
The Effects of Poplar Plantations on Vascular Plant Diversity in Riparian Landscapes


Abstract
:1. Introduction
- To what extent does vascular plant diversity differ in poplar plantations and riparian forests?
- What are the respective influences of stress and disturbance on vascular plant diversity in riparian landscapes?
2. Material and Methods
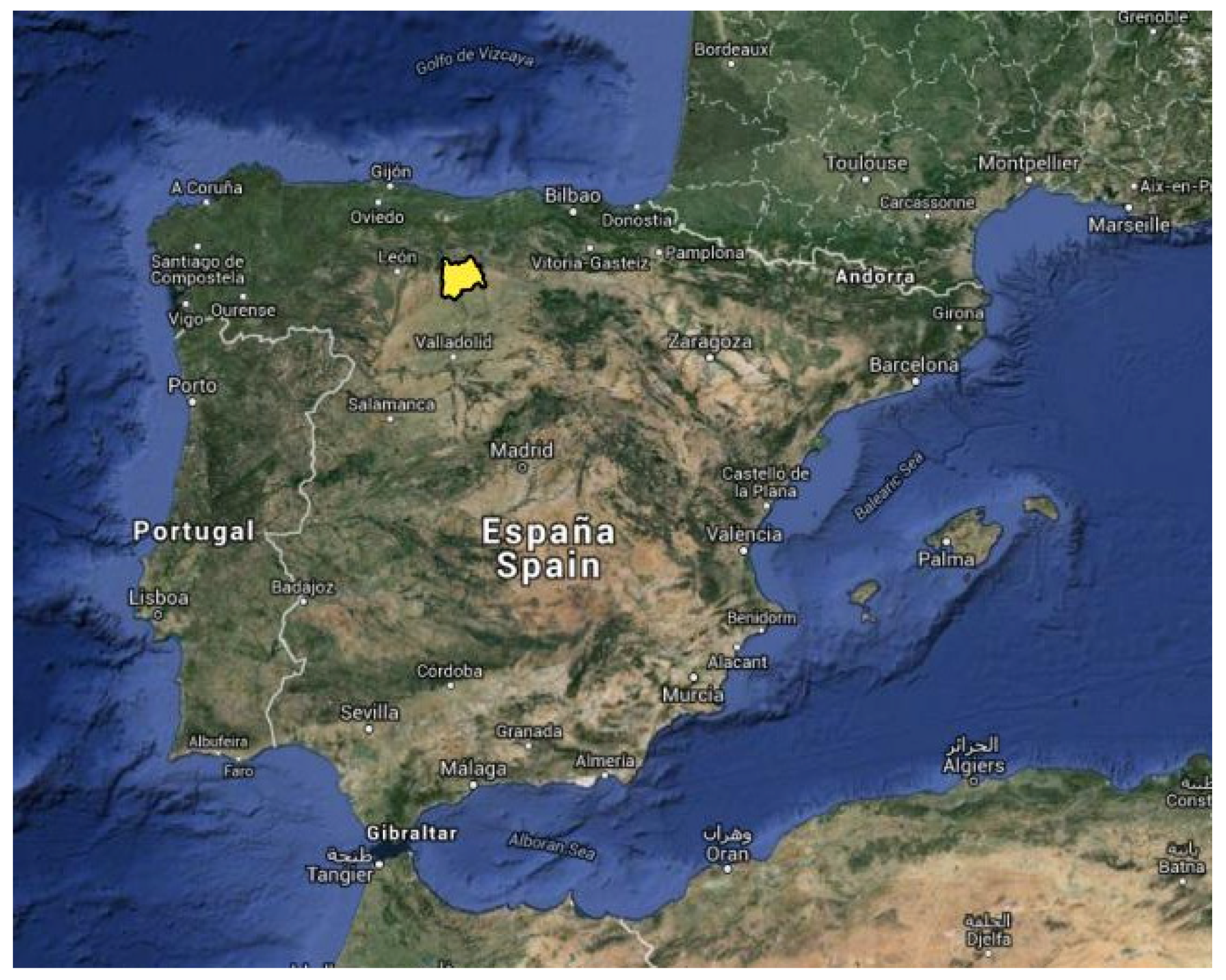
2.1. Site Description and Experimental Design
2.2. Growth Measurements, Assessment of Crown Condition Variables and Soil Sampling
2.3. Vascular Plant Sampling
2.4. Data Analysis
2.4.1. Comparison between Riparian Forests and Poplar Plantations
2.4.2. Comparison of Different Types of Poplar Plantations
3. Results
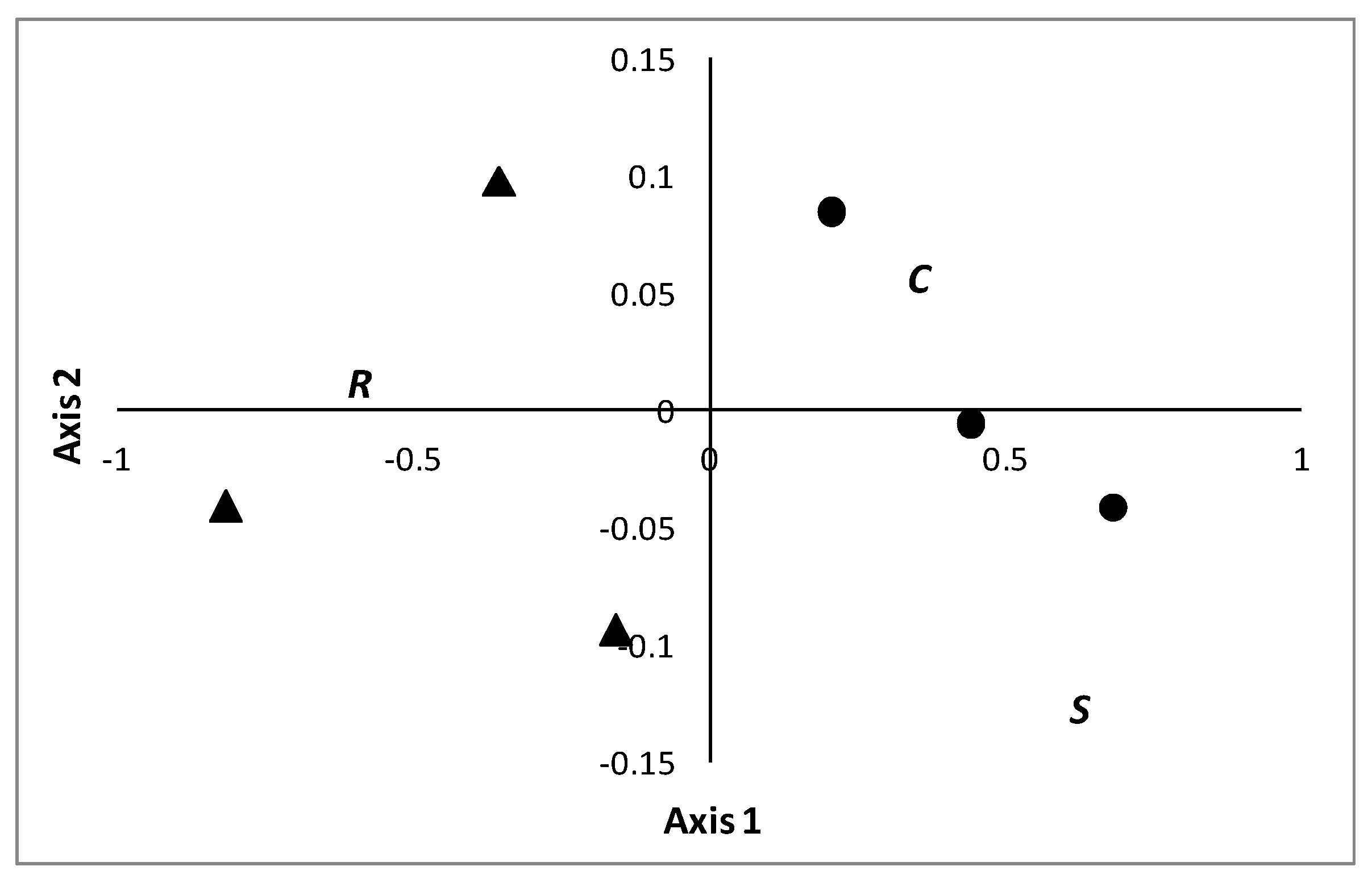
3.1. Comparison between Riparian Forests and Poplar Plantations
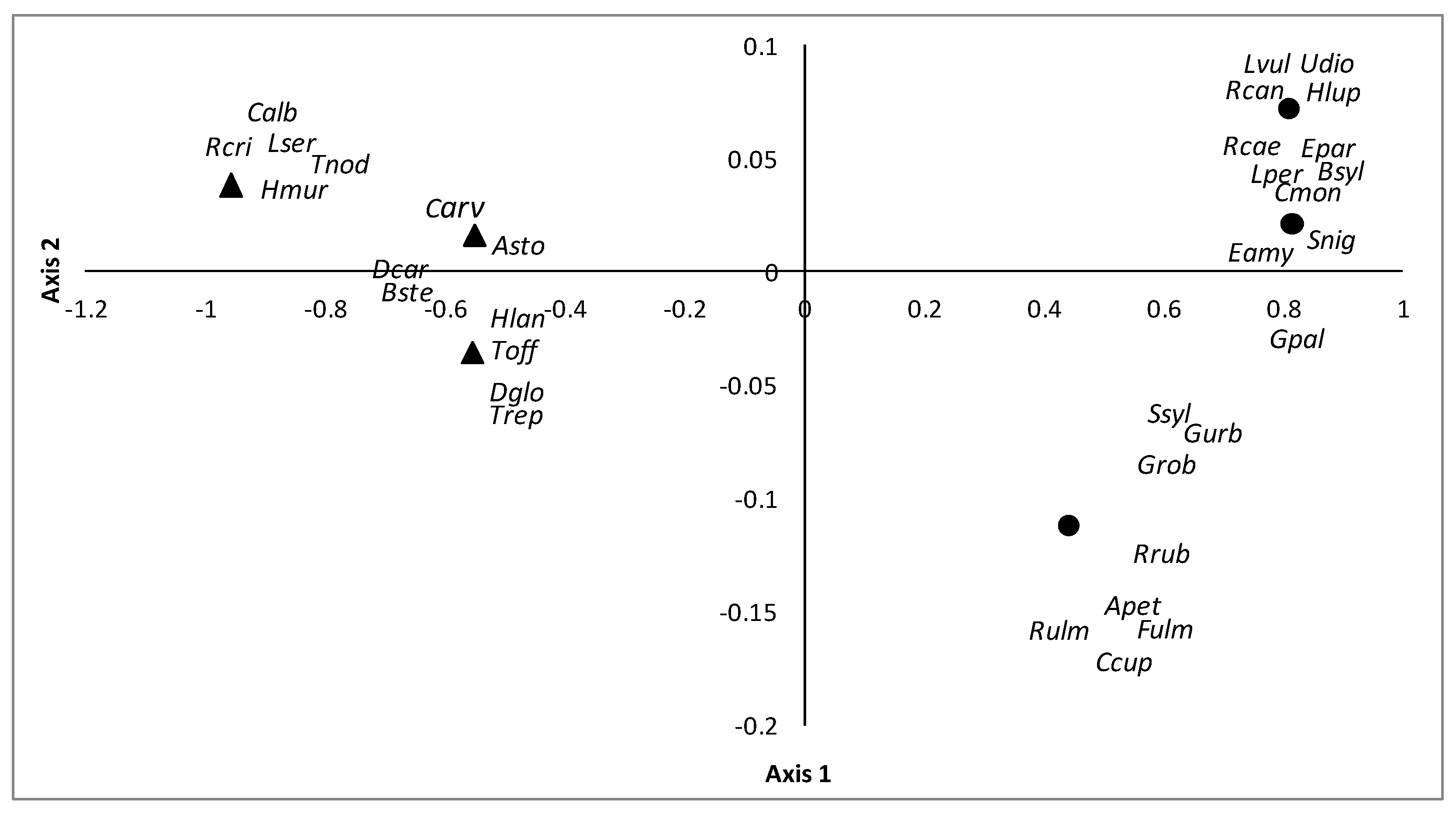
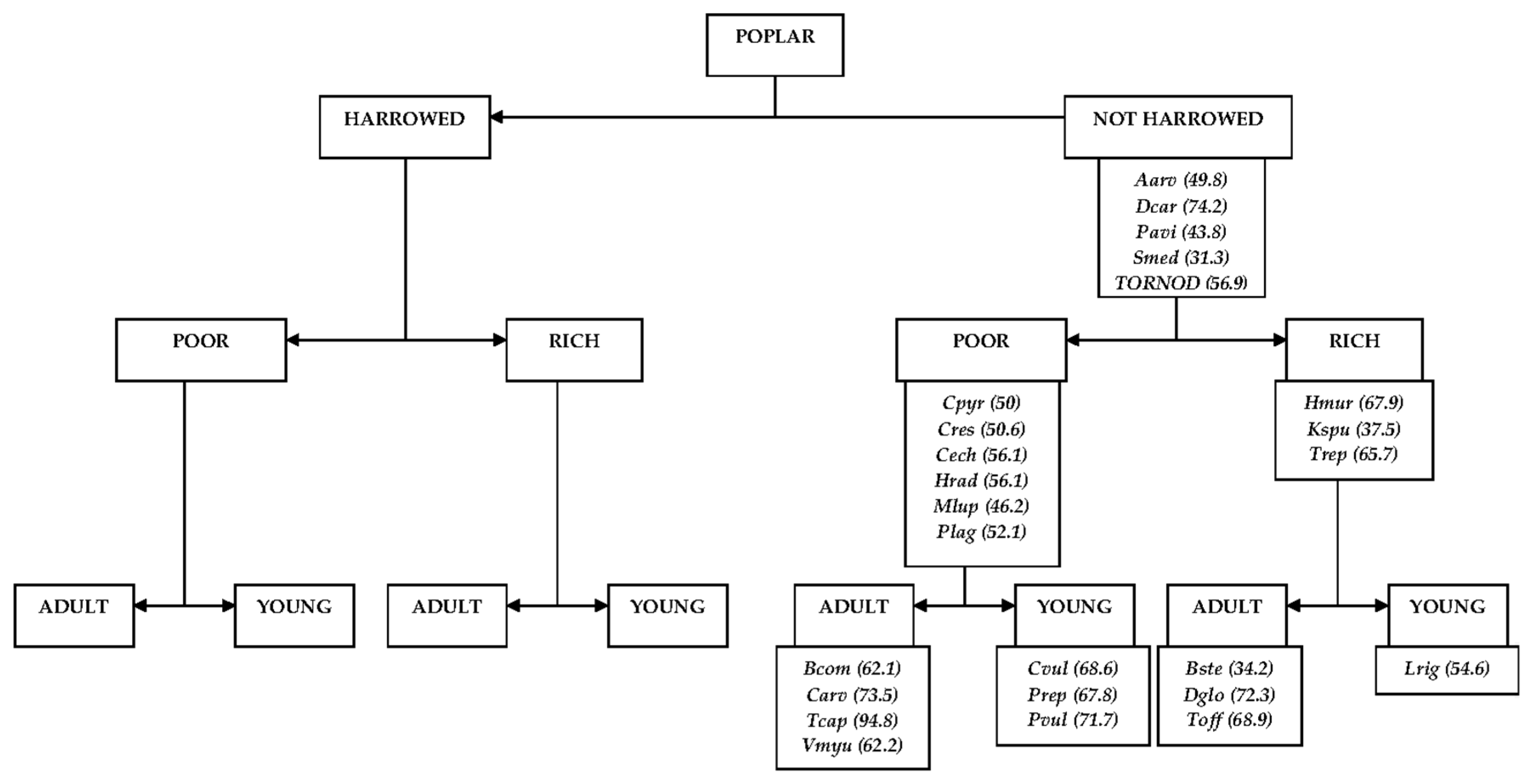
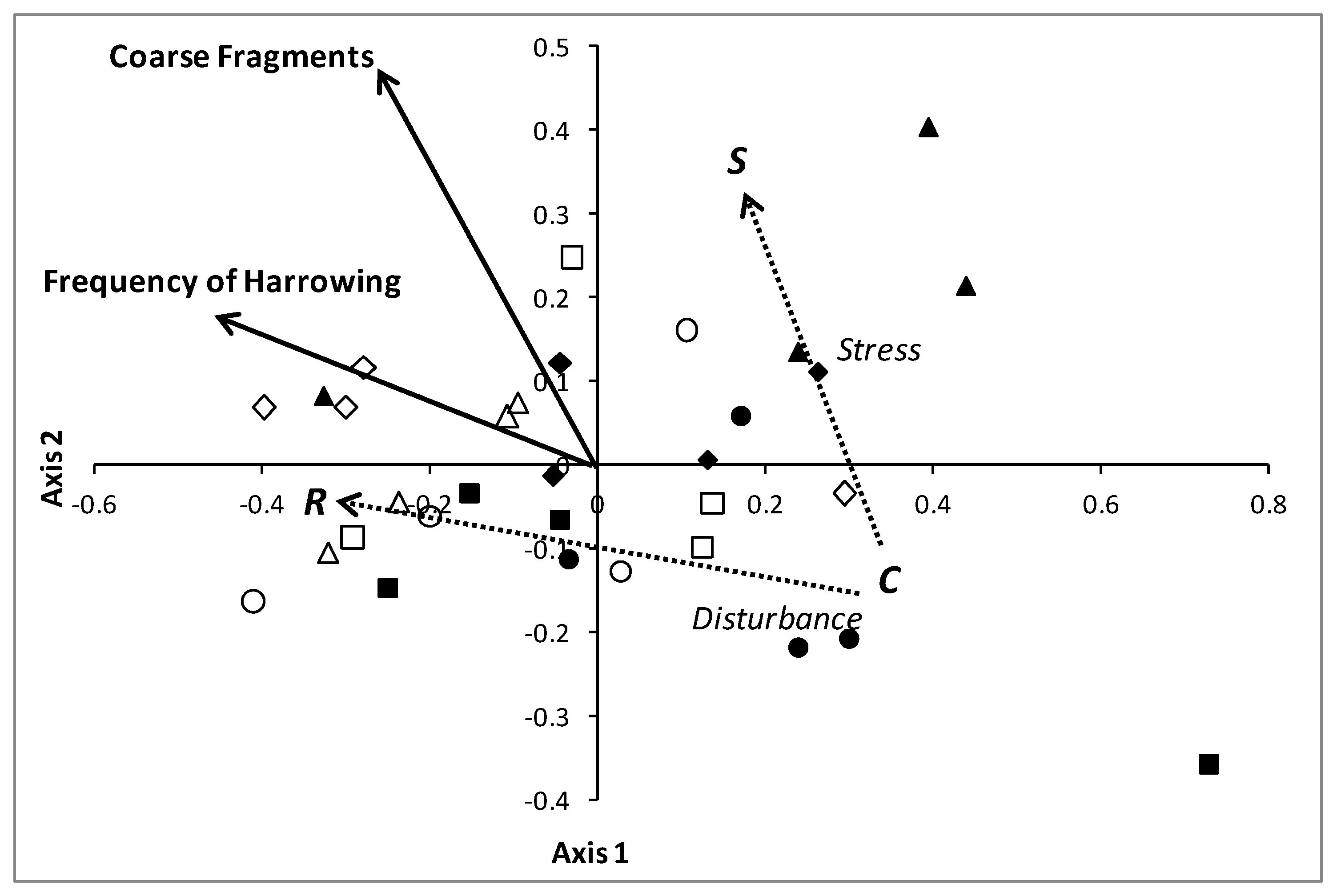
3.2. Comparison of Different Types of Poplar Plantations
4. Discussion
4.1. Comparison between Riparian Forests and Poplar Plantations
4.2. Comparison of Different Types of Poplar Plantations
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones. BioScience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Gundersen, P.; Laurén, A.; Finér, L.; Ring, E.; Koivusalo, H.; Sætersdal, M.; Weslien, J.O.; Sigurdsson, B.B.; Högbom, L.; Laine, J.; et al. Environmental services provided from riparian forests in the Nordic countries. AMBIO 2010, 39, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Naiman, R.J.; Decamps, H.; Pollock, M. The role of riparian corridors in maintaining regional biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Beschta, R.L.; Otting, N.; Lytjen, D. An ecological perspective of riparian and stream restoration in the Western United States. Fisheries 1997, 22, 12–24. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H. The ecology of interfaces: Riparian zones. Ann. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef]
- Stutter, M.I.; Chardon, W.J.; Kronvang, B. Riparian buffer strips as a multifunctional management tool in agricultural landscapes: Introduction. J. Environ. Qual. 2012, 41, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.; Berggren, K. Alterations of Riparian Ecosystems Caused by River Regulation. BioScience 2000, 50, 783–792. [Google Scholar] [CrossRef]
- Schnitzler, A. Conservation of biodiversity in alluvial hardwood forests of the temperate zone. The example of the Rhine valley. For. Ecol. Manag. 1994, 68, 385–398. [Google Scholar] [CrossRef]
- González, M.; García, D. Restauración de Ríos. Guía Metodológica para la Elaboración de Proyectos; Secretaria General Técnica; Centro de Publicaciones, Ministerio de Medio Ambiente: Madrid, Spain, 2007; pp. 1–318. [Google Scholar]
- Bolpagni, R.; Piotti, A. Hydro-hygrophilous vegetation diversity and distribution patterns in riverine wetlands in an agricultural landscape: A case study from the Oglio River (Po Plain, Northern Italy). Phytocoenologia 2015, 45, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Boothroyd-Roberts, K.; Gagnon, D.; Truax, B. Can hybrid poplar plantations accelerate the restoration of forest understory attributes on abandoned fields? For. Ecol. Manag. 2013, 287, 77–89. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A. Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef]
- Brockerhoff, E.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Carnus, J.-M.; Parrotta, J.; Brockerhoff, E.; Arbez, M.; Jactel, H.; Kremer, A.; Lamb, D.; O’Hara, K.; Walters, B. Planted forests and biodiversity. J. For. 2006, 104, 65–77. [Google Scholar]
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95. [Google Scholar] [CrossRef]
- Irwin, S.; Pedley, S.M.; Coote, L.; Dietzsch, A.C.; Wilson, M.W.; Oxbrough, A.; Sweeney, O.; Moore, K.M.; Martin, R.; Kelly, D.L.; et al. The value of plantation forests for plant, invertebrate and bird diversity and the potential for cross-taxon surrogacy. Biodivers. Conserv. 2014, 23, 697–714. [Google Scholar] [CrossRef]
- Loyn, R.; Mcnabb, E.G.; Macak, P.; Noble, P. Eucalypt plantations as habitat for birds on previously cleared farmland in south-eastern Australia. Biodivers. Conserv. 2007, 137, 533–548. [Google Scholar]
- Paquette, A.; Messier, C. The role of plantations in managing the world’s forests in the Anthropocene. Front. Ecol. Environ. 2010, 8, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Archaux, F.; Chevalier, R.; Berthelot, A. Towards practices favourable to plant diversity in hybrid poplar plantations. For. Ecol. Manag. 2010, 259, 2410–2417. [Google Scholar] [CrossRef]
- Laquerbe, M. Communautés de sous-bois des peupleraies artificielles: Relation entre phytomasse, richesse spécifique et perturbations. Ann. For. Sci. 1999, 56, 607–614. [Google Scholar] [CrossRef]
- Laquerbe, M. Richesse spécifique et phytomasse des sous-bois de peupleraies cultivées en bordure de Garonne (Sud-Ouest de la France). Ann. For. Sci. 2000, 57, 767–776. [Google Scholar] [CrossRef]
- Birmele, J.; Gabriele, K.; Frank, B.; Werner, K.; Hans, S.U. Successional changes of phytodiversity on a short rotation coppice plantation in Oberschwaben, Germany. Front. Plant Sci. 2015, 6, 124:1–124:8. [Google Scholar] [CrossRef] [PubMed]
- Soo, T.; Tullus, A.; Tullus, H.; Roosaluste, E. Floristic diversity responses in young hybrid aspen plantations to land-use history and site preparation treatments. For. Ecol. Manag. 2009, 257, 858–867. [Google Scholar] [CrossRef]
- Haeussler, S.; Bedford, L.; Boateng, J.O.; MacKinnon, A. Plant community responses to mechanical site preparation in northern interior British Columbia. Can. J. For. Res. 1999, 29, 1084–1100. [Google Scholar] [CrossRef]
- Gao, T.; Nielsenb, A.B.; Hedblom, M. Reviewing the strength of evidence of biodiversity indicators for forest ecosystems in Europe. Ecol. Indic. 2015, 57, 420–434. [Google Scholar] [CrossRef]
- Gray, A.N.; Azuma, D.L. Repeatability and implementation of a forest vegetation indicator. Ecol. Indic. 2005, 5, 57–71. [Google Scholar] [CrossRef]
- Lindenmayer, D.V.; Margules, C.R.; Botkin, D.A. Indicator of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Aubin, I.; Messier, C.; Bouchard, A. Can plantations develop understory biological and physical attributes of naturally regenerated forests? Biol. Conserv. 2008, 141, 2461–2476. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Fleishman, E.; Noss, R.F.; Noon, B.R. Utility and limitations of species richness metrics for conservation planning. Ecol. Indic. 2006, 6, 543–553. [Google Scholar] [CrossRef]
- Grime, J.P. Competitive exclusion in herbaceous vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties, 2nd ed.; John Wiley & Sons Ltd: Chichester, UK, 2001. [Google Scholar]
- Ninyerola, M.; Pons, X.; Roure, J.M. Atlas Climático Digital de la Península Ibérica. Metodología y Aplicaciones en Bioclimatología y Geobotánica; Universidad Autónoma de Barcelona: Bellaterra, Spain, 2005. [Google Scholar]
- Lara, F.; Garilleti, R.; Calleja, J.A. La Vegetación de Ribera de la Mitad Norte Española; Serie Monografías, 81; Centro de Estudios de Técnicas Aplicadas del CEDEX: Madrid, Spain, 2004; pp. 1–536. [Google Scholar]
- Díaz, L.; Romero, C. Caracterización económica de las choperas en Castilla y León: Rentabilidad y turnos óptimos. In Proceedings of the I Simposio del Chopo, Zamora, Spain, 9–11 May 2001; pp. 489–500.
- Fernández, A.; Hernanz, G. El Chopo (Populus sp.) Manual de Gestión Forestal Sostenible; Junta de Castilla y León: Burgos, Spain, 2004; pp. 1–53. [Google Scholar]
- De Mier, A. Optimización de los sistemas de plantación y producción de chopo. In Proceedings of the I Simposio del Chopo, Zamora, Spain, 9–11 May 2001; pp. 97–105.
- Bravo, F.; Grau, J.M.; Antoñanzas, F.G. Curvas de calidad y tablas de producción para Populus x euramericana en la cuenca del Duero. Montes 1995, 44, 43–46. [Google Scholar]
- Eichhorn, J.; Roskams, P.; Ferretti, M.; Mues, V.; Szepesi, A.; Durrant, D. Part II: Visual assessment of tree condition. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; United Nations Economics Commission for Europe, International Cooperative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP-Forests): Hamburg, Germany, 2010. [Google Scholar]
- Martín-García, J.; Diez, J.J.; Jactel, H. Towards standardised crown condition assessment in poplar plantations. Ann. For. Sci. 2009, 66, 308–314. [Google Scholar] [CrossRef]
- Martín-García, J.; Merino, A.; Diez, J.J. Relating visual crown conditions to nutritional status and site quality in monoclonal poplar plantations (Populus x euramericana). Eur. J. For. Res. 2012, 131, 1185–1198. [Google Scholar] [CrossRef]
- Hunt, R.; Hodgson, J.G.; Thompson, K.; Bungener, P.; Dunnett, N.P.; Askew, A.P. A new practical tool for deriving a functional signature for herbaceous vegetation. Appl. Veg. Sci. 2004, 7, 163–170. [Google Scholar] [CrossRef]
- Wilcox, R.R.; Schönbrodt, F.D. The WRS Package for Robust Statistics in R. R Package Version 0.24, 2014. Available online: https://github.com/nicebread/WRS (accessed on 10 November 2015).
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- R-Development-Core-Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.3–0, 2015. Available online: http://CRAN.R-project.org/package=vegan (accessed on 10 November 2015).
- Stephens, S.S.; Wagner, M.R. Forest plantations and biodiversity: A fresh perspective. J. For. 2007, 105, 307–313. [Google Scholar]
- Fortier, J.; Gagnon, D.; Truax, B.; Lambert, F. Understory plant diversity and biomass in hybrid poplar riparian buffer strips in pastures. New For. 2011, 42, 241–265. [Google Scholar] [CrossRef]
- Roberts, M.R. Effects of forest plantation management on herbaceous-layer composition and diversity. Can. J. Bot. 2002, 80, 378–389. [Google Scholar] [CrossRef]
- Schoonmaker, P.; McKee, A. Species composition and diversity during secondary succession of coniferous forests in the Western Cascade Mountains of Oregon. For. Sci. 1988, 34, 960–979. [Google Scholar]
- Aizpuru, I.; Aseguinolaza, C.; Uribe-Echebarría, P.M.; Urrutia, P.; Zorrakín, I. Claves Ilustradas de la Flora del País Vasco y Territorios Limítrofes; Servicio Central de Publicaciones del Gobierno Vasco: Vitoria-Gasteiz, Spain, 1999; pp. 1–831. [Google Scholar]
- Alejandre, J.A.; García, J.M.; Mateo, J. Atlas de la Flora Vascular Silvestre de Burgos; Monografías de Botánica Ibérica n°2; Jolube Consultor Botánico y Editor: Jaca, Spain, 2006; pp. 1–925. [Google Scholar]
- Li, Y.; Chen, X.; Xie, Y.; Li, X.; Li, F.; Hou, Z. Effects of young poplar plantations on understory plant diversity in the Dongting Lake wetlands, China. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The functional basis of a primary succession resolved by CSR classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Ciccarelli, D. Mediterranean coastal dune vegetation: Are disturbance and stress the key selective forces that drive the psammophilous succession? Estuar. Coast. Shelf. Sci. 2015, 165, 247–253. [Google Scholar] [CrossRef]
- Baum, S.; Weih, M.; Bolte, A. Floristic diversity in Short Rotation Coppice (SRC) plantations: Comparison between soil seed bank and recent vegetation. Appl. Agric. For. Res. 2013, 3, 221–228. [Google Scholar]
- Decocq, G.; Valentin, B.; Toussaint, B.; Hendoux, F.; Saguez, R.; Bardat, J. Soil seed bank composition and diversity in a managed temperate deciduous forest. Biodivers. Conserv. 2004, 13, 2485–2509. [Google Scholar] [CrossRef]
- Baum, S.; Weih, M.; Bolte, A. Stand age characteristics and soil properties affect species composition of vascular plants in short rotation coppice plantations. BioRisk 2012, 7, 51–71. [Google Scholar] [CrossRef]
- Berthelot, A.; Augustin, S.; Godin, J.; Decocq, G. Biodiversity in poplar plantations in the Picardie region of France. Unasylva 2005, 55, 18–19. [Google Scholar]
- Small, C.J.; McCarthy, B.C. Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. For. Ecol. Manag. 2005, 217, 229–243. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Turrill, N.L.; Adams, M.B. Herbaceous-Layer and Overstory Species in Clear-cut and Mature Central Appalachian Hardwood Forests. Ecol. Appl. 1995, 5, 947–955. [Google Scholar] [CrossRef]
- Berthelot, A.; Roguier, S.; Landeau, S. Type d’entretien et diversité floristique sous peupleraie. Rev. For. Fr. 2001, 51, 333–336. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Légaré, S.; Bergeron, Y. Variation of the understory composition and diversity along a gradient of productivity in Populus tremuloides stands in northern of British Columbia, Canada. Can. J. Bot. 2004, 82, 1314–1323. [Google Scholar] [CrossRef]
- Decocq, G.; Aubert, M.; Dupont, F.; Alard, D.; Saguez, R.; Wattez-Franger, A.; De Foucault, B.; Delelis-Dusollier, A.; Bardat, J. Plant diversity in a managed temperate deciduous forest: Understory response to two silvicultural systems. J. Appl. Ecol. 2004, 41, 1065–1079. [Google Scholar] [CrossRef]
- Klinka, K.; Chen, H.Y.H.; Wang, Q.; Montigny, L. Forest canopies and their influence on understory vegetation early-seral stands on West Vancouver Island. Northwest Sci. 1996, 70, 193–200. [Google Scholar]
- Das, D.K.; Chaturvedi, O.P. Structure and function of Populus deltoides agroforestry systems in eastern India: 2. Nutrient dynamics. Agrofor. Syst. 2005, 65, 223–230. [Google Scholar] [CrossRef]
- Browaldh, M. Change in soil mineral nitrogen and respiration following tree harvesting from an agrisilvicultural system in Sweden. Agrofor. Syst. 1997, 35, 131–138. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | ||
|---|---|---|
| Mean (± S.D.) | Range | |
| Coarse fragments (%) | 28.0 (24.2) | 0.64–82.1 |
| Sand (%) | 68.1 (7.0) | 50.9–83.7 |
| Silt (%) | 17.5 (5.5) | 9.6–32.1 |
| Clay (%) | 14.3 (3.1) | 6.7–23.7 |
| pH | 7.5 (0.93) | 5.9–8.6 |
| OM (%) | 2.2 (0.94) | 0.84–5.1 |
| N (%) | 0.11 (0.04) | 0.05–0.22 |
| C/N | 12.1 (4.4) | 6.2–25.9 |
| P (mg·kg−1) | 9.5 (8.8) | 0.1–34.8 |
| K (mg·kg−1) | 105.9 (85.3) | 25.0–374.2 |
| Ca (cmolc·kg−1) | 9.8 (5.4) | 2.8–19.4 |
| Mg (cmolc·kg−1) | 0.61 (0.21) | 0.2–1.13 |
| CEC (cmolc·kg−1) | 13.9 (3.1) | 8.8–22.2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-García, J.; Jactel, H.; Oria-de-Rueda, J.A.; Diez, J.J. The Effects of Poplar Plantations on Vascular Plant Diversity in Riparian Landscapes. Forests 2016, 7, 50. https://doi.org/10.3390/f7030050
Martín-García J, Jactel H, Oria-de-Rueda JA, Diez JJ. The Effects of Poplar Plantations on Vascular Plant Diversity in Riparian Landscapes. Forests. 2016; 7(3):50. https://doi.org/10.3390/f7030050
Chicago/Turabian StyleMartín-García, Jorge, Hervé Jactel, Juan Andrés Oria-de-Rueda, and Julio Javier Diez. 2016. "The Effects of Poplar Plantations on Vascular Plant Diversity in Riparian Landscapes" Forests 7, no. 3: 50. https://doi.org/10.3390/f7030050