Fine Root Biomass and Its Relationship with Aboveground Traits of Larix gmelinii Trees in Northeastern China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
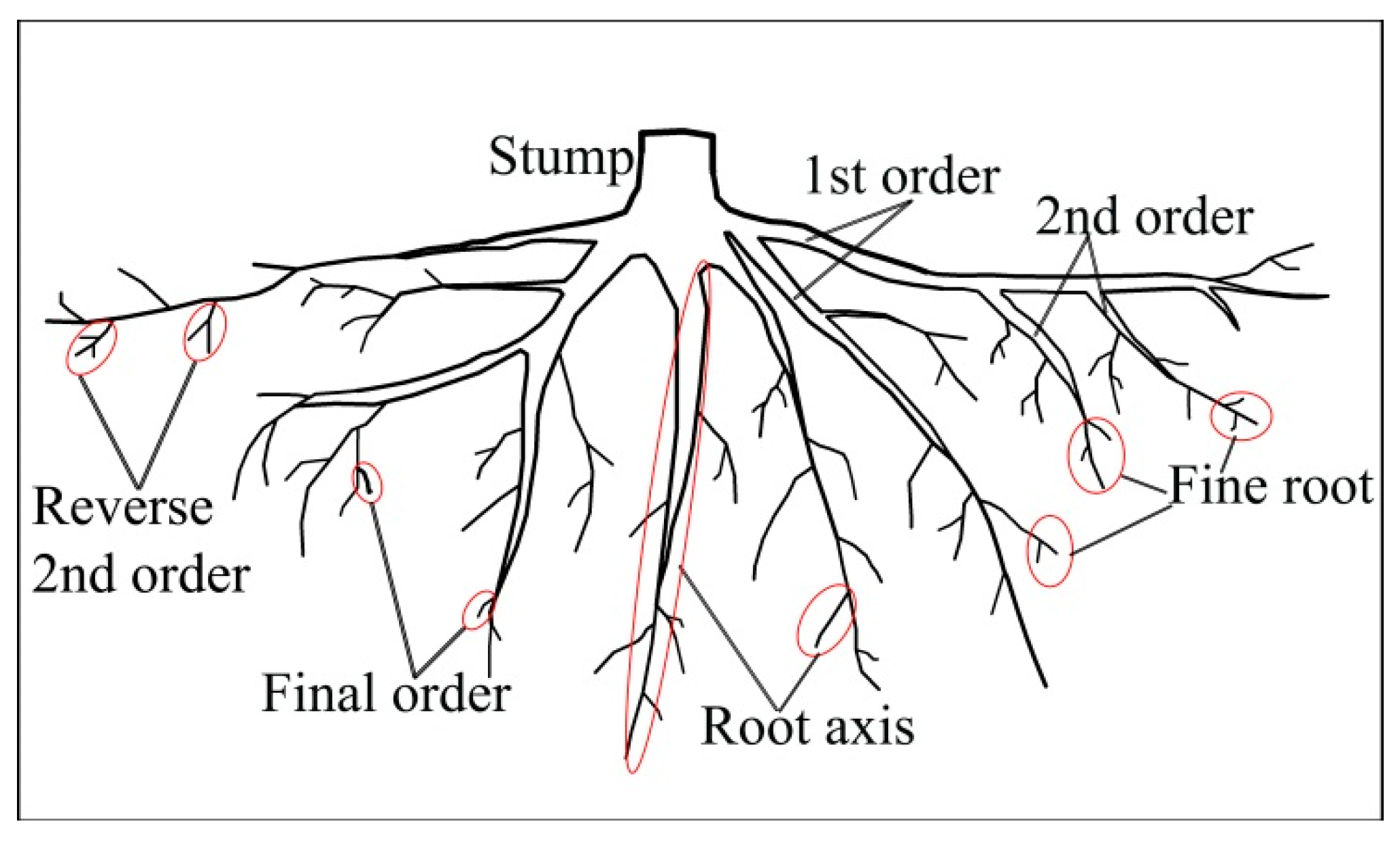
2.2. Measurement and Data Collection
2.3. Step-Fitting Model and Statistical Analyses
3. Results
3.1. Root Branch Biomass and Surface Area Allometrics
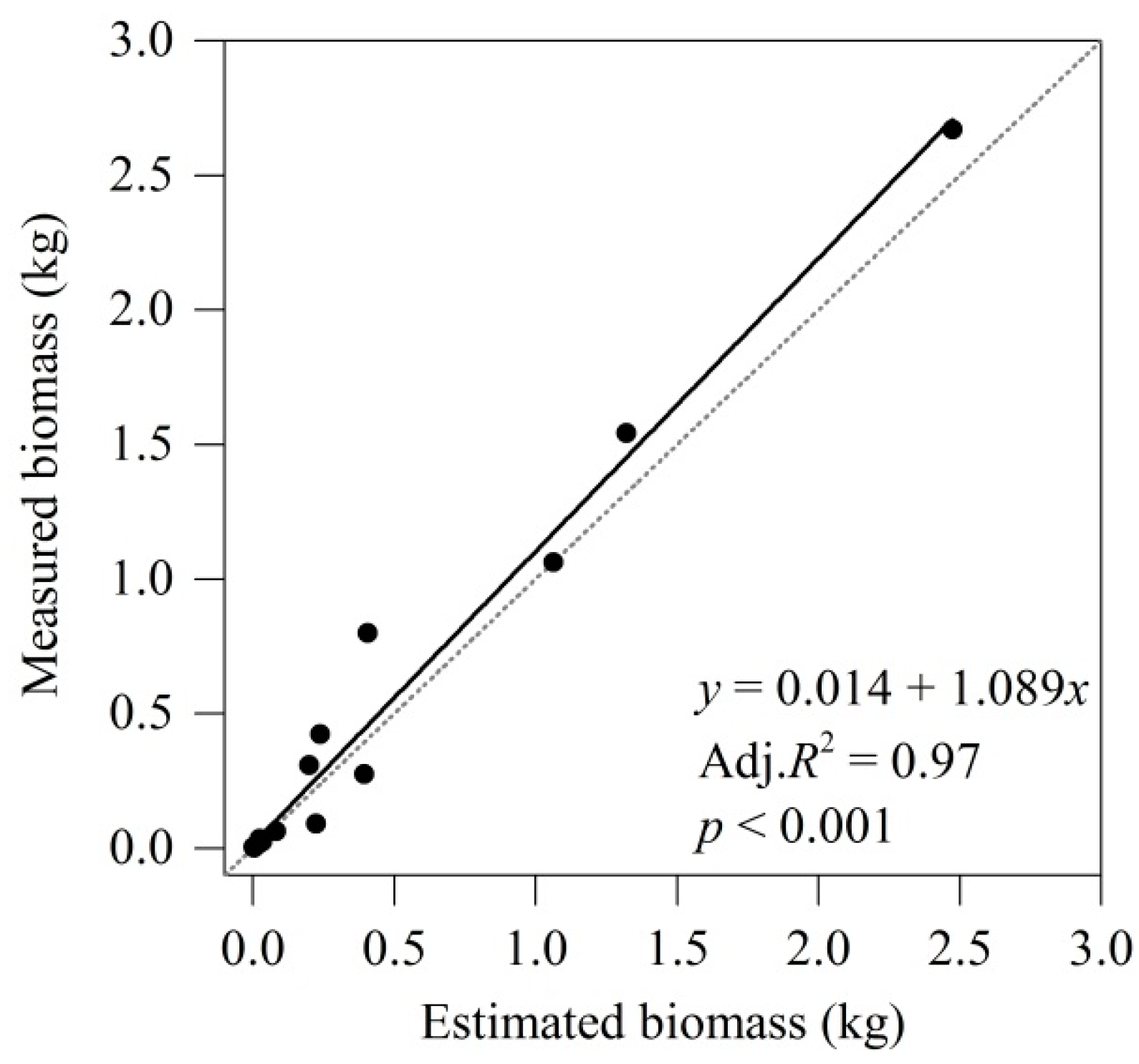
3.2. Validation of Root Branch Biomass Model
3.3. Individual-Tree Biomass and Surface Area Allometrics
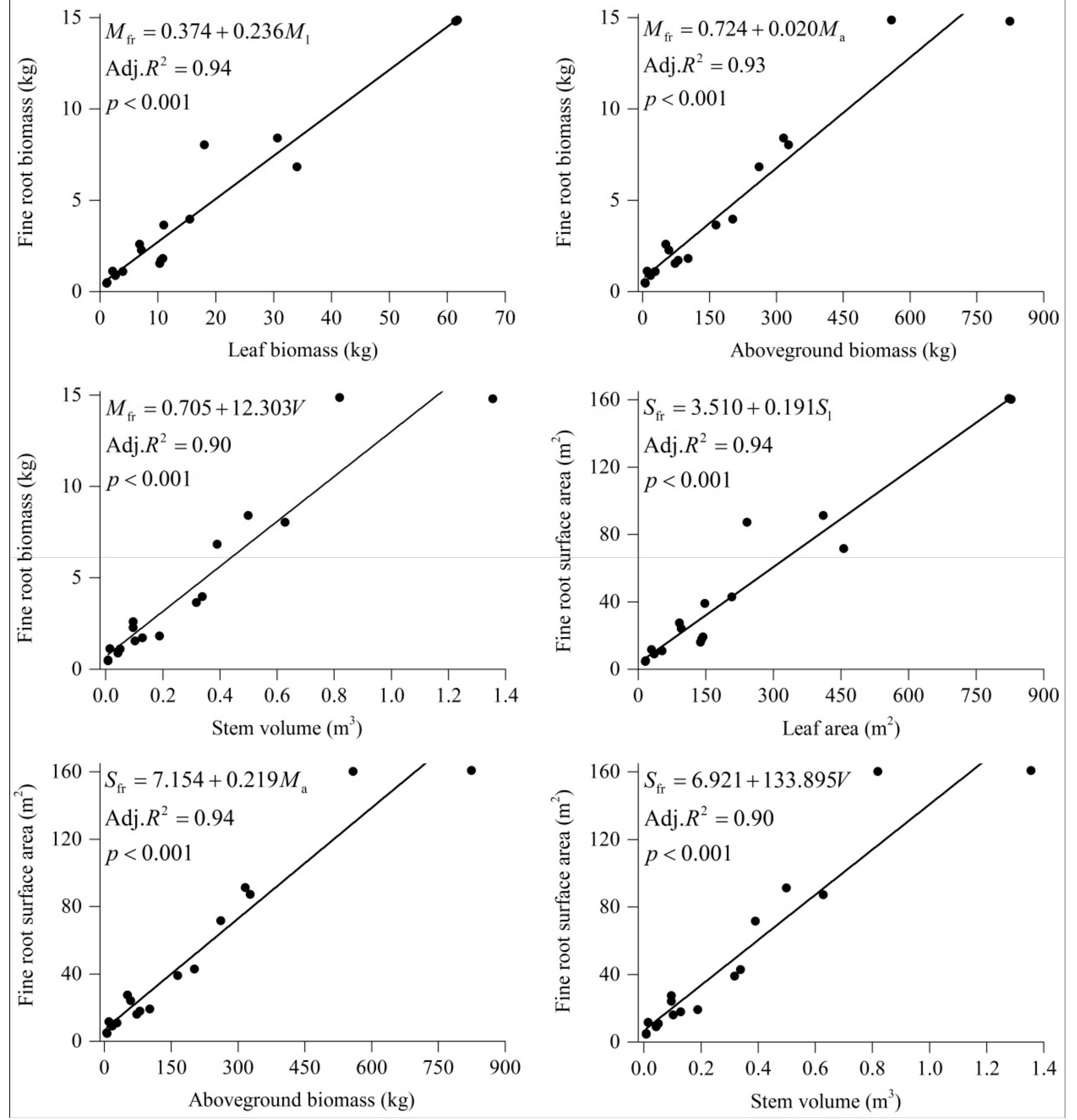
3.4. Fine Root and Aboveground Relationships
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Vogt, K.A.; Vogt, D.J.; Bloomfield, J. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant. Soil 1998, 200, 71–89. [Google Scholar] [CrossRef]
- Silver, W.L.; Thompson, A.W.; McGroddy, M.E.; Varner, R.K.; Dias, J.D.; Silva, H.; Crill, P.M.; Keller, M. Fine root dynamics and trace gas fluxes in two lowland tropical forest soils. Glob. Chang. Biol. 2005, 11, 290–306. [Google Scholar] [CrossRef]
- Hertel, D.; Leuschner, C. A comparison of four different fine root production estimates with ecosystem carbon balance data in a fagus–quercus mixed forest. Plant. Soil 2002, 245, 335. [Google Scholar]
- Brunner, I.; Godbold, D.L. Tree roots in a changing world. J. For. Res. 2007, 12, 78–82. [Google Scholar] [CrossRef]
- Xiao, C.W.; Sang, W.G.; Wang, R.-Z. Fine root dynamics and turnover rate in an asia white birch forest of donglingshan mountain, china. For. Ecol. Manag. 2008, 255, 765–773. [Google Scholar]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Yunusa, I.A.M.; Zolfaghar, S.; Zeppel, M.J.B.; Li, Z.; Palmer, A.R.; Eamus, D. Fine root biomass and its relationship to evapotranspiration in woody and grassy vegetation covers for ecological restoration of waste storage and mining landscapes. Ecosystems 2012, 15, 113–127. [Google Scholar] [CrossRef]
- Van Do, T.; Osawa, A.; Sato, T. Estimation of fine-root production using rates of diameter-dependent root mortality, decomposition and thickening in forests. Tree Physiol. 2016, 36, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Fang, J. Linking belowground and aboveground phenology in two boreal forests in northeast china. Oecologia 2014, 176, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.B. A review of evidence on the control of shoot: Root ratio, in relation to models. Ann. Bot. 1988, 61, 433–449. [Google Scholar] [CrossRef]
- Vanninen, P.; Mäkelä, A. Fine root biomass of scots pine stands differing in age and soil fertility in southern finland. Tree Physiol. 1999, 19, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Cannell, M.G.; Dewar, R. Carbon allocation in trees: A review of concepts for modelling. Adv. Ecol. Res. 1994, 25, 59–104. [Google Scholar]
- O’Grady, A.P.; Worledge, D.; Battaglia, M. Above- and below-ground relationships, with particular reference to fine roots, in a young eucalyptus globulus (labill.) stand in southern tasmania. Trees 2006, 20, 531–538. [Google Scholar] [CrossRef]
- Johansson, T. Biomass production and allometric above- and below-ground relations for young birch stands planted at four spacings on abandoned farmland. Forestry 2007, 80, 41–52. [Google Scholar] [CrossRef]
- Koteen, L.E.; Raz-Yaseef, N.; Baldocchi, D.D. Spatial heterogeneity of fine root biomass and soil carbon in a california oak savanna illuminates plant functional strategy across periods of high and low resource supply. Ecohydrology 2015, 8, 294–308. [Google Scholar] [CrossRef]
- Wilson, S.D.; Kalamees, R. Below-ground opportunities in vegetation science. J. Veg. Sci. 2014, 25, 1117–1125. [Google Scholar] [CrossRef]
- Mommer, L.; Weemstra, M. The role of roots in the resource economics spectrum. New Phytol. 2012, 195, 725. [Google Scholar] [CrossRef] [PubMed]
- Vogt, K.A.; Vogt, D.J.; Palmiotto, P.A.; Boon, P.; O’Hara, J.; Asbjornsen, H. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. Plant. Soil 1996, 187, 159–219. [Google Scholar] [CrossRef]
- Hendricks, J.J.; Hendrick, R.L.; Wilson, C.A.; Mitchell, R.J.; Pecot, S.D.; Guo, D. Assessing the patterns and controls of fine root dynamics: An empirical test and methodological review. J. Ecol. 2006, 94, 40–57. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Ouimet, R.; Camire, C.; Brazeau, M.; Moore, J.-D. Estimation of coarse root biomass and nutrient content for sugar maple, jack pine, and black spruce using stem diameter at breast height. Can. J. For. Res. 2008, 38, 92–100. [Google Scholar] [CrossRef]
- McNicol, I.M.; Berry, N.J.; Bruun, T.B.; Hergoualc’h, K.; Mertz, O.; de Neergaard, A.; Ryan, C.M. Development of allometric models for above and belowground biomass in swidden cultivation fallows of northern laos. For. Ecol. Manag. 2015, 357, 104–116. [Google Scholar] [CrossRef]
- Cole, T.G.; Ewel, J.J. Allometric equations for four valuable tropical tree species. For. Ecol. Manag. 2006, 229, 351–360. [Google Scholar] [CrossRef]
- Liu, Q.J. Nested regression for establishing tree biomass equations. J. Plant. Ecol. 2009, 33, 331–337. (In Chinese) [Google Scholar]
- Jia, Q.Q.; Liu, Q.J.; Li, J.Q. Individual-based fine root biomass and its functional relationship with leaf for Pinus tabuliformis in northern china. Eur. J. For. Res. 2015, 134, 705–714. [Google Scholar] [CrossRef]
- Quan, X.; Wang, C.; Zhang, Q.; Wang, X.; Luo, Y.; Bond-Lamberty, B. Dynamics of fine roots in five chinese temperate forests. J. Plant. Res. 2010, 123, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Li, R.; Wang, H.; He, B. Quantitative assessment of human appropriation of aboveground net primary production in china. Ecol. Model. 2015, 312, 54–60. [Google Scholar] [CrossRef]
- Kajimoto, T. Aboveground net production and dry matter allocation of Pinus pumila forests in the kiso mountain range, central Japan. Ecol. Res. 1994, 9, 193–204. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zhang, L.; Li, F. A compatible system of biomass equations for three conifer species in northeast, china. For. Ecol. Manag. 2014, 329, 306–317. [Google Scholar] [CrossRef]
- Zeng, W.S. Integrated individual tree biomass simultaneous equations for two larch species in northeastern and northern china. Scand. J. For. Res. 2015, 30, 594–604. [Google Scholar] [CrossRef]
- Tyree, M.T.; Velez, V.; Dalling, J.W. Growth dynamics of root and shoot hydraulic conductance in seedlings of five neotropical tree species: Scaling to show possible adaptation to differing light regimes. Oecologia 1998, 114, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Kajimoto, T.; Matsuura, Y.; Osawa, A.; Abaimov, A.P.; Zyryanova, O.A.; Isaev, A.P.; Yefremov, D.P.; Mori, S.; Koike, T. Size-mass allometry and biomass allocation of two larch species growing on the continuous permafrost region in siberia. For. Ecol. Manag. 2006, 222, 314–325. [Google Scholar] [CrossRef]
- Kajimoto, T.; Matsuura, Y.; Sofronov, M.A.; Volokitina, A.V.; Mori, S.; Osawa, A.; Abaimov, A.P. Above- and belowground biomass and primary productivity of a larix gmelinii stand near tura, central siberia. Tree Physiol. 1999, 19, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Jomura, M.; Wang, W.J.; Masyagina, O.V.; Homma, S.; Kanazawa, Y.; Zu, Y.G.; Koike, T. Carbon dynamics of larch plantations in northeastern China and Japan. In Permafrost Ecosystems; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar]
- Kachamba, D.J.; Eid, T.; Gobakken, T. Above- and belowground biomass models for trees in the miombo woodlands of malawi. Forests 2016, 7, 38. [Google Scholar] [CrossRef]
- Cheng, Y. Timing and Depth-Related Patterns of Fine Root Biomass, Production and Turnover in Larix olgensis Henry Plantation. Master’s Thesis, Northeast Forestry University, Harbin, China, 2004. (In Chinese). [Google Scholar]
- Bao, C.; Bai, Y.; Qing, M.; Chen, G.; Zhang, Q.; Wang, L. Study on the Larix gmelinii natural forest biomass carbon storage. J. Inn. Mong. Agric. Univ. 2010, 31, 77–82. (In Chinese) [Google Scholar]
- Wang, C.; Han, S.; Zhou, Y.; Zhang, J.; Zheng, X.; Dai, G.; Li, M.-H. Fine root growth and contribution to soil carbon in a mixed mature Pinus koraiensis forest. Plant. Soil 2016, 400, 275–284. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Q.; Cihlar, J.; Bauhus, J.; Price, D.T. Estimating fine-root biomass and production of boreal and cool temperate forests using aboveground measurements: A new approach. Plant. Soil 2004, 265, 31–46. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Factors causing variation in fine root biomass in forest ecosystems. For. Ecol. Manag. 2011, 261, 265–277. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Derome, J.; Nöjd, P.; Kukkola, M. Fine root biomass in relation to site and stand characteristics in norway spruce and scots pine stands. Tree Physiol. 2007, 27, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.W.; Ceulemans, R. Allometric relationships for below- and aboveground biomass of young scots pines. For. Ecol. Manag. 2004, 203, 177–186. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Definition | Min | Max | Mean | S.D. |
|---|---|---|---|---|---|
| - | Basic density of branch (g/cm3) | 0.40 | 0.52 | 0.46 | 0.05 |
| - | Basic density of stem wood (g/cm3) | 0.39 | 0.51 | 0.50 | 0.03 |
| - | Basic density of stem bark (g/cm3) | 0.35 | 0.39 | 0.37 | 0.01 |
| - | Number of needles per leaf fascicle | 22 | 28 | 26 | 3 |
| - | Leaf fascicle mass (g) | 0.016 | 0.024 | 0.021 | 0.0027 |
| - | Needle area (cm2) | 0.066 | 0.18 | 0.13 | 0.032 |
| - | Short shoot mass (g) | 0.0035 | 0.0075 | 0.0060 | 0.0021 |
| Mtr | Total root biomass per tree (kg) | 1.51 | 222.52 | 51.25 | 67.51 |
| Mfr | Fine root biomass per tree (kg) | 0.46 | 14.86 | 4.38 | 4.66 |
| Sfr | Fine root surface area per tree (m2) | 4.51 | 160.71 | 46.96 | 50.66 |
| Ml | Leaf biomass per tree (kg) | 1.17 | 61.77 | 17.02 | 19.22 |
| Sl | Leaf area per tree (m2) | 15.70 | 827.48 | 227.92 | 257.58 |
| Ma | Aboveground biomass per tree (kg) | 5.33 | 824.17 | 181.59 | 224.13 |
| V | Stem volume per tree (m3) | 0.0078 | 1.36 | 0.30 | 0.36 |
| Component | Application Range (mm) | a | b | RMSE | Adj. R2 | n | p Value |
|---|---|---|---|---|---|---|---|
| Root branch biomass | 1.99 ≤ x ≤ 240 | 337.17 | 1.99 | 1.24 kg | 0.99 | 270 | <0.001 |
| Fine root biomass | 0 < x < 0.34 | 2835.45 | 6.95 | 0.83 mg | 0.92 | 619 | <0.001 |
| 0.34 ≤ x ≤ 2.14 | 95.38 | 3.83 | 0.20 g | 0.97 | 568 | <0.001 | |
| 2.14 ≤ x ≤ 240 | 599.10 | 1.42 | 85.44 g | 0.99 | 270 | <0.001 | |
| Fine root surface area | 0 < x < 0.34 | 89,242.00 | 6.59 | 34.82 mm2 | 0.88 | 619 | <0.001 |
| 0.34 ≤ x ≤ 1.01 | 1944.70 | 3.07 | 1.84 cm2 | 0.87 | 338 | <0.001 | |
| 1.01 < x ≤ 5.48 | 1970.50 | 2.05 | 0.02 m2 | 0.94 | 352 | <0.001 | |
| 5.48 < x ≤ 240 | 5342.30 | 1.46 | 1.04 m2 | 0.99 | 148 | <0.001 |
| Model | RMSE | Adj. R2 | p Value |
|---|---|---|---|
| Mtr = 0.056 dbh 2.230 | 16.47 kg | 0.94 | <0.001 |
| Mfr = 0.016 dbh 1.866 | 0.92 kg | 0.97 | <0.001 |
| Sfr = 0.155 dbh 1.890 | 9.75 m2 | 0.92 | <0.001 |
| Ml = 0.057 dbh 1.876 | 5.76 kg | 0.96 | <0.001 |
| Sl = 0.757 dbh 1.877 | 77.56 m2 | 0.96 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, S.; Jia, Q.; Zhou, G.; Zhou, H.; Liu, Q.; Yu, J. Fine Root Biomass and Its Relationship with Aboveground Traits of Larix gmelinii Trees in Northeastern China. Forests 2018, 9, 35. https://doi.org/10.3390/f9010035
Meng S, Jia Q, Zhou G, Zhou H, Liu Q, Yu J. Fine Root Biomass and Its Relationship with Aboveground Traits of Larix gmelinii Trees in Northeastern China. Forests. 2018; 9(1):35. https://doi.org/10.3390/f9010035
Chicago/Turabian StyleMeng, Shengwang, Quanquan Jia, Guang Zhou, Hua Zhou, Qijing Liu, and Jian Yu. 2018. "Fine Root Biomass and Its Relationship with Aboveground Traits of Larix gmelinii Trees in Northeastern China" Forests 9, no. 1: 35. https://doi.org/10.3390/f9010035