |||
| SST | Forward | GGTCCACGGTTATGGTGAAAG | NM_205336.1 | [22] |
| Reverse | GGTCAGAAATCACAACTCAAGCA | |||
| KHSRP | Forward | CAGCGGGGAAATGATTAAGAAG | NM_204277.1 | [22] |
| Reverse | TTTGTGTGTGGGGATGGAGA | |||
| Perforin | Forward | ATGGCGCAGGTGACAGTGA | XM_425355 | [23] |
| Reverse | TGGCCTGCACCGGTAATTC | |||
| PARP | Forward | ATTGTGGAGGAGCTGGGAGGAA | NM_205263 | [23] |
| Reverse | AGGCTTGCTGCACTTCCCATC | |||
| HMG-2 | Forward | AGAGCACAAGAAGAAGCAC | M80574 | [23] |
| Reverse | GTCTTTTAGGAGCGTTGGGGTC | |||
| NK lysin | Forward | GATGGTTCAGCTGCGTGGGATGC | DQ186291 | [23] |
| Reverse | CTGCCGGAGCTTCTTCAACA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, M.; Li, S.; Shi, K.; Liao, J.; Sun, H.; Liao, M. Systematic Identification of Host Immune Key Factors Influencing Viral Infection in PBL of ALV-J Infected SPF Chicken. Viruses 2020, 12, 114. https://doi.org/10.3390/v12010114
Dai M, Li S, Shi K, Liao J, Sun H, Liao M. Systematic Identification of Host Immune Key Factors Influencing Viral Infection in PBL of ALV-J Infected SPF Chicken. Viruses. 2020; 12(1):114. https://doi.org/10.3390/v12010114
Chicago/Turabian StyleDai, Manman, Shibing Li, Keyi Shi, Jiayu Liao, Hui Sun, and Ming Liao. 2020. "Systematic Identification of Host Immune Key Factors Influencing Viral Infection in PBL of ALV-J Infected SPF Chicken" Viruses 12, no. 1: 114. https://doi.org/10.3390/v12010114