Support for the Transmission-Clearance Trade-Off Hypothesis from a Study of Zika Virus Delivered by Mosquito Bite to Mice

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Study Design and Ethics Statement
2.2. Mosquitoes
2.3. Cell Lines and Viruses
2.4. Titrations
2.5. Focus Forming Assay
2.6. Plaque Reduction Neutralization Test Assay
3. Results
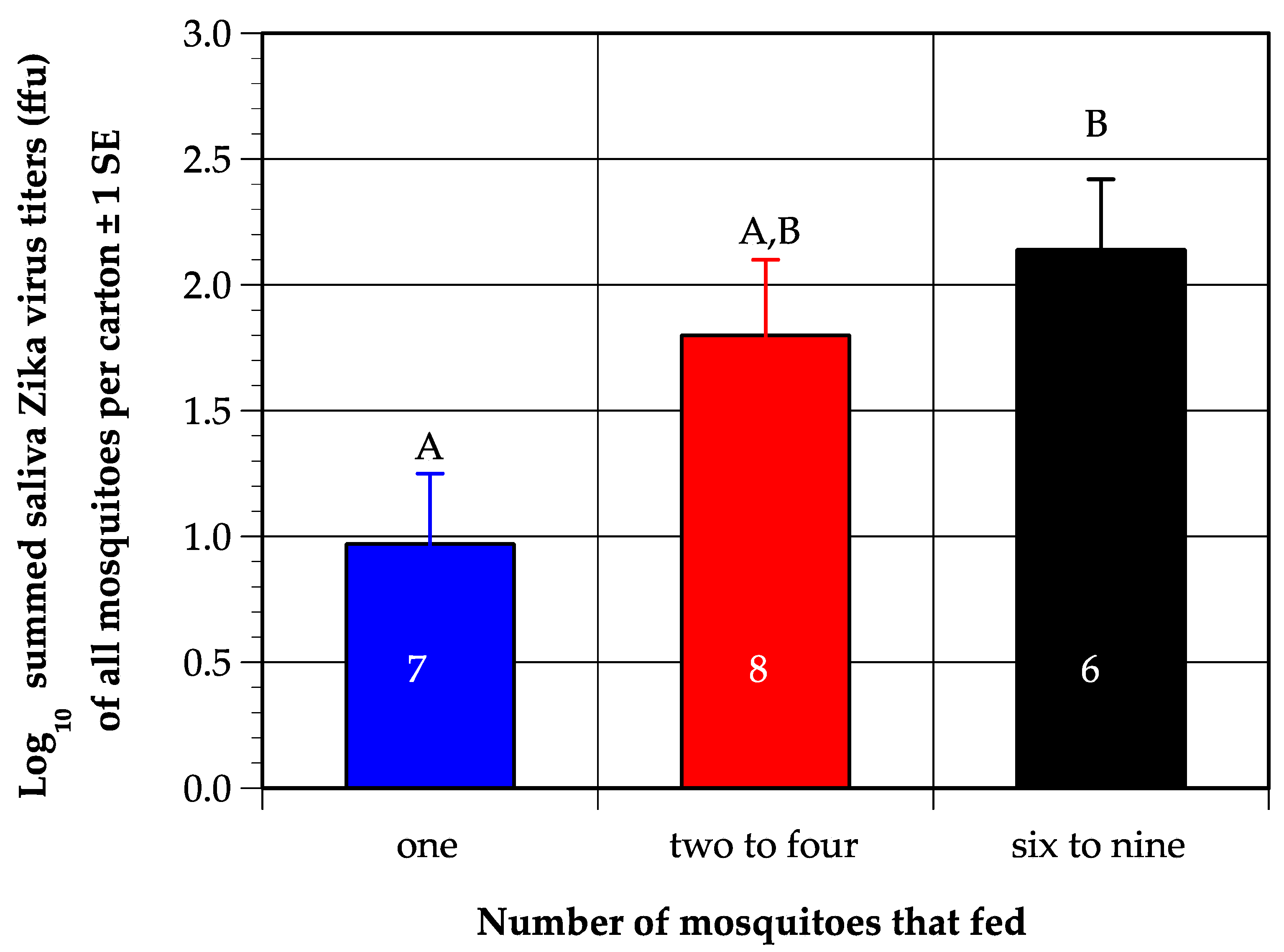
3.1. Mosquito Feeding Success, Detection of Virus in Saliva, and Grouping of Mice for Analysis
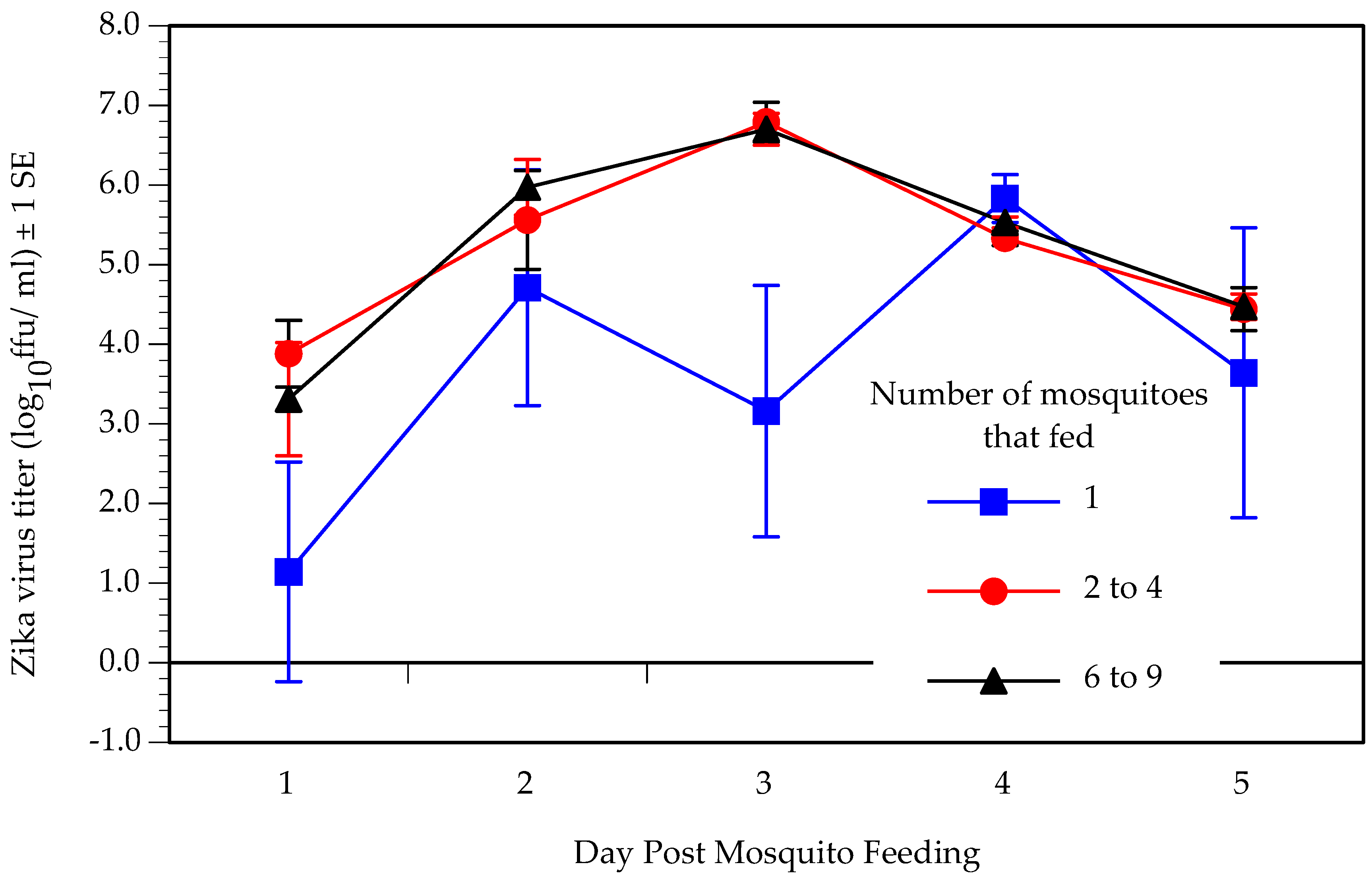
3.2. Intra-Host Virus Dynamics
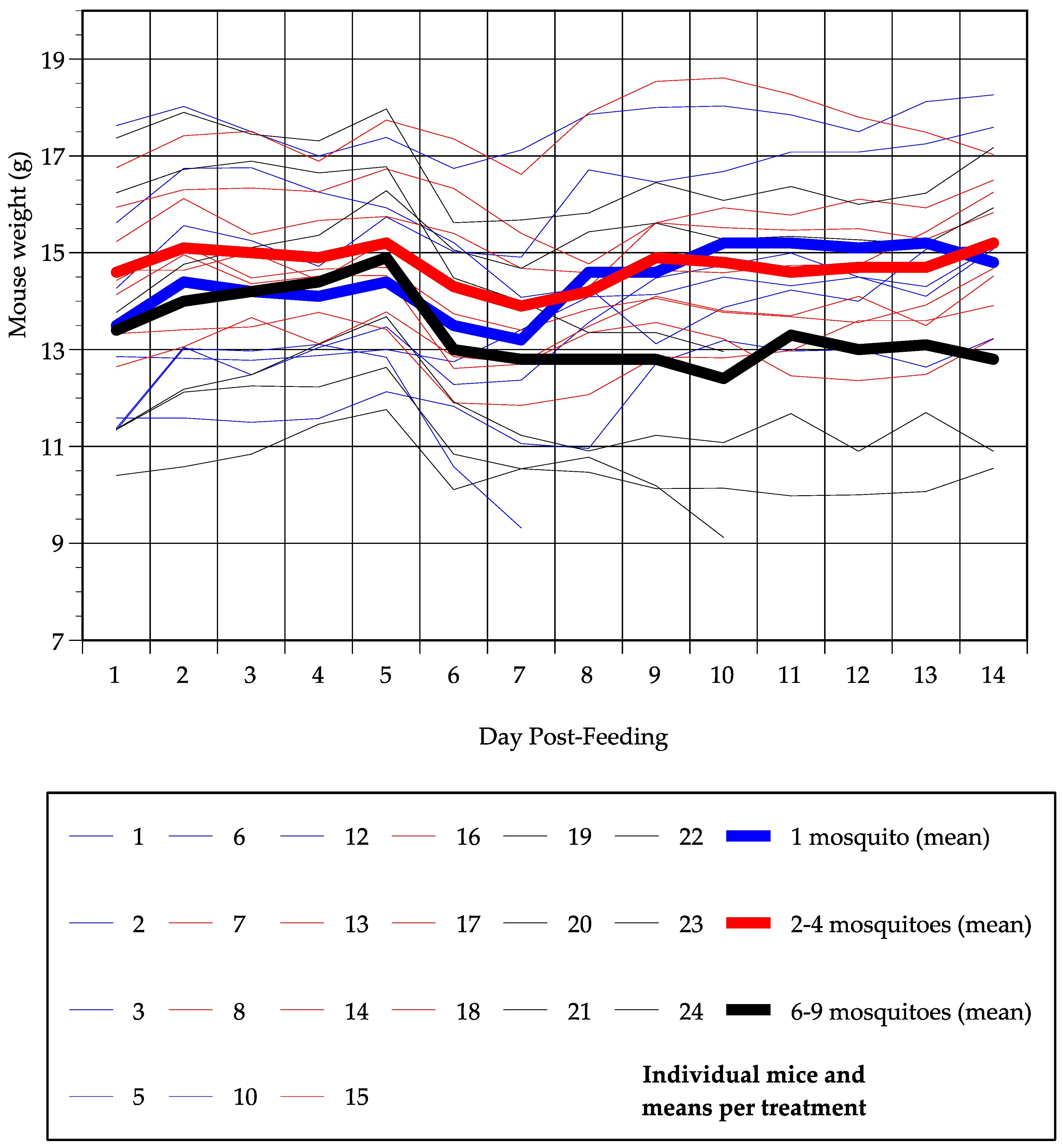
3.3. Mouse Survival and Weight Changes
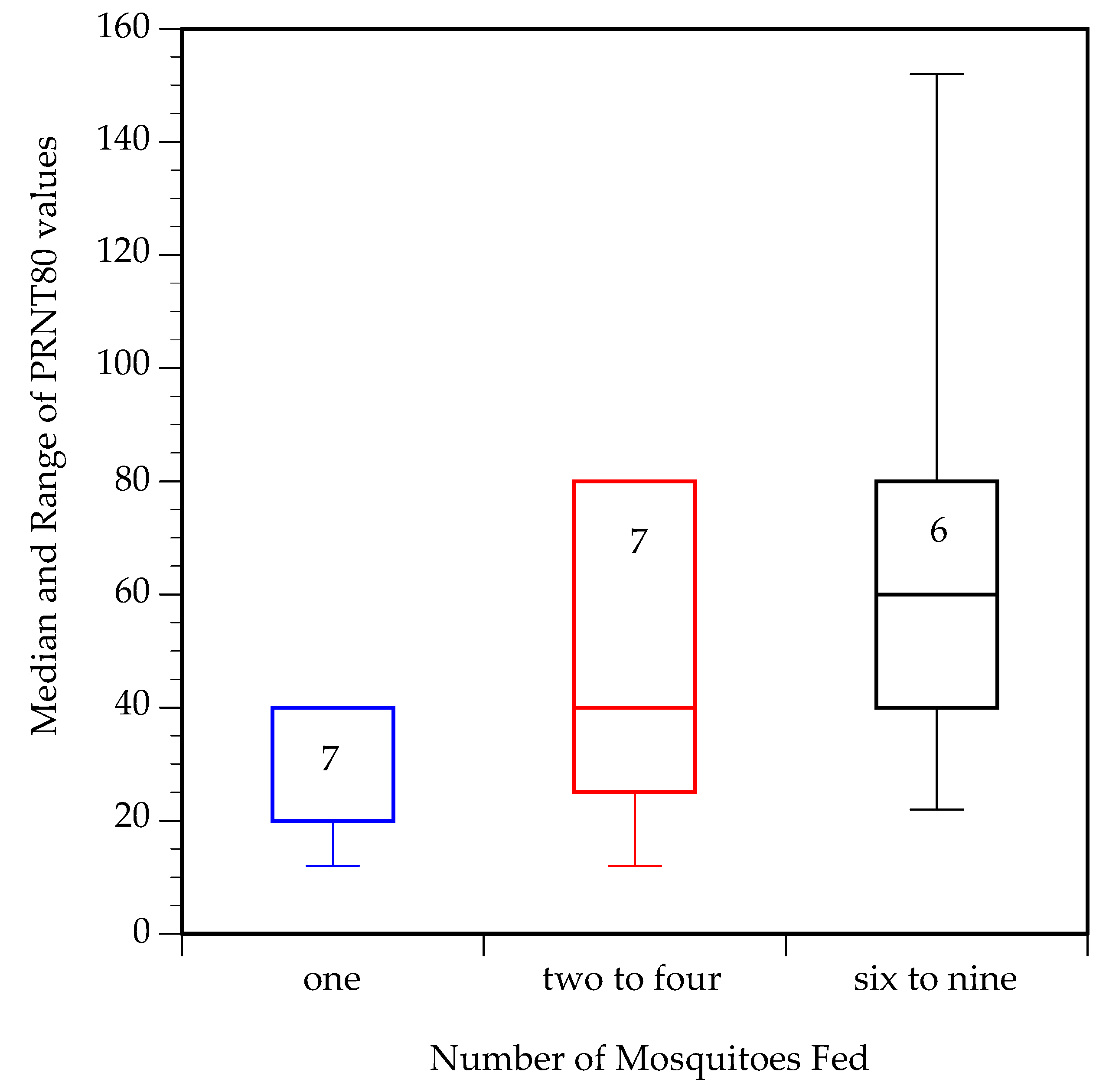
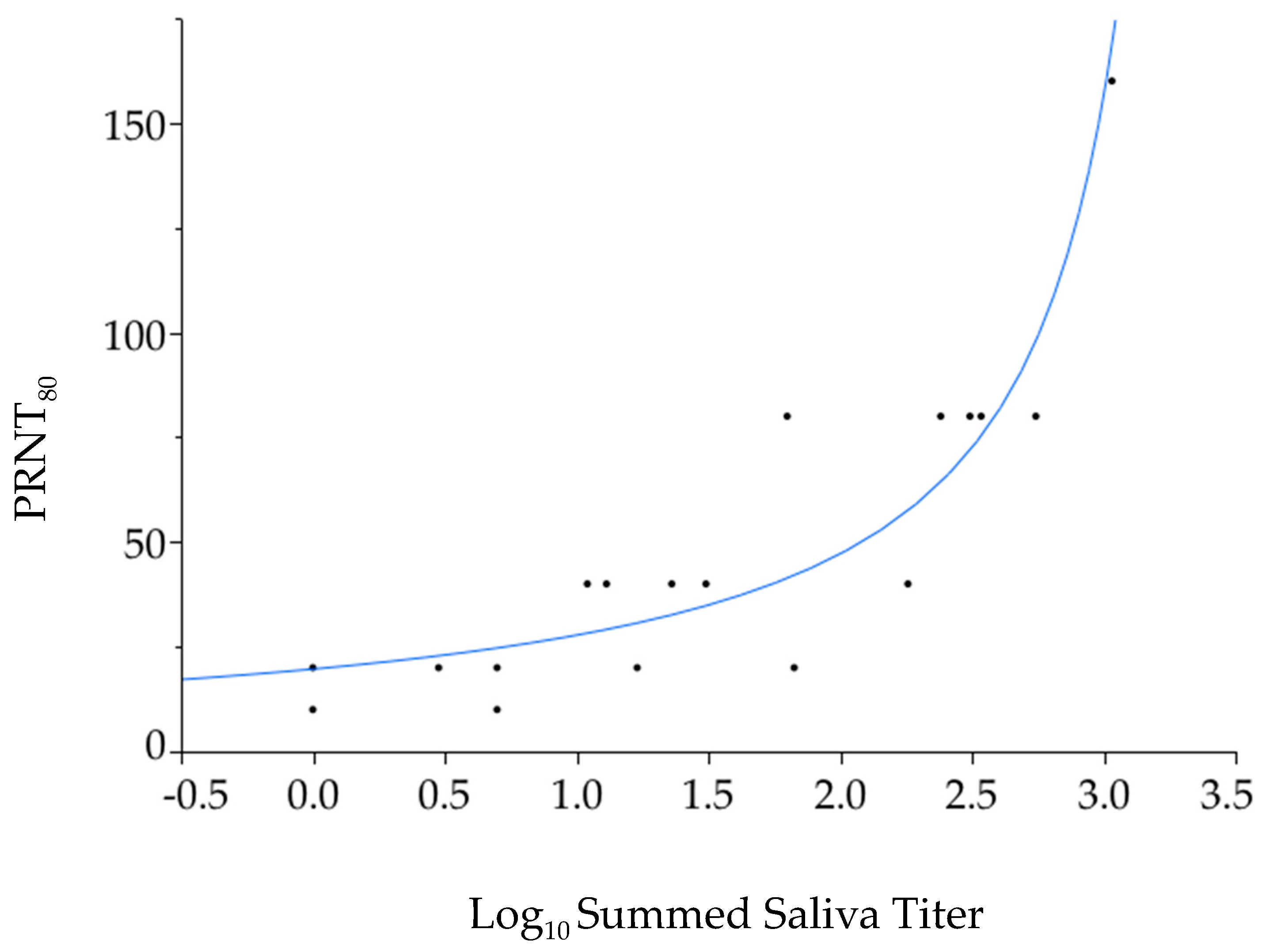
3.4. Seroconversion
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chao, L.; Hanley, K.A.; Burch, C.L.; Dahlberg, C.; Turner, P.E. Kin selection and parasite evolution: Higher and lower virulence with hard and soft selection. Q. Rev. Biol. 2000, 75, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.A. Models of parasite virulence. Q. Rev. Biol. 1996, 71, 37–78. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.A.; Schmid-Hempel, P. Mechanisms of pathogenesis and the evolution of parasite virulence. J. Evol. Biol. 2008, 21, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, M.J.; Read, A.F. Virulence in malaria: An evolutionary viewpoint. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 965–986. [Google Scholar] [CrossRef]
- Ewald, P.W. Evolution of virulence. Infect. Dis. Clin. N. Am. 2004, 18, 1–15. [Google Scholar] [CrossRef]
- Ebert, D.; Bull, J.J. Challenging the trade-off model for the evolution of virulence: Is virulence management feasible? Trends Microbiol. 2003, 11, 15–20. [Google Scholar] [CrossRef]
- Bull, J.J.; Lauring, A.S. Theory and empiricism in virulence evolution. PLoS Pathog. 2014, 10, e1004387. [Google Scholar] [CrossRef]
- Feng, Z.; Cen, X.; Zhao, Y.; Velasco-Hernandez, J.X. Coupled within-host and between-host dynamics and evolution of virulence. Math. Biosci. 2015, 270, 204–212. [Google Scholar] [CrossRef]
- Humplik, J.; Hill, A.L.; Nowak, M.A. Evolutionary dynamics of infectious diseases in finite populations. J. Theor. Biol. 2014, 360, 149–162. [Google Scholar] [CrossRef]
- Kamo, M.; Sasaki, A.; Boots, M. The role of trade-off shapes in the evolution of parasites in spatial host populations: An approximate analytical approach. J. Theor. Biol. 2007, 244, 588–596. [Google Scholar] [CrossRef]
- Alizon, S.; Hurford, A.; Mideo, N.; Van Baalen, M. Virulence evolution and the trade-off hypothesis: History, current state of affairs and the future. J. Evol. Biol. 2009, 22, 245–259. [Google Scholar] [CrossRef]
- Alizon, S. Transmission-recovery trade-offs to study parasite evolution. Am. Nat. 2008, 172, E113–E121. [Google Scholar] [CrossRef] [PubMed]
- Long, G.H.; Graham, A.L. Consequences of immunopathology for pathogen virulence evolution and public health: Malaria as a case study. Evol. Appl. 2011, 4, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.P.; McPherson, K.; Van Vinh Chau, N.; Hoai Tam, D.T.; Young, P.; Mackenzie, J.; Wills, B. Recent advances in dengue pathogenesis and clinical management. Vaccine 2015, 33, 7061–7068. [Google Scholar] [CrossRef] [PubMed]
- Simonin, Y.; Loustalot, F.; Desmetz, C.; Foulongne, V.; Constant, O.; Fournier-Wirth, C.; Leon, F.; Moles, J.P.; Goubaud, A.; Lemaitre, J.M.; et al. Zika Virus Strains Potentially Display Different Infectious Profiles in Human Neural Cells. EBioMedicine 2016, 12, 161–169. [Google Scholar] [CrossRef]
- Simonin, Y.; van Riel, D.; Van de Perre, P.; Rockx, B.; Salinas, S. Differential virulence between Asian and African lineages of Zika virus. PLoS Negl. Trop. Dis. 2017, 11, e0005821. [Google Scholar] [CrossRef]
- Martina, B.E.; Koraka, P.; Osterhaus, A.D. Dengue virus pathogenesis: An integrated view. Clin. Microbiol. Rev. 2009, 22, 564–581. [Google Scholar] [CrossRef]
- Nunes, P.C.G.; Nogueira, R.M.R.; Heringer, M.; Chouin-Carneiro, T.; Damasceno Dos Santos Rodrigues, C.; de Filippis, A.M.B.; Lima, M.; Dos Santos, F.B. NS1 Antigenemia and Viraemia Load: Potential Markers of Progression to Dengue Fatal Outcome? Viruses 2018, 10, 326. [Google Scholar] [CrossRef]
- Ng, D.H.L.; Ho, H.J.; Chow, A.; Wong, J.; Kyaw, W.M.; Tan, A.; Chia, P.Y.; Choy, C.Y.; Tan, G.; Yeo, T.W.; et al. Correlation of clinical illness with viremia in Zika virus disease during an outbreak in Singapore. BMC Infect. Dis. 2018, 18, 301. [Google Scholar] [CrossRef]
- Althouse, B.M.; Hanley, K.A. The tortoise or the hare? Impacts of within-host dynamics on transmission success of arthropod-borne viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140299. [Google Scholar] [CrossRef]
- Azar, S.R.; Roundy, C.M.; Rossi, S.L.; Huang, J.H.; Leal, G.; Yun, R.; Fernandez-Salas, I.; Vitek, C.J.; Paploski, I.A.D.; Stark, P.M.; et al. Differential vector competency of Aedes albopictus populations from the Americas for Zika virus. Am. J. Trop. Med. Hyg. 2017, 97, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, J.; Kien, D.T.; Nguyen, N.M.; Nguyen, H.L.; Kyrylos, P.P.; Carrington, L.B.; Tran, C.N.; Quyen, N.T.; Thi, L.V.; Le Thi, D.; et al. Comparative Susceptibility of Aedes albopictus and Aedes aegypti to Dengue Virus Infection After Feeding on Blood of Viremic Humans: Implications for Public Health. J. Infect. Dis. 2015, 212, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Roundy, C.M.; Azar, S.R.; Rossi, S.L.; Huang, J.H.; Leal, G.; Yun, R.; Fernandez-Salas, I.; Vitek, C.J.; Paploski, I.A.; Kitron, U.; et al. Variation in Aedes aegypti Mosquito Competence for Zika Virus Transmission. Emerg. Infect. Dis. 2017, 23, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Tesla, B.; Demakovsky, L.R.; Packiam, H.S.; Mordecai, E.A.; Rodriguez, A.D.; Bonds, M.H.; Brindley, M.A.; Murdock, C.C. Estimating the effects of variation in viremia on mosquito susceptibility, infectiousness, and R0 of Zika in Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006733. [Google Scholar] [CrossRef] [PubMed]
- Dudley, D.M.; Newman, C.M.; Lalli, J.; Stewart, L.M.; Koenig, M.R.; Weiler, A.M.; Semler, M.R.; Barry, G.L.; Zarbock, K.R.; Mohns, M.S.; et al. Infection via mosquito bite alters Zika virus tissue tropism and replication kinetics in rhesus macaques. Nat. Commun. 2017, 8, 2096. [Google Scholar] [CrossRef]
- Le Coupanec, A.; Babin, D.; Fiette, L.; Jouvion, G.; Ave, P.; Misse, D.; Bouloy, M.; Choumet, V. Aedes mosquito saliva modulates Rift Valley fever virus pathogenicity. PLoS Negl. Trop. Dis. 2013, 7, e2237. [Google Scholar] [CrossRef]
- Thangamani, S.; Higgs, S.; Ziegler, S.; Vanlandingham, D.; Tesh, R.; Wikel, S. Host immune response to mosquito-transmitted chikungunya virus differs from that elicited by needle inoculated virus. PLoS ONE 2010, 5, e12137. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Coffey, L.L.; Stevenson, H.L.; McGee, C.E.; Higgs, S. Aedes aegypti saliva alters leukocyte recruitment and cytokine signaling by antigen-presenting cells during West Nile virus infection. PLoS ONE 2010, 5, e11704. [Google Scholar] [CrossRef]
- Jin, L.; Guo, X.; Shen, C.; Hao, X.; Sun, P.; Li, P.; Xu, T.; Hu, C.; Rose, O.; Zhou, H.; et al. Salivary factor LTRIN from Aedes aegypti facilitates the transmission of Zika virus by interfering with the lymphotoxin-beta receptor. Nat. Immunol. 2018, 19, 342–353. [Google Scholar] [CrossRef]
- Ben-Shachar, R.; Koelle, K. Transmission-clearance trade-offs indicate that dengue virulence evolution depends on epidemiological context. Nat. Commun. 2018, 9, 2355. [Google Scholar] [CrossRef]
- Serman, T.M.; Gack, M.U. Evasion of Innate and Intrinsic Antiviral Pathways by the Zika Virus. Viruses 2019, 11, 970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, C.; Muruato, A.E.; Nunes, B.T.D.; Luo, H.; Xie, X.; Medeiros, D.B.A.; Wakamiya, M.; Tesh, R.B.; Barrett, A.D.; Wang, T.; et al. A live-attenuated Zika virus vaccine candidate induces sterilizing immunity in mouse models. Nat. Med. 2017, 23, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Durbin, A.P.; Mayer, S.V.; Rossi, S.L.; Amaya-Larios, I.Y.; Ramos-Castaneda, J.; Eong Ooi, E.; Jane Cardosa, M.; Munoz-Jordan, J.L.; Tesh, R.B.; Messer, W.B.; et al. Emergence potential of sylvatic dengue virus type 4 in the urban transmission cycle is restrained by vaccination and homotypic immunity. Virology 2013, 439, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azar, S.R.; Rossi, S.L.; Haller, S.H.; Yun, R.; Huang, J.H.; Plante, J.A.; Zhou, J.; Olano, J.P.; Roundy, C.M.; Hanley, K.A.; et al. ZIKV demonstrates minimal pathologic effects and mosquito infectivity in viremic cynomolgus macques. Viruses 2018, 10, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.L.; Tesh, R.B.; Azar, S.R.; Muruato, A.E.; Hanley, K.A.; Auguste, A.J.; Langsjoen, R.M.; Paessler, S.; Vasilakis, N.; Weaver, S.C. Characterization of a Novel Murine Model to Study Zika Virus. Am. J. Trop. Med. Hyg. 2016, 94, 1362–1369. [Google Scholar] [CrossRef] [Green Version]
- Styer, L.M.; Kent, K.A.; Albright, R.G.; Bennett, C.J.; Kramer, L.D.; Bernard, K.A. Mosquitoes inoculate high doses of West Nile virus as they probe and feed on live hosts. PLoS Pathog. 2007, 3, 1262–1270. [Google Scholar] [CrossRef] [Green Version]
- Pingen, M.; Schmid, M.A.; Harris, E.; McKimmie, C.S. Mosquito Biting Modulates Skin Response to Virus Infection. Trends Parasitol. 2017, 33, 645–657. [Google Scholar] [CrossRef]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host Inflammatory Response to Mosquito Bites Enhances the Severity of Arbovirus Infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef] [Green Version]
- Hastings, A.K.; Uraki, R.; Gaitsch, H.; Dhaliwal, K.; Stanley, S.; Sproch, H.; Williamson, E.; MacNeil, T.; Marin-Lopez, A.; Hwang, J.; et al. Aedes aegypti NeSt1 Protein Enhances Zika Virus Pathogenesis by Activating Neutrophils. J. Virol. 2019, 93, e00395-19. [Google Scholar] [CrossRef] [Green Version]
- Althouse, B.M.; Durbin, A.P.; Hanley, K.A.; Halstead, S.B.; Weaver, S.C.; Cummings, D.A. Viral kinetics of primary dengue virus infection in non-human primates: A systematic review and individual pooled analysis. Virology 2014, 452–453, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Gorman, M.J.; Caine, E.A.; Zaitsev, K.; Begley, M.C.; Weger-Lucarelli, J.; Uccellini, M.B.; Tripathi, S.; Morrison, J.; Yount, B.L.; Dinnon, K.H., 3rd; et al. An Immunocompetent Mouse Model of Zika Virus Infection. Cell Host Microbe 2018, 23, 672–685 e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.V.; Shaeer, K.M.; Patel, P.; Garmaza, A.; Wiangkham, K.; Franks, R.B.; Pane, O.; Carris, N.W. EPA-Registered Repellents for Mosquitoes Transmitting Emerging Viral Disease. Pharmacotherapy 2016, 36, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Che-Mendoza, A.; Medina-Barreiro, A.; Koyoc-Cardena, E.; Uc-Puc, V.; Contreras-Perera, Y.; Herrera-Bojorquez, J.; Dzul-Manzanilla, F.; Correa-Morales, F.; Ranson, H.; Lenhart, A.; et al. House screening with insecticide-treated netting provides sustained reductions in domestic populations of Aedes aegypti in Merida, Mexico. PLoS Negl. Trop. Dis. 2018, 12, e0006283. [Google Scholar] [CrossRef] [PubMed]
- Lenhart, A.; Orelus, N.; Maskill, R.; Alexander, N.; Streit, T.; McCall, P.J. Insecticide-treated bednets to control dengue vectors: Preliminary evidence from a controlled trial in Haiti. Trop. Med. Int. Health 2008, 13, 56–67. [Google Scholar] [CrossRef]
- LaRocque, R.L.; Ryan, E.T. Personal Actions to Minimize Mosquito-Borne Illnesses, Including Zika Virus. Ann. Intern. Med. 2016, 165, 589–590. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID # (Gen-der) | Original Treatment Group (no. Mosquitoes/Carton) | No. Mosquitoes that Fed | No. Mosquitoes with Detectable Virus in Saliva (Summed Virus Titer log10ffu) | Final Treatment Group (Used for Statistical Analysis) | Mouse Viremia (log10ffu/mL Serum) on Designated Day Post-Feeding. Greyed out Cells Indicate that Mouse Was Not Sampled on that Day. | PRNT80 at Day 14 Post-Feeding | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ||||||
| 1 (M) | 1 | 1 | 1 (1.1) | 1 | 3.32 | 4.52 | 5.60 | 40 | ||
| 2 (M) | 1 | 1 | 1 (1.23) | 1 | <LOD 1 | 4.95 | 5.32 | 20 | ||
| 3 (M) | 1 | 1 | 1 (0.70) | 1 | <LOD | <LOD | <LOD | <20 | ||
| 4 (F) | 1 | 0 | NA 2 | Excluded | <LOD | <LOD | <20 | |||
| 5 (F) | 1 | 1 | 0 | 1 | 3.59 | 6.38 | 20 | |||
| 6 (F) | 1 | 1 | 1 (0.48) | 1 | 3.38 | 5.38 | 20 | |||
| 7 (M) | 2 | 2 | 1 (2.79) | 2 to 4 | 3.36 | 7.15 | 4.51 | ND 3 | ||
| 8 (M) | 2 | 2 | 1 (0.70) | 2 to 4 | 3.65 | 7.28 | 4.30 | 20 | ||
| 9 (M) | 2 | 1 | 0 | Excluded | <LOD | <LOD | <LOD | <20 | ||
| 10 (M) | 2 | 1 | 1 (1.04) | 1 | 6.49 | 6.30 | 40 | |||
| 11 (M) | 2 | 1 | 0 | Excluded | <LOD | <LOD | <20 | |||
| 12 (M) | 2 | 1 | 1 (2.26) | 1 | 5.38 | 5.26 | 40 | |||
| 13 (M) | 5 | 3 | 1 (2.26) | 2 to 4 | 5.28 | 6.32 | 3.62 | 40 | ||
| 14 (M) | 5 | 4 | 3 (2.49) | 2 to 4 | 2.85 | 7.15 | 4.48 | 80 | ||
| 15 (M) | 5 | 3 | 3 (1.80) | 2 to 4 | 4.26 | 6.04 | 5.30 | 80 | ||
| 16 (F) | 5 | 4 | 2 (2.54) | 2 to 4 | 4.3 | 5.4 | 80 | |||
| 17 (F) | 5 | 4 | 1 (0.70) | 2 to 4 | 6.20 | 5.43 | <20 | |||
| 18 (F) | 5 | 3 | 3 (1.11) | 2 to 4 | 6.15 | 5.15 | 40 | |||
| 19 (M) | 10 | 7 | 4 (1.83) | 6 to 9 | 2.0 | 6.59 | 4.63 | 20 | ||
| 20 (M) | 10 | 7 | 4 (1.49) | 6 to 9 | 4.45 | 6.42 | 4.63 | 40 | ||
| 21 (M) | 10 | 6 | 6 (3.03) | 6 to 9 | 3.49 | 7.08 | 4.15 | 160 | ||
| 22 (F) | 10 | 8 | 6 (2.74) | 6 to 9 | 6.49 | 5.38 | 80 | |||
| 23 (F) | 10 | 9 | 3 (1.36) | 6 to 9 | 5.30 | 5.62 | 40 | |||
| 24 (F) | 10 | 8 | 4 (2.38) | 6 to 9 | 6.11 | 5.58 | 80 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanley, K.A.; Azar, S.R.; Campos, R.K.; Vasilakis, N.; Rossi, S.L. Support for the Transmission-Clearance Trade-Off Hypothesis from a Study of Zika Virus Delivered by Mosquito Bite to Mice. Viruses 2019, 11, 1072. https://doi.org/10.3390/v11111072
Hanley KA, Azar SR, Campos RK, Vasilakis N, Rossi SL. Support for the Transmission-Clearance Trade-Off Hypothesis from a Study of Zika Virus Delivered by Mosquito Bite to Mice. Viruses. 2019; 11(11):1072. https://doi.org/10.3390/v11111072
Chicago/Turabian StyleHanley, Kathryn A., Sasha R. Azar, Rafael K. Campos, Nikos Vasilakis, and Shannan L. Rossi. 2019. "Support for the Transmission-Clearance Trade-Off Hypothesis from a Study of Zika Virus Delivered by Mosquito Bite to Mice" Viruses 11, no. 11: 1072. https://doi.org/10.3390/v11111072