Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Variants Isolated in Eastern Canada Show Evidence of Recombination

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation
2.2. RNA Extraction and Complementary Deoxyribonucleic Acid (cDNA) Synthesis
2.3. Real-Time PCR and Sequencing
2.4. Genotyping and Comparison of S1 Gene Sequences
2.5. Characteristics of the Complete Genome
2.6. Recombination Analysis
2.7. GenBank Accession Numbers
3. Results
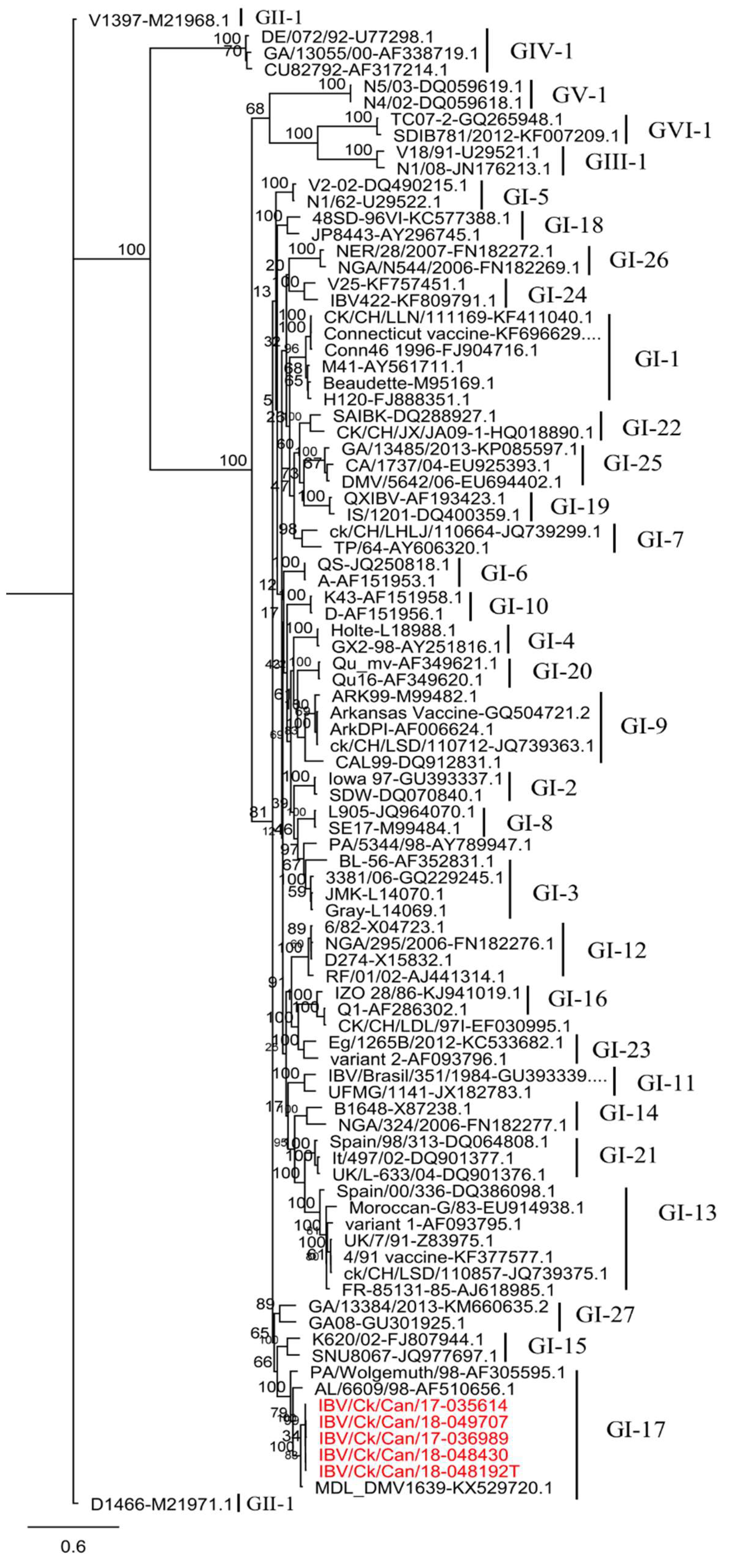
3.1. Phylogenetic Analysis and Sequence Comparison Based on the S1 Gene
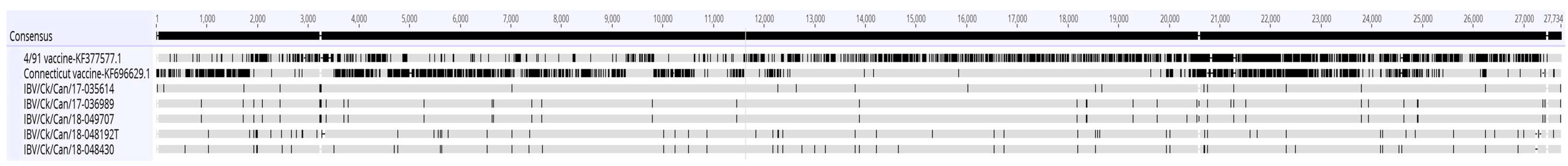
3.2. Complete Genome Sequences
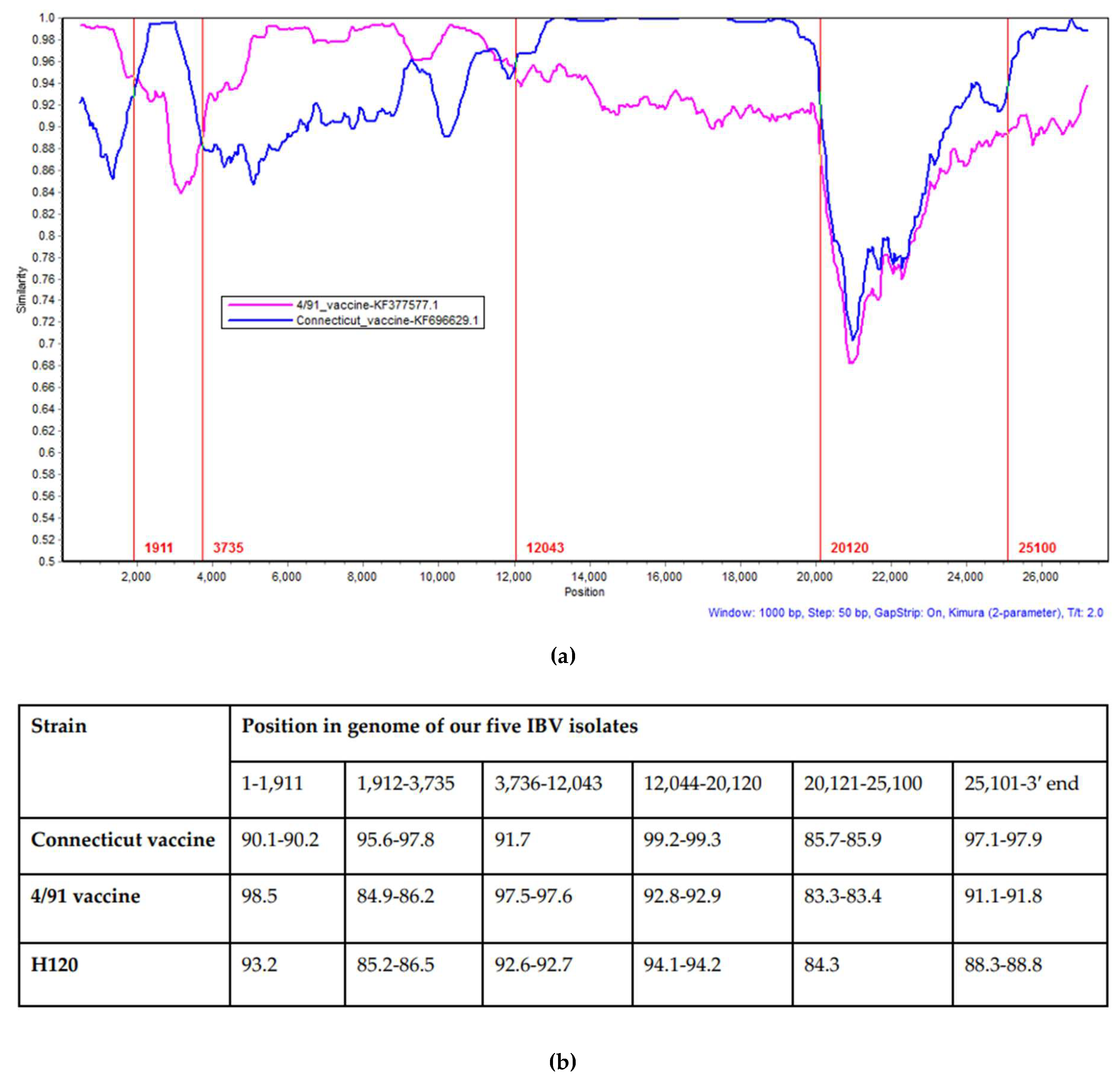
3.3. Recombination Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jackwood, M.W.; de Wit, S. Infectious bronchitis. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, R.J., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; John Wiley and Sons: Ames, IA, USA, 2013; Volume 13, pp. 139–159. [Google Scholar]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Armesto, M.; Cavanagh, D.; Britton, P. The replicase gene of avian coronavirus infectious bronchitis virus is a determinant of pathogenicity. PLoS ONE 2009, 4, e7384. [Google Scholar] [CrossRef] [PubMed]
- Quinteros, J.A.; Markham, P.F.; Lee, S.-W.; Hewson, K.A.; Hartley, C.A.; Legione, A.R.; Coppo, M.J.; Vaz, P.K.; Browning, G.F. Analysis of the complete genomic sequences of two virus subpopulations of the Australian infectious bronchitis virus vaccine VicS. Avian Pathol. 2015, 44, 182–191. [Google Scholar] [CrossRef]
- Wickramasinghe, I.N.; van Beurden, S.J.; Weerts, E.A.; Verheije, M.H. The avian coronavirus spike protein. Virus Res. 2014, 194, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef]
- Ammayappan, A.; Upadhyay, C.; Gelb, J., Jr.; Vakharia, V.N. Complete genomic sequence analysis of infectious bronchitis virus Ark DPI strain and its evolution by recombination. Virol. J. 2008, 5, 157. [Google Scholar] [CrossRef]
- Lim, T.H.; Lee, H.J.; Lee, D.H.; Lee, Y.N.; Park, J.K.; Youn, H.N.; Kim, M.S.; Lee, J.B.; Park, S.Y.; Choi, I.S.; et al. An emerging recombinant cluster of nephropathogenic strains of avian infectious bronchitis virus in Korea. Infect. Genet. Evol. 2011, 11, 678–685. [Google Scholar] [CrossRef]
- Moreno, A.; Franzo, G.; Massi, P.; Tosi, G.; Blanco, A.; Antilles, N.; Biarnes, M.; Majo, N.; Nofrarias, M.; Dolz, R.; et al. A novel variant of the infectious bronchitis virus resulting from recombination events in Italy and Spain. Avian Pathol. 2017, 46, 28–35. [Google Scholar] [CrossRef]
- Zhou, S.; Tang, M.; Jiang, Y.; Chen, X.; Shen, X.; Li, J.; Dai, Y.; Zou, J. Complete genome sequence of a novel infectious bronchitis virus strain circulating in China with a distinct S gene. Virus Genes 2014, 49, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Han, Z.; Xu, Q.; Wang, Q.; Gao, M.; Wu, W.; Shao, Y.; Li, H.; Kong, X.; Liu, S. Serotype shift of a 793/B genotype infectious bronchitis coronavirus by natural recombination. Infect. Genet. Evol. 2015, 32, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ma, H.; Xu, Q.; Sun, N.; Han, Z.; Sun, C.; Guo, H.; Shao, Y.; Kong, X.; Liu, S. Characterization of a recombinant coronavirus infectious bronchitis virus with distinct S1 subunits of spike and nucleocapsid genes and a 3′ untranslated region. Vet. Microbiol. 2013, 162, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Brash, M.; Stalker, M.; Ojkic, D. Using phylogenetic analysis to examine the changing strains of infectious bronchitis virus infections in Ontario over time. In Proceedings of the 16th Annual Meeting of the Canadian Animal Health Laboratorians Network, Guelph, ON, Canada, 4–7 June 2017. [Google Scholar]
- Petrik, M. Clinical Presentation of False Layer Syndrome Caused by Infectious Bronchitis. In Proceedings of the AVMA/American Association of Avian Pathology Annual Meeting, Denver, CO, USA, 13–17 July 2018. [Google Scholar]
- Grgic, H.; Hunter, D.B.; Hunton, P.; Nagy, E. Pathogenicity of infectious bronchitis virus isolates from Ontario chickens. Can. J. Vet. Res. 2008, 72, 403–410. [Google Scholar] [PubMed]
- Martin, E.A.; Brash, M.L.; Hoyland, S.K.; Coventry, J.M.; Sandrock, C.; Guerin, M.T.; Ojkic, D. Genotyping of infectious bronchitis viruses identified in Canada between 2000 and 2013. Avian Pathol. 2014, 43, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, A.; Popowich, S.; De Silva Senapathi, U.; Abdul-Cader, M.S.; Marshall, F.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate. Viruses 2018, 10, 437. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, A.; De Silva Senapathi, U.; Abdul-Cader, M.S.; Popowich, S.; Marshall, F.; Cork, S.C.; van der Meer, F.; Gomis, S.; Abdul-Careem, M.F. Comparative features of infections of two Massachusetts (Mass) infectious bronchitis virus (IBV) variants isolated from Western Canadian layer flocks. BMC Vet. Res. 2018, 14, 391. [Google Scholar] [CrossRef]
- Kameka, A.M.; Haddadi, S.; Kim, D.S.; Cork, S.C.; Abdul-Careem, M.F. Induction of innate immune response following infectious bronchitis corona virus infection in the respiratory tract of chickens. Virology 2014, 450–451, 114–121. [Google Scholar] [CrossRef]
- Grgic, H.; Hunter, D.B.; Hunton, P.; Nagy, E. Vaccine efficacy against Ontario isolates of infectious bronchitis virus. Can. J. Vet. Res. 2009, 73, 212–216. [Google Scholar]
- Smati, R.; Silim, A.; Guertin, C.; Henrichon, M.; Marandi, M.; Arella, M.; Merzouki, A. Molecular characterization of three new avian infectious bronchitis virus (IBV) strains isolated in Quebec. Virus Genes 2002, 25, 85–93. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Goraichuk, I.V.; Kulkarni, A.B.; Williams-Coplin, D.; Suarez, D.L.; Afonso, C.L. First Complete Genome Sequence of Currently Circulating Infectious Bronchitis Virus Strain DMV/1639 of the GI-17 Lineage. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Holmes, E.C. Evolution and Emergence of RNA Viruses; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Gelb, J., Jr.; Ladman, B.S.; Pope, C.R.; Ruano, J.M.; Brannick, E.M.; Bautista, D.A.; Coughlin, C.M.; Preskenis, L.A. Characterization of nephropathogenic infectious bronchitis virus DMV/1639/11 recovered from Delmarva broiler chickens in 2011. Avian Dis. 2012, 57, 65–70. [Google Scholar] [CrossRef]
- McKinley, E.T.; Hilt, D.A.; Jackwood, M.W. Avian coronavirus infectious bronchitis attenuated live vaccines undergo selection of subpopulations and mutations following vaccination. Vaccine 2008, 26, 1274–1284. [Google Scholar] [CrossRef]
- Reddy, V.R.; Theuns, S.; Roukaerts, I.D.; Zeller, M.; Matthijnssens, J.; Nauwynck, H.J. Genetic Characterization of the Belgian Nephropathogenic Infectious Bronchitis Virus (NIBV) Reference Strain B1648. Viruses 2015, 7, 4488–4506. [Google Scholar] [CrossRef]
- Feng, K.Y.; Chen, T.; Zhang, X.; Shao, G.M.; Cao, Y.; Chen, D.K.; Lin, W.C.; Chen, F.; Xie, Q.M. Molecular characteristic and pathogenicity analysis of a virulent recombinant avain infectious bronchitis virus isolated in China. Poult. Sci. 2018, 97, 3519–3531. [Google Scholar] [CrossRef] [PubMed]
- Abozeid, H.H.; Paldurai, A.; Khattar, S.K.; Afifi, M.A.; El-Kady, M.F.; El-Deeb, A.H.; Samal, S.K. Complete genome sequences of two avian infectious bronchitis viruses isolated in Egypt: Evidence for genetic drift and genetic recombination in the circulating viruses. Infect. Genet. Evol. 2017, 53, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Hewson, K.A.; Ignjatovic, J.; Browning, G.F.; Devlin, J.M.; Noormohammadi, A.H. Infectious bronchitis viruses with naturally occurring genomic rearrangement and gene deletion. Arch. Virol. 2011, 156, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Mardani, K.; Noormohammadi, A.H.; Hooper, P.; Ignjatovic, J.; Browning, G.F. Infectious bronchitis viruses with a novel genomic organization. J. Virol. 2008, 82, 2013–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Zhang, Q.; Chen, J.; Han, Z.; Shao, Y.; Kong, X.; Tong, G. Identification of the avian infectious bronchitis coronaviruses with mutations in gene 3. Gene 2008, 412, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Van Beurden, S.J.; Berends, A.J.; Kramer-Kuhl, A.; Spekreijse, D.; Chenard, G.; Philipp, H.C.; Mundt, E.; Rottier, P.J.M.; Verheije, M.H. Recombinant live attenuated avian coronavirus vaccines with deletions in the accessory genes 3ab and/or 5ab protect against infectious bronchitis in chickens. Vaccine 2018, 36, 1085–1092. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Geng, H.; Deng, Y.; Huang, B.; Guo, Y.; Zhao, Z.; Tan, W. The structural and accessory proteins M, ORF 4a, ORF 4b, and ORF 5 of Middle East respiratory syndrome coronavirus (MERS-CoV) are potent interferon antagonists. Protein Cell 2013, 4, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Wong, C.K.; Li, P.; Xie, Y. A SARS-CoV protein, ORF-6, induces caspase-3 mediated, ER stress and JNK-dependent apoptosis. Biochim. Biophys. Acta 2008, 1780, 1383–1387. [Google Scholar] [CrossRef]
- Sjaak de Wit, J.J.; Cook, J.K.; van der Heijden, H.M. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef]
- Hughes, L.A.; Savage, C.; Naylor, C.; Bennett, M.; Chantrey, J.; Jones, R. Genetically diverse coronaviruses in wild bird populations of northern England. Emerg. Infect. Dis. 2009, 15, 1091–1094. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A.F.; Ladman, B.S.; Dunn, P.A.; Schneider, A.; Davison, S.; Miller, P.G.; Lu, H.; Weinstock, D.; Salem, M.; Eckroade, R.J.; et al. Nephropathogenic infectious bronchitis in Pennsylvania chickens 1997–2000. Avian Dis. 2002, 46, 847–858. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Thor, S.W.; Hilt, D.A.; Kissinger, J.C.; Paterson, A.H.; Jackwood, M.W. Recombination in avian gamma-coronavirus infectious bronchitis virus. Viruses 2011, 3, 1777–1799. [Google Scholar] [CrossRef] [PubMed]
- Khanh, N.P.; Tan, S.W.; Yeap, S.K.; Satharasinghe, D.A.; Hair-Bejo, M.; Bich, T.N.; Omar, A.R. Molecular Characterization of QX-Like and Variant Infectious Bronchitis Virus Strains in Malaysia Based on Partial Genomic Sequences Comprising the S-3a/3b-E-M-Intergenic Region-5a/5b-N Gene Order. Avian Dis. 2017, 61, 442–452. [Google Scholar] [CrossRef]
- Naguib, M.M.; Hoper, D.; Arafa, A.S.; Setta, A.M.; Abed, M.; Monne, I.; Beer, M.; Harder, T.C. Full genome sequence analysis of a newly emerged QX-like infectious bronchitis virus from Sudan reveals distinct spots of recombination. Infect. Genet. Evol. 2016, 46, 42–49. [Google Scholar] [CrossRef]
- Xu, L.; Ren, M.; Sheng, J.; Ma, T.; Han, Z.; Zhao, Y.; Sun, J.; Liu, S. Genetic and biological characteristics of four novel recombinant avian infectious bronchitis viruses isolated in China. Virus Res. 2019, 263, 87–97. [Google Scholar] [CrossRef]
- McKinley, E.T.; Jackwood, M.W.; Hilt, D.A.; Kissinger, J.C.; Robertson, J.S.; Lemke, C.; Paterson, A.H. Attenuated live vaccine usage affects accurate measures of virus diversity and mutation rates in avian coronavirus infectious bronchitis virus. Virus Res. 2011, 158, 225–234. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IBV Isolate | Type of Sample | Commodity | Age | Size of the Flock | Main Complaint |
|---|---|---|---|---|---|
| IBV/Ck/Can/17-035614 | Cecal tonsils | Layers | 6 weeks | 100,000 | Uneven flock |
| IBV/Ck/Can/17-036989 | Kidneys | Layers | 22 weeks | NA | Drop in egg production |
| IBV/Ck/Can/18-048192T | Trachea | Broilers | NA | NA | NA |
| IBV/Ck/Can/18-048430 | Tracheal swab | Broilers | 34 days | 9408 | Routine sampling |
| IBV/Ck/Can/18-049707 | Cecal tonsils | Broiler breeders | 41 weeks | 20,358 | Increased mortality |
| Nucleotide Identity (%) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 18-048192T | 18-049707 | 18-048430 | 17-036989 | 17-035614 | DMV/1639 | PA/Wolg/98 | Qu-mv | Conn | Mass | 4/91 | CAL1737 | CU_82792 | DE/072 | ||
| Amino Acid Identity (%) | 18-048192T | 99.5 | 99.8 | 99.4 | 99.7 | 96.5 | 90.1 | 78.8 | 77.2 | 77.2 | 76.0 | 76.5 | 56.4 | 55.8 | |
| 18-049707 | 99.3 | 99.6 | 99.9 | 99.7 | 96.5 | 90.1 | 78.6 | 77.2 | 77.2 | 75.8 | 76.2 | 56.3 | 55.6 | ||
| 18-048430 | 99.6 | 99.3 | 99.5 | 99.8 | 96.4 | 89.9 | 78.7 | 77.2 | 77.2 | 75.3 | 76.4 | 56.4 | 55.8 | ||
| 17-036989 | 99.1 | 99.8 | 99.1 | 99.6 | 96.4 | 90.0 | 78.5 | 77.2 | 77.2 | 75.7 | 75.5 | 55.3 | 55.6 | ||
| 17-035614 | 99.4 | 99.4 | 99.6 | 99.3 | 96.7 | 89.9 | 78.6 | 77.3 | 77.3 | 75.8 | 76.1 | 56.3 | 55.6 | ||
| DMV/1639 | 94.8 | 95.2 | 95.0 | 95.0 | 95.3 | 91.1 | 79.7 | 78.4 | 78.4 | 76.2 | 75.1 | 56.0 | 55.3 | ||
| PA/Wolg/98 | 87.5 | 87.3 | 87.1 | 87.1 | 87.1 | 87.5 | 79.6 | 78.6 | 78.6 | 76.3 | 76.1 | 56.5 | 56.0 | ||
| Qu-mv | 76.3 | 76.1 | 76.1 | 75.9 | 75.9 | 75.9 | 76.1 | 78.6 | 79.3 | 77.0 | 75.9 | 57.2 | 56.8 | ||
| Conn 46 | 75.5 | 76.9 | 75.6 | 75.5 | 75.6 | 75.3 | 75.7 | 74.3 | 95.1 | 77.6 | 80.3 | 58.7 | 57.9 | ||
| M41 | 73.0 | 73.3 | 73.2 | 73.2 | 73.3 | 73.4 | 73.9 | 74.2 | 90.3 | 78.3 | 80.6 | 58.6 | 57.9 | ||
| 4/91 | 75.5 | 75.5 | 75.3 | 75.3 | 75.3 | 74.7 | 74.4 | 75.1 | 73.9 | 73.8 | 77.2 | 56.2 | 55.8 | ||
| CAL1737 | 72.8 | 72.8 | 72.7 | 72.7 | 72.8 | 71.2 | 71.6 | 72.3 | 77.6 | 78.3 | 74.4 | 57.6 | 57.2 | ||
| CU_82792 | 45.2 | 45.5 | 45.4 | 45.5 | 45.4 | 44.6 | 46.1 | 48..0 | 48.4 | 49.3 | 45.3 | 47.1 | 96.3 | ||
| DE/072 | 45.7 | 45.9 | 45.9 | 45.9 | 45.9 | 45.0 | 46.6 | 47.8 | 48.8 | 49.5 | 45.5 | 48.0 | 93.6 | ||
| IBV/Ck/Can/17-035614 | IBV/Ck/Can/17-036989 | IBV/Ck/Can/18-048192T | IBV/Ck/Can/18-048430 | IBV/Ck/Can/18-049707 | |
|---|---|---|---|---|---|
| 5′UTR | 1–530 (530 nt) | 1–528 (528 nt) | 1–528 (528 nt) | 1–528 (528 nt) | 1–528 (528 nt) |
| 1a | 531–12,419 (11889 nta-3962 aab) | 529–12,417 (11889 nt-3962 aa) | 529–12,360 (11832 nt-3943 aa) | 529–12,393 (11865 nt-3954 aa) | 529–12,417 (11889 nt-3962 aa) |
| 1ab | 531–20,452 (19922 nt-6639 aa) | 529–20,450 (19922 nt-6639 aa) | 529–20,393 (19865 nt-6620 aa) | 529–20,426 (19898 nt-6631 aa) | 529–20,450 (19922 nt-6639 aa) |
| S | 20,403–23,903 (3501 nt-1166 aa) | 20,401–23,901 (3501 nt-1166 aa) | 20,344–23,844 (3501 nt-1166 aa) | 20,377–23,877 (3501 nt-1166 aa) | 20,401–23,901 (3501 nt-1166 aa) |
| 3a | 23,903–24,076 (174 nt-57 aa) | 23,901–24,074 (174 nt-57 aa) | 23,844–24,017 (174 nt-57 aa) | 23,877–24,050 (174 nt-57 aa) | 23,901–24,074 (174 nt-57 aa) |
| 3b | 24,076–24,270 (195 nt-64 aa) | 24,074–24,268 (195 nt-64 aa) | 24,017–24,211 (195 nt-64 aa) | 24,050–24,244 (195 nt-64 aa) | 24,074–24,268 (195 nt-64 aa) |
| E | 24,251–24,580 (330 nt-109 aa) | 24,249–24,578 (330 nt-109 aa) | 24,192–24,521 (330 nt-109 aa) | 24,225–24,554 (330 nt-109 aa) | 24,249–24,578 (330 nt-109 aa) |
| M | 24,552–25,229 (678 nt-225 aa) | 24,550–25,227 (678 nt-225 aa) | 24,493–25,170 (678 nt-225 aa) | 24,526–25,203 (678 nt-225 aa) | 24,550–25,227 (678 nt-225 aa) |
| 4b | 25,230–25,514 (285 nt-94 aa) | 25,228–25,512 (285 nt-94 aa) | 25,171–25,455 (285 nt-94 aa) | 25,204–25,488 (285 nt-94 aa) | 25,228–25,512 (285 nt-94 aa) |
| 4c | 25,435–25,605 (171 nt-56 aa) | 25,433–25,603 (171 nt-56 aa) | 25,376–25,546 (171 nt-56 aa) | 25,409–25,579 (171 nt-56 aa) | 25,433–25,603 (171 nt-56 aa) |
| 5a | 25,589–25,786 (198 nt-65 aa) | 25,587–25,784 (198 nt-65 aa) | 25,530–25,727 (198 nt-65 aa) | 25,563–25,760 (198 nt-65 aa) | 25,587–25,784 (198 nt-65 aa) |
| 5b | 25,783–26,031 (249 nt-82 aa) | 25,781–26,029 (249 nt-82 aa) | 25,724–25,972 (249 nt-82 aa) | 25,757–26,005 (249 nt-82 aa) | 25,781–26,029 (249 nt-82 aa) |
| N | 25,974–27,203 (1230 nt-409 aa) | 25,972–27,201 (1230 nt-409 aa) | 25,915–27,144 (1230 nt-409 aa) | 25,948–27,177 (1230 nt-409 aa) | 25,972–27,201 (1230 nt-409 aa) |
| 6b | 27,212–27,436 (225 nt-74 aa) | 27,210–27,434 (225 nt-74 aa) | 27,153–27296 (144 nt-47 aa) | 27,186–27,410 (225 nt-74 aa) | 27210–27,434 (225 nt-74 aa) |
| 3′UTR | 27,437–27,710 (274 nt) | 27,435–27,708 (274 nt) | 27,297–27,638 (342 nt) | 27,411–27,684 (274 nt) | 27,435–27,708 (274 nt) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.S.H.; Ojkic, D.; Coffin, C.S.; Cork, S.C.; van der Meer, F.; Abdul-Careem, M.F. Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Variants Isolated in Eastern Canada Show Evidence of Recombination. Viruses 2019, 11, 1054. https://doi.org/10.3390/v11111054
Hassan MSH, Ojkic D, Coffin CS, Cork SC, van der Meer F, Abdul-Careem MF. Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Variants Isolated in Eastern Canada Show Evidence of Recombination. Viruses. 2019; 11(11):1054. https://doi.org/10.3390/v11111054
Chicago/Turabian StyleHassan, Mohamed S. H., Davor Ojkic, Carla S. Coffin, Susan C. Cork, Frank van der Meer, and Mohamed Faizal Abdul-Careem. 2019. "Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Variants Isolated in Eastern Canada Show Evidence of Recombination" Viruses 11, no. 11: 1054. https://doi.org/10.3390/v11111054