A Novel Enzootic Nasal Tumor Virus Circulating in Goats from Southern China

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Description
2.2. Detection and Complete Genome Amplification of ENTV-2
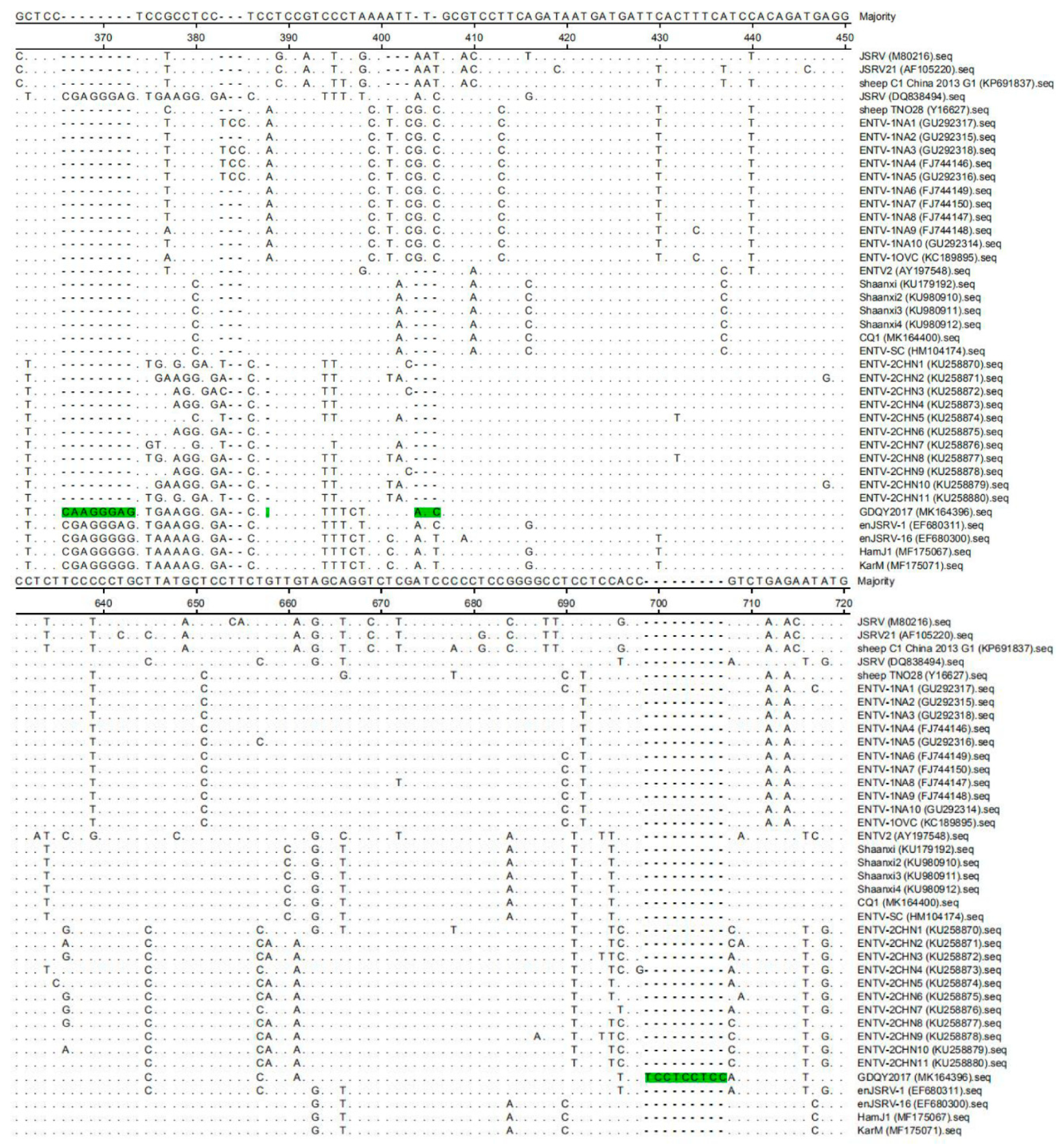
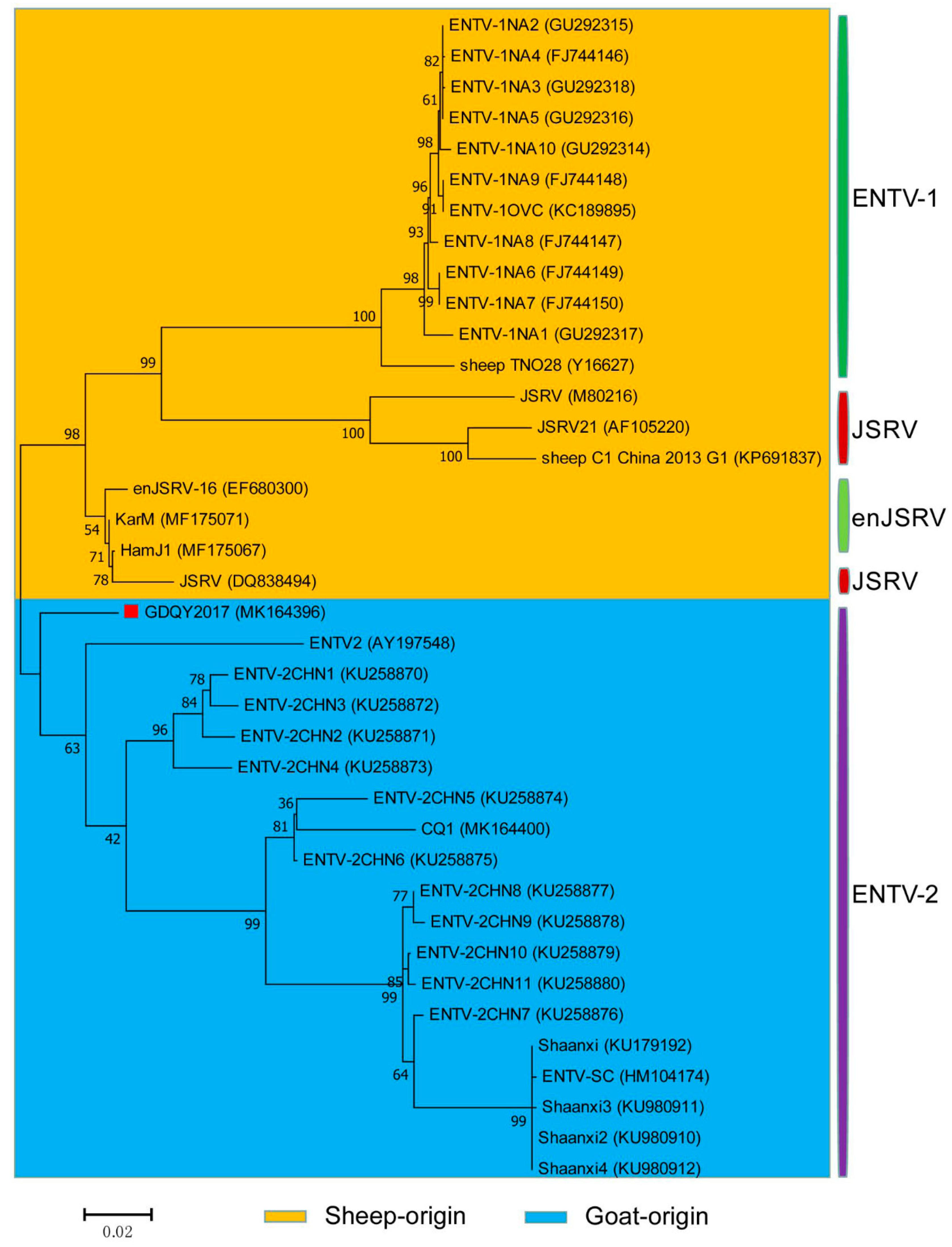
2.3. Sequence and Phylogenetic Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Palmarini, M.; Sharp, J.; de las Heras, M.; Fan, H. Jaagsiekte sheep retrovirus is necessary and sufficient to induce a contagious lung cancer in sheep. J. Virol. 1999, 73, 6964–6972. [Google Scholar] [PubMed]
- Cousens, C.; Minguijon, E.; Dalziel, R.G.; Ortin, A.; Garcia, M.; Park, J.; Gonzalez, L.; Sharp, J.M.; de las Heras, M. Complete sequence of enzootic nasal tumor virus, a retrovirus associated with transmissible intranasal tumors of sheep. J. Virol. 1999, 73, 3986–3993. [Google Scholar] [PubMed]
- Ortín, A.; Cousens, C.; Minguijón, E.; Pascual, Z.; Villarreal, M.P.; Sharp, J.M.; Heras Mde, L. Characterization of enzootic nasal tumour virus of goats: Complete sequence and tissue distribution. J. Gen. Virol. 2003, 84, 2245–2252. [Google Scholar] [CrossRef]
- De las Heras, M.; García de Jalón, J.A.; Minguijón, E.; Gray, E.W.; Dewar, P.; Sharp, J.M. Experimental transmission of enzootic intranasal tumors of goats. Vet. Pathol. 1995, 32, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.R.; Linnerth-Petrik, N.M.; Yu, D.L.; Foster, R.A.; Menzies, P.I.; Diaz-Méndez, A.; Chalmers, H.J.; Wootton, S.K. Experimental transmission of enzootic nasal adenocarcinoma in sheep. Vet. Res. 2013, 44, 66. [Google Scholar] [CrossRef]
- Liu, F.X.; Lin, X.; Huang, F.; Hao, X.P.; Zhao, Z.H.; Yu, W.S.; Wang, H.Z. An investigation and pathologic morphological study on tumors of domestic animals in Inner Mongolia. Vet. Ori. 1984, 3, 15–25. (in Chinese). [Google Scholar]
- Lei, H.Y.; Su, J.M.; Ning, L.Z.; Kang, Y.; Chen, K.Y.; Zeng, D.N. Investigation and diagnosis of an enzootic nasal tumor of goat. Prog. Vet. Med. 2006, 27, 112–114. (in Chinese). [Google Scholar]
- Yi, G.; Kaiyu, W.; Qigui, Y.; Zhongqiong, Y.; Yingdong, Y.; Defang, C.; Jinlu, H. Descriptive study of enzootic nasal adenocarcinoma in goats in southwestern China. Transbound. Emerg. Dis. 2010, 57, 197–200. [Google Scholar] [CrossRef]
- Liu, F.; Feng, Y.C.; Yan, Q.G.; Han, G.Q. Diagnosis of an enzootic nasal tumor of goat in Sichuan. Anim. Husb. Vet. Med. 2011, 43, 83–86. (in Chinese). [Google Scholar]
- Yu, Y.D.; Wei, L.F.; Huang, X.J.; Zhang, B.; Yu, X.H.; Tang, C. Diagnosis of four cases of enzootic nasal adenocarcinoma of goats. Prog. Vet. Med. 2014, 35, 129–131. (in Chinese). [Google Scholar]
- Jiang, J.X.; Lin, Y.S.; Jiang, B.; Mao, K.M.; You, W.; Zhang, J.P.; Hu, Q.L. Molecular epidemiology of enzootic nasal tumor virus on goats in Fujian. Fujian J. Agr. Sci. 2017, 32, 837–841. (in Chinese). [Google Scholar]
- Hou, H.; Zhu, D.; Zhang, D.; Hu, X.; Zhao, R.; Dai, Y. Cloning and Analysis of gag Gene of Enzootic Nasal Tumor Virus in Goats. Chin. Herb. Sci. 2018, 38, 53–55. (in Chinese). [Google Scholar]
- He, Y.; Zhang, Q.; Wang, J.; Zhou, M.; Fu, M.; Xu, X. Full-length genome sequence analysis of enzootic nasal tumor virus isolated from goats in China. Virol. J. 2017, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Huang, Q.; Chen, T.; Jiang, J.; Hou, F.; Xu, D.; Peng, Y.; Fang, R.; Chen, J. First detection and genotypic analysis of goat enzootic nasal tumor virus 2 in Chongqing, China. Arch. Virol. 2019, 164, 1647–1650. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.R.; Linnerth-Petrik, N.M.; Laporte, A.N.; Menzies, P.I.; Foster, R.A.; Wootton, S.K. Full-length genome sequence analysis of enzootic nasal tumor virus reveals an unusually high degree of genetic stability. Virus Res. 2010, 151, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Hallwirth, C.; Maeda, N.; York, D.; Fan, H. Variable regions 1 and 2 (VR1 and VR2) in JSRV gag are not responsible for the endogenous JSRV particle release defect. Virus Genes. 2005, 30, 59–68. [Google Scholar] [CrossRef]
- Mura, M.; Murcia, P.; Caporale, M.; Spencer, T.E.; Nagashima, K.; Rein, A.; Palmarini, M. Late viral interference induced by transdominant Gag of an endogenous retrovirus. Proc. Natl. Acad. Sci. USA 2004, 101, 11117–11122. [Google Scholar] [CrossRef] [Green Version]
- Palmarini, M.; Maeda, N.; Murgia, C.; De-Fraja, C.; Hofacre, A.; Fan, H. A phosphatidylinositol 3-kinase docking site in the cytoplasmic tail of the Jaagsiekte sheep retrovirus transmembrane protein is essential for envelope-induced transformation of NIH 3T3 cells. J. Virol. 2001, 75, 11002–11009. [Google Scholar] [CrossRef]
- Caporale, M.; Martineau, H.; De las Heras, M.; Murgia, C.; Huang, R.; Centorame, P.; Di Francesco, G.; Di Gialleonardo, L.; Spencer, T.E.; Griffiths, D.J.; et al. Host species barriers to Jaagsiekte sheep retrovirus replication and carcinogenesis. J. Virol. 2013, 87, 10752–10762. [Google Scholar] [CrossRef]
- De las Heras, M.; Garcia de Jalon, J.A.; Sharp, J.M. Pathology of enzootic intranasal tumor in thirty-eight goats. Vet. Pathol. 1991, 28, 474–481. [Google Scholar] [CrossRef]
- Vitellozzi, G.; Mughetti, L.; Palmarini, M.; Mandara, M.T.; Mechelli, L.; Sharp, J.M.; Manocchio, I. Enzootic intranasal tumour of goats in Italy. J. Vet. Med. Ser. B 1993, 40, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.R.; Gerpe, M.C.; Wootton, S.K. Construction of a molecular clone of ovine enzootic nasal tumor virus. Virol. J. 2016, 13, 209. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence(5′-3′) | Location | Product Length |
|---|---|---|---|
| 1F | ACAAGGCATCAGCCATTTTGGTCTGATCCTCTCAACCCCA | 1–40 | |
| 1R | AGGAGGAGGAGCATCATAACCAGGCTCTGGGTCAGGAATA | 627–588 | 627 bp |
| 2F | GTTTTCCTCGCCACTACTCTTG | 151–172 | |
| 2R | TACCCAATAAGCGTCGGATGAT | 2399–2378 | 2249 bp |
| 3F | CACTCCTAATTTGTGCCCACG | 1848–1869 | |
| 3R | GGCCACTGATCGACCCATAC | 2934–2915 | 1087 bp |
| 4F | GAAGAGGTTTGGGGTGTTTTCCCTAGGGACCTCTGATTCTCCTGTGAC | 2830–2878 | |
| 4R | GTTTAAGACGTTGATGAGCTCGTTCTACAATCCCTTGTCCCTGTGGGT | 5087–5040 | 2258 bp |
| 5F | AGAACGAGCTCATCAACGTCTTAAACATCAACT | 5062–5094 | |
| 5R | CTTGTTGTTTTATTGTGTCATAGTATATAT | 7440–7411 | 2379 bp |
| Isolation (GenBank No.) | Complete Genome | gag Gene | gag Protein | env Gene | env Protein |
|---|---|---|---|---|---|
| JSRV (M80216) | 88.2% | 86.6% | 88.9% | 85.9% | 89.4% |
| JSRV21 (AF105220) | 88.2% | 86.1% | 89.4% | 86.2% | 89.7% |
| sheep C1 China 2013 G1 (KP691837) | 87.6% | 85.7% | 89.1% | 85.8% | 89.2% |
| JSRV (DQ838494) | 92.2% | 93.6% | 87.7% | 93.6% | 92.2% |
| sheep TNO28 (Y16627) | 88.4% | 87.7% | 89.7% | 87.7% | 88.2% |
| ENTV-1NA1 (GU292317) | 88.2% | 87.5% | 89.4% | 87.8% | 87.9% |
| ENTV-1NA2 (GU292315) | 88.2% | 87.9% | 89.4% | 87.7% | 88.2% |
| ENTV-1NA3 (GU292318) | 88.2% | 87.7% | 89.3% | 87.6% | 88.2% |
| ENTV-1NA4 (FJ744146) | 88.2% | 87.8% | 89.3% | 87.6% | 88.1% |
| ENTV-1NA5 (GU292316) | 88.2% | 87.8% | 89.3% | 87.7% | 88.2% |
| ENTV-1NA6 (FJ744149) | 88.4% | 87.9% | 89.4% | 87.8% | 88.1% |
| ENTV-1NA7 (FJ744150) | 88.4% | 87.9% | 89.4% | 87.8% | 88.1% |
| ENTV-1NA8 (FJ744147) | 88.3% | 87.8% | 89.6% | 87.8% | 88.1% |
| ENTV-1NA9 (FJ744148) | 88.3% | 87.8% | 89.6% | 87.7% | 88.2% |
| ENTV-1NA10 (GU292314) | 88.2% | 88% | 89.6% | 87.5% | 88.1% |
| ENTV-1OVC (KC189895) | 88.2% | 87.7% | 89.4% | 87.7% | 88.2% |
| ENTV2 (AY197548) | 88.2% | 87.2% | 88.6% | 91.2% | 89.8% |
| Shaanxi (KU179192) | 91.9% | 86.7% | 88.7% | 89.1% | 91% |
| Shaanxi2 (KU980910) | 92% | 86.6% | 88.7% | 89.1% | 91% |
| Shaanxi3 (KU980911) | 92.2% | 87.2% | 88.9% | 89.2% | 91.1% |
| Shaanxi4 (KU980912) | 91.8% | 86.7% | 88.7% | 89.1% | 91% |
| CQ1 (MK164400) | 92.7% | 86.6% | 88.3% | 90.8% | 90.5% |
| ENTV-SC (HM104174) | 90.9% | 86.5% | 88.3% | 89% | 90.8% |
| ENTV-2CHN1 (KU258870) | 95.7% | 96.3% | 94.9% | 93.3% | 92.4% |
| ENTV-2CHN2 (KU258871) | 96% | 95.8% | 93.3% | 93.3% | 91.6% |
| ENTV-2CHN3 (KU258872) | 95.8% | 95.9% | 94.1% | 93.5% | 92.2% |
| ENTV-2CHN4 (KU258873) | 96.6% | 96.8% | 94.9% | 94.8% | 93.7% |
| ENTV-2CHN5 (KU258874) | 95.2% | 96% | 93.5% | 91.7% | 91.6% |
| ENTV-2CHN6 (KU258875) | 94.7% | 96.8% | 95.1% | 92.1% | 93.1% |
| ENTV-2CHN7 (KU258876) | 94.4% | 95.5% | 95.1% | 90.3% | 93.9% |
| ENTV-2CHN8 (KU258877) | 95% | 96.6% | 94.9% | 90.4% | 93.5% |
| ENTV-2CHN9 (KU258878) | 92.6% | 95.9% | 93.3% | 90.3% | 93.4% |
| ENTV-2CHN10 (KU258879) | 93.3% | 96.1% | 93.6% | 90.1% | 93.2% |
| ENTV-2CHN11 (KU258880) | 92.2% | 95.6% | 95.1% | 90.3% | 93.4% |
| enJSRV-1 (EF680311) | 94.6% | 95.1% | 94.5% | -a | -a |
| enJSRV-16 (EF680300) | 93.3% | 91.6% | 91.4% | 94.6% | 94.3% |
| HamJ1 (MF175067) | 93.6% | 91.8% | 91.7% | 94.9% | 94.4% |
| KarM (MF175071) | 93.6% | 91.8% | 91.7% | 95% | 94.6% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, S.-L.; Lv, D.-H.; Xu, Z.-H.; Yu, J.-S.; Wen, X.-H.; Zhang, H.; Chen, Q.-L.; Jia, C.-L.; Zhou, X.-R.; Zhai, Q.; et al. A Novel Enzootic Nasal Tumor Virus Circulating in Goats from Southern China. Viruses 2019, 11, 956. https://doi.org/10.3390/v11100956
Zhai S-L, Lv D-H, Xu Z-H, Yu J-S, Wen X-H, Zhang H, Chen Q-L, Jia C-L, Zhou X-R, Zhai Q, et al. A Novel Enzootic Nasal Tumor Virus Circulating in Goats from Southern China. Viruses. 2019; 11(10):956. https://doi.org/10.3390/v11100956
Chicago/Turabian StyleZhai, Shao-Lun, Dian-Hong Lv, Zhi-Hong Xu, Jie-Shi Yu, Xiao-Hui Wen, He Zhang, Qin-Ling Chen, Chun-Ling Jia, Xiu-Rong Zhou, Qi Zhai, and et al. 2019. "A Novel Enzootic Nasal Tumor Virus Circulating in Goats from Southern China" Viruses 11, no. 10: 956. https://doi.org/10.3390/v11100956