Belowground Competition Directs Spatial Patterns of Seedling Growth in Boreal Pine Forests in Fennoscandia

Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Growing season (days) | Temperature sum | Precipitation (mm) |
|---|---|---|---|
| 2000* | 173 | 950 | 483 |
| 2001 | 169 | 995 | 561 |
| 2002 | 150 | 1228 | 216 |
| 2003 | 139 | 1062 | 392 |
| 2004 | 160 | 914 | 394 |
| 2005 | 148 | 992 | 323 |
| 2006 | 168 | 1206 | 307 |
| 2007 | 144 | 922 | 312 |
| 2008 | 170 | 844 | 341 |
| 2009 | 151 | 988 | 382 |
| 2010 | 151 | 1015 | 372 |
| 2011 | 169 | 1225 | 404 |
| 2012 | 146 | 889 | 361 |
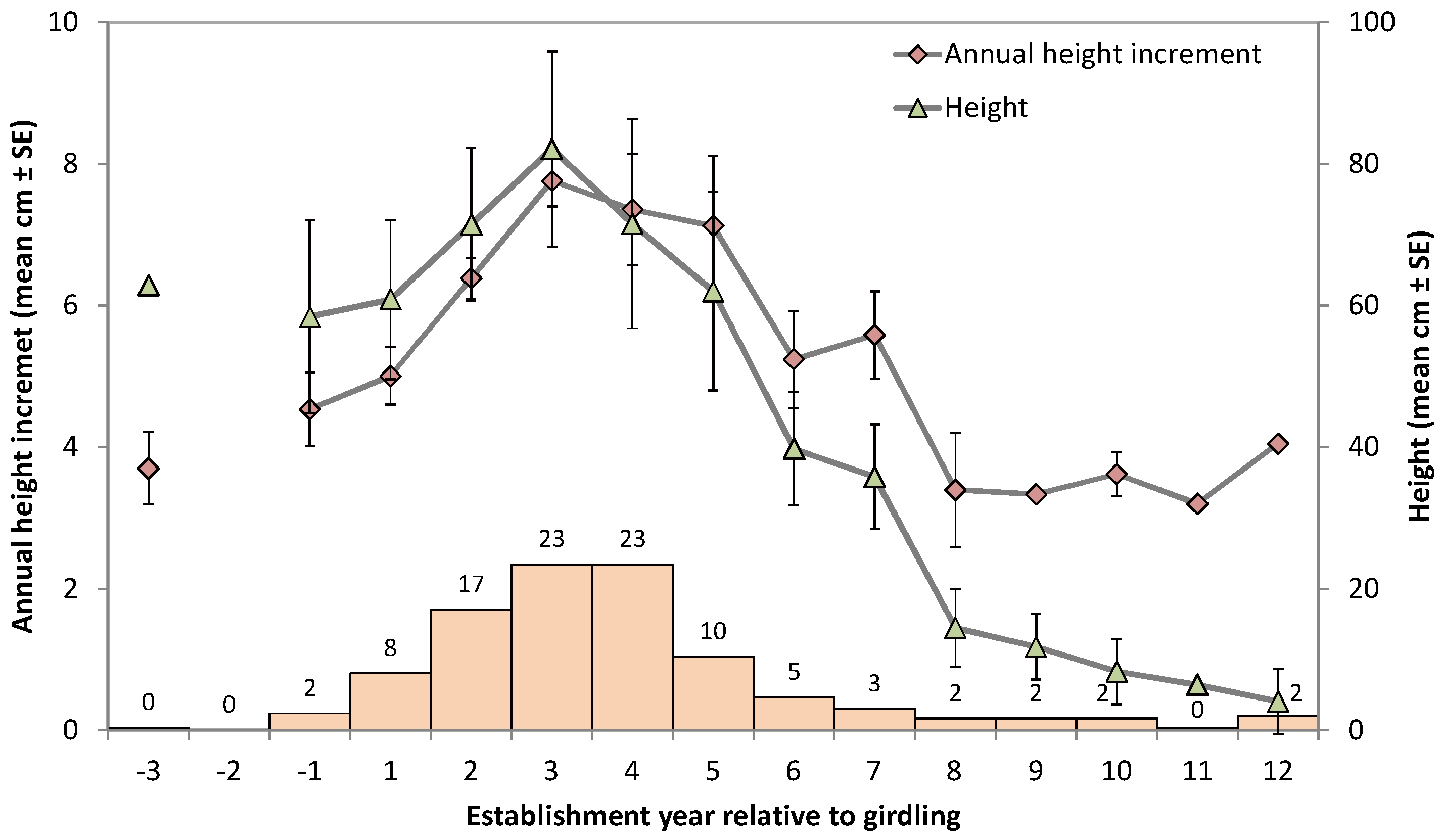
2.2. Pine Seedling Establishment and Growth in a Tree Girdling Experiment
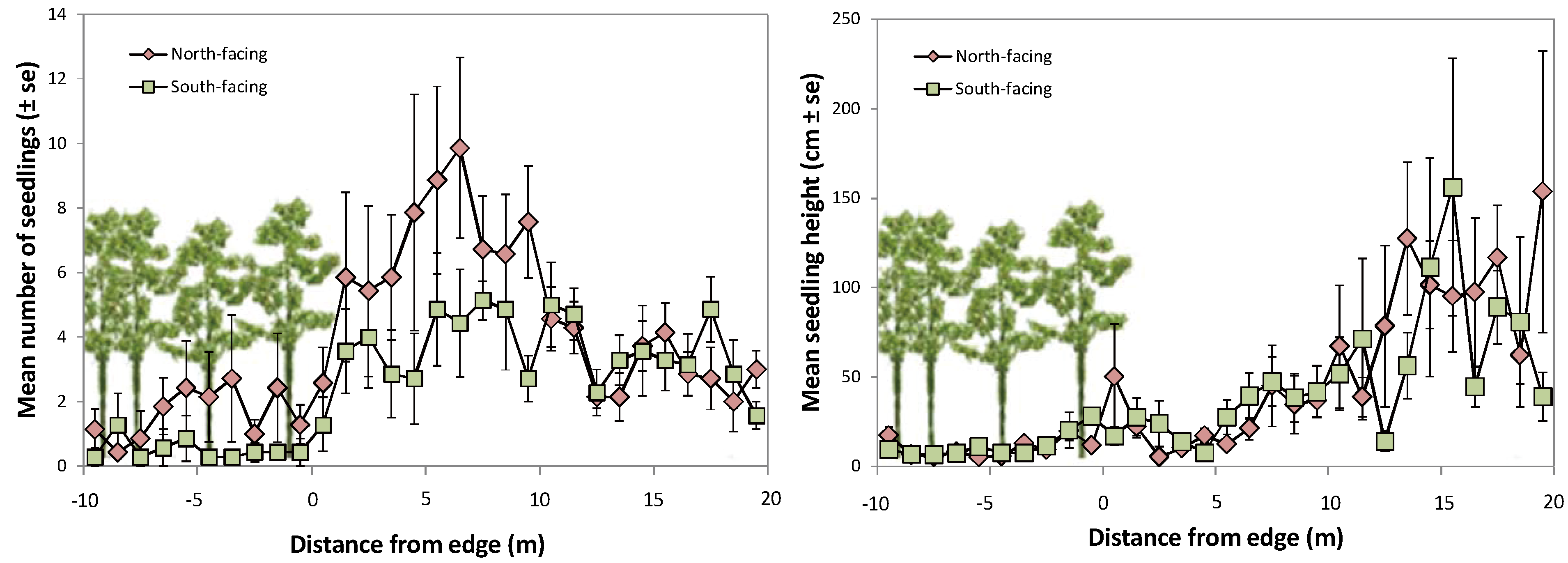
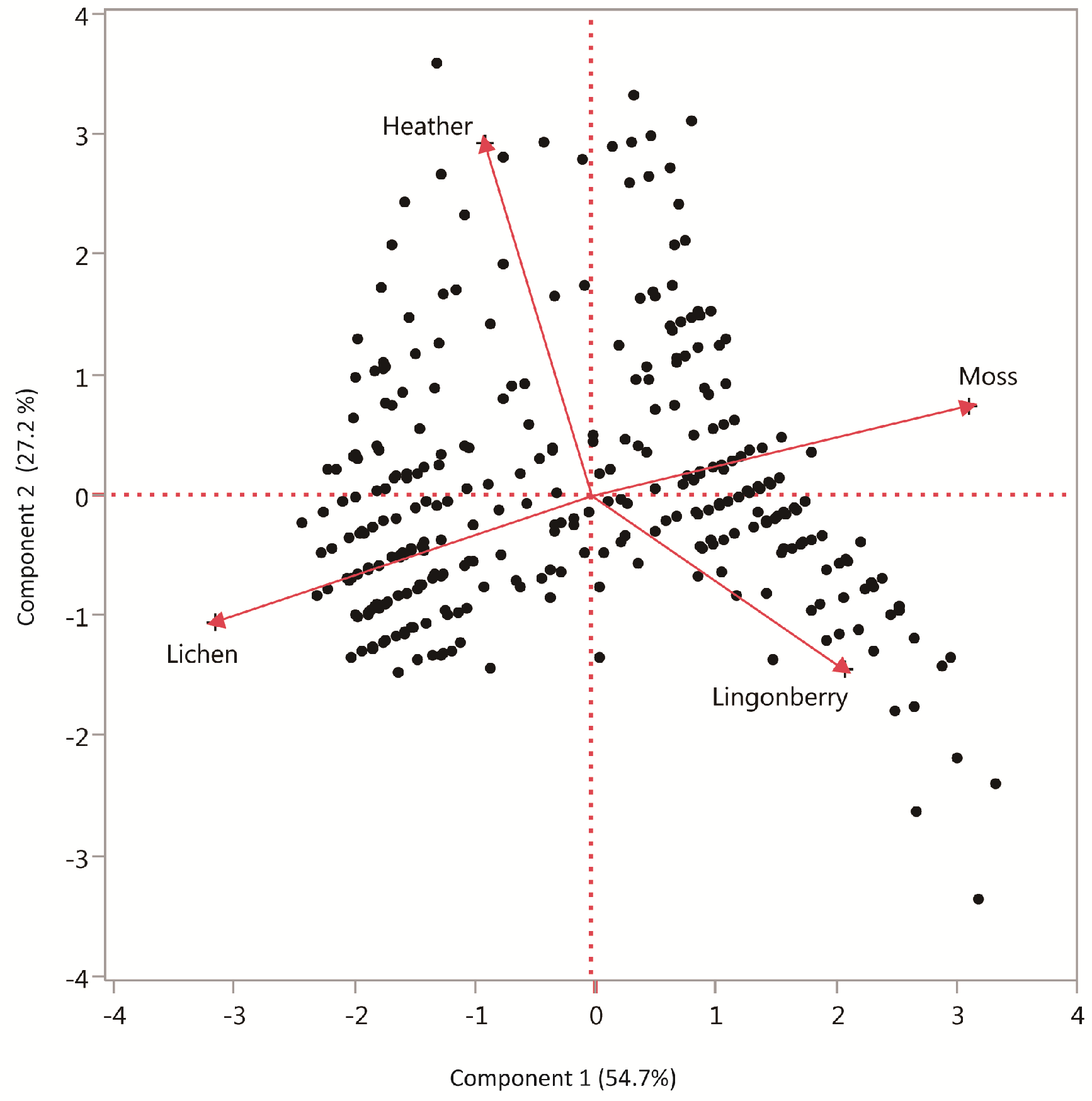
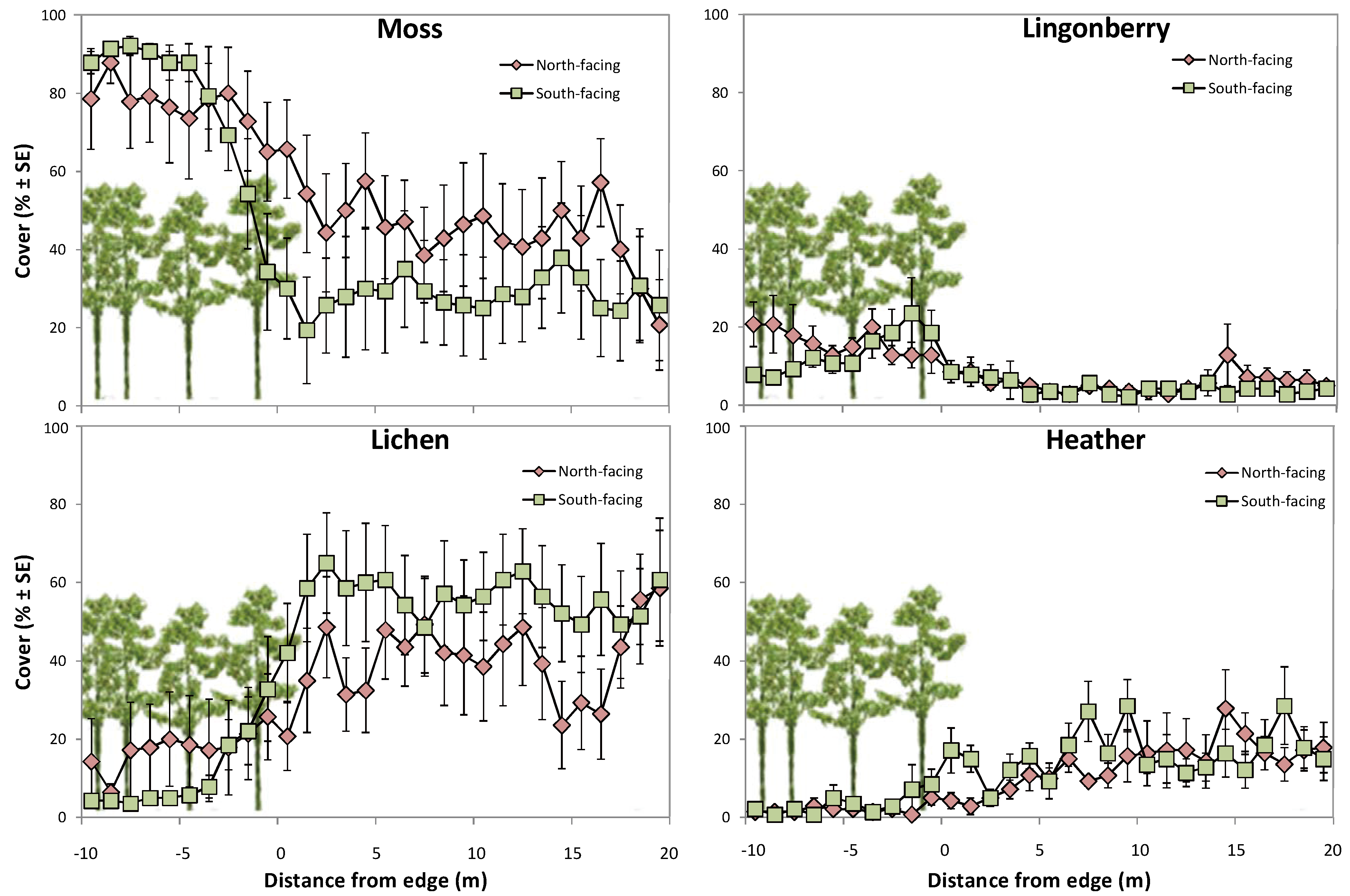
2.3. Pine Seedlings and Ground Vegetation in Forest Gaps in Relation to Edges
2.4. Statistical Analyses
3. Results
3.1. Effect of Release from Belowground Competition through Tree Girdling

3.2. Pine Seedling Establishment in Forest Gaps in Relation to Edges

| Source | Degrees of freedom | F-value | p-value |
|---|---|---|---|
| Number of pine | |||
| Facing | 1 | 6.3974 | 0.0205 |
| Distance | 1 | 7.8733 | 0.0062 |
| Facing × Distance | 1 | 0.2602 | 0.6119 |
| Principal Component 1 (−) | 1 | 16.6519 | <0.0001 |
| Principal Component 2 (+) | 1 | 0.0110 | 0.9165 |
| Mean height | |||
| Facing | 1 | 0.8088 | 0.3768 |
| Distance | 1 | 68.1455 | <0.0001 |
| Facing × Distance | 1 | 0.5661 | 0.4538 |
| Principal Component 1 (+) | 1 | 13.0360 | 0.0005 |
| Principal Component 2 (+) | 1 | 7.2251 | 0.0077 |

| Source | Degrees of freedom | F-value | p-value |
|---|---|---|---|
| Moss | |||
| Facing | 1 | 0.3240 | 0.5772 |
| Distance | 1 | 25.3807 | <0.0001 |
| Facing × Distance | 1 | 0.0116 | 0.9151 |
| Lichen | |||
| Facing | 1 | 0.0402 | 0.844 |
| Distance | 1 | 27.4193 | <0.0001 |
| Facing × Distance | 1 | 0.8469 | 0.3643 |
| Lingonberry | |||
| Facing | 1 | 2.2270 | 0.1518 |
| Distance | 1 | 29.0796 | <0.0001 |
| Facing × Distance | 1 | 0.2140 | 0.6454 |
| Heather | |||
| Facing | 1 | 1.2409 | 0.2828 |
| Distance | 1 | 78.2066 | <0.0001 |
| Facing × Distance | 1 | 0.3274 | 0.5687 |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Caplat, P.; Anand, M.; Bauch, C. Symmetric competition causes population oscillations in an individual-based model of forest dynamics. Ecol. Model. 2008, 211, 491–500. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y.H. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient. J. Veg. Sci. 2013, 24, 543–552. [Google Scholar] [CrossRef]
- Thorpe, A.S.; Aschehoug, E.T.; Atwater, D.Z.; Callaway, R.M. Interactions among plants and evolution. J. Ecol. 2011, 99, 729–740. [Google Scholar]
- McCarthy, J. Gap dynamics of forest trees: A review with particular attention to boreal forests. Environ. Rev. 2001, 9, 1–59. [Google Scholar] [CrossRef]
- Petritan, I.C.; von Lupke, B.; Petritan, A.M. Effects of root trenching of overstorey norway spruce (Picea abies) on growth and biomass of underplanted beech (Fagus sylvatica) and douglas fir (Pseudotsuga menziesii) saplings. Eur. J. For. Res. 2011, 130, 813–828. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Hokkanen, T.J.; Jarvinen, E.; Pukkala, T. Factors related to seedling growth in a boreal scots pine stand: A spatial-analysis of a vegetation-soil system. Can. J. For. Res. 1993, 23, 2101–2109. [Google Scholar] [CrossRef]
- Wagner, S.; Fischer, H.; Huth, F. Canopy effects on vegetation caused by harvesting and regeneration treatments. Eur. J. For. Res. 2011, 130, 17–40. [Google Scholar] [CrossRef]
- Chapin, F.S.; Bloom, A.J.; Field, C.B.; Waring, R.H. Plant-responces to multiple environmental-factors. Bioscience 1987, 37, 49–57. [Google Scholar]
- Schwinning, S.; Weiner, J. Mechanisms determining the degree of size asymmetry in competition among plants. Oecologia 1998, 113, 447–455. [Google Scholar] [CrossRef]
- Coomes, D.A.; Grubb, P.J. Impacts of root competition in forests and woodlands: A theoretical framework and review of experiments. Ecol. Monogr. 2000, 70, 171–207. [Google Scholar] [CrossRef]
- Aaltonen, V.T. Über die natürliche verjungung der heidevälder im Finnischen lappland I. Commun. Inst. Forest. Finlandiae 1919, 1, 1–319, (In Finnish with German summary). [Google Scholar]
- Björkman, E. On the influence of light on the height-growth of pine plants on pine heaths in Norrland. Medd. Stat. Skogsfors. Anst. 1945, 34, 497–542. [Google Scholar]
- Romell, L.G.; Malmström, C. Henrik Hesselmans tallhedsförsök åren 1922–1942 the Ecology of Lichen-Pine Forest: Experiments (1922–1942) by the late dr Henrik Hesselman; Statens skogsförsöksanstalt: Stockholm, Sweden, 1945. [Google Scholar]
- Strand, M.; Löfvenius, M.O.; Bergsten, U.; Lundmark, T.; Rosvall, O. Height growth of planted conifer seedlings in relation to solar radiation and position in scots pine shelterwood. For. Ecol. Manage. 2006, 224, 258–265. [Google Scholar] [CrossRef]
- de la Rosa, T.M.; Aphalo, P.J.; Lehto, T. Effects of far-red light on the growth, mycorrhizas and mineral nutrition of scots pine seedlings. Plant Soil 1998, 201, 17–25. [Google Scholar] [CrossRef]
- Pukkala, T.; Kuuluvainen, T.; Stenberg, P. Below-canopy distribution of photosynthetically active radiation and its relation to seedling growth in a boreal Pinus sylvestris stand—A simulation approach. Scand. J. For. Res. 1993, 8, 313–325. [Google Scholar]
- De Chantal, M.; Leinonen, K.; Kuuluvainen, T.; Cescatti, A. Early response of Pinus sylvestris and Picea abies seedlings to an experimental canopy gap in a boreal spruce forest. For. Ecol. Manage. 2003, 176, 321–336. [Google Scholar] [CrossRef]
- Björkman, E.; Lundeberg, G. Studies on root competition in poor pine forests by supply of labeled nitrogen and phosphorus. Stud. For. Suec. 1971, 94, 1–16. [Google Scholar]
- Hesselman, H. Studier över de norrländska tallhedarnas föryngringsvilllkor i. (studien uber die verjungungsbedingungen der norrlandischen kiefernheiden i.). Medd. Stat. Skogsfors. Anst. 1910, 7, 25–68. [Google Scholar]
- Kuuluvainen, T.; Yllasjarvi, I. On the natural regeneration of dry heath forests in finnish lapland: A review of V. T. Aaltonen (1919). Scand. J. For. Res. 2011, 26, 34–44. [Google Scholar] [CrossRef]
- Rouvinen, S.; Kouki, J. Tree regeneration in artificial canopy gaps established for restoring natural structural variability in a scots pine stand. Silva Fennica 2011, 45, 1079–1091. [Google Scholar]
- Valkonen, S.; Ruuska, J.; Siipilehto, J. Effect of retained trees on the development of young scots pine stands in Southern Finland. For. Ecol. Manage. 2002, 166, 227–243. [Google Scholar] [CrossRef]
- Hancock, M.; Egan, S.; Summers, R.; Cowie, N.; Amphlett, A.; Rao, S.; Hamilton, A. The effect of experimental prescribed fire on the establishment of scots pine Pinus sylvestris seedlings on heather Calluna vulgaris moorland. For. Ecol. Manage. 2005, 212, 199–213. [Google Scholar] [CrossRef]
- Scott, D.; Welch, D.; Thurlow, M.; Elston, D.A. Regeneration of Pinus sylvestris in a natural pinewood in ne scotland following reduction in grazing by Cervus elaphus. For. Ecol. Manage. 2000, 130, 199–211. [Google Scholar] [CrossRef]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef]
- Bhupinderpal-Singh; Nordgren, A.; Löfvenius, M.O.; Högberg, M.N.; Mellander, P.E.; Högberg, P. Tree root and soil heterotrophic respiration as revealed by girdling of boreal scots pine forest: Extending observations beyond the first year. Plant Cell Environ. 2003, 26, 1287–1296. [Google Scholar] [CrossRef]
- Högberg, M.N.; Högberg, P. Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytol. 2002, 154, 791–795. [Google Scholar]
- Hesselman, H. Studier över de norrländska tallhedarnas föryngringsvilllkor ii. (studien uber die verjungungsbedingungen der norrlandischen kiefernheiden ii.). Medd. Stat. Skogsfors. Anst. 1917, 13–14, 1221–1286. [Google Scholar]
- Högberg, M.N.; Högberg, P.; Myrold, D.D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 2007, 150, 590–601. [Google Scholar]
- Yarwood, S.A.; Myrold, D.D.; Högberg, M.N. Termination of belowground C allocation by trees alters soil fungal and bacterial communities in a boreal forest. FEMS Microbiol. Ecol. 2009, 70, 151–162. [Google Scholar] [CrossRef]
- Gaudio, N.; Balandier, P.; Perret, S.; Ginisty, C. Growth of understorey scots pine (Pinus sylvestris L.) saplings in response to light in mixed temperate forest. Forestry 2011, 84, 187–195. [Google Scholar] [CrossRef]
- Brumelis, G.; Elferts, D.; Liepina, L.; Luce, I.; Tabors, G.; Tjarve, D. Age and spatial structure of natural Pinus sylvestris stands in latvia. Scand. J. For. Res. 2005, 20, 471–480. [Google Scholar] [CrossRef]
- Chojnacki, Y. About the method to determine accurately the age of saplings and the annual height growth of some forest tree species. Sylwan 1964, 1, 71–76. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- SAS. Discovering JMP; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
- Melin, J.; Nommik, H.; Lohm, U.; Flower-Ellis, J. Fertilizer nitrogen budgets in a scots pine ecosytem attained by using root-isolated plots an N-15 tracer technique. Plant Soil 1983, 74, 249–263. [Google Scholar] [CrossRef]
- Göttlicher, S.G.; Taylor, A.F.S.; Grip, H.; Betson, N.R.; Valinger, E.; Högberg, M.N.; Högberg, P. The lateral spread of tree root systems in boreal forests: Estimates based on N-15 uptake and distribution of sporocarps of ectomycorrhizal fungi. For. Ecol. Manage. 2008, 255, 75–81. [Google Scholar] [CrossRef]
- Erefur, C.; Bergsten, U.; de Chantal, M. Establishment of direct seeded seedlings of norway spruce and scots pine: Effects of stand conditions, orientation and distance with respect to shelter tree, and fertilisation. For. Ecol. Manage. 2008, 255, 1186–1195. [Google Scholar] [CrossRef]
- Steijlen, I.; Nilsson, M.C.; Zackrisson, O. Seed regeneration of scots pine in boreal forest stands dominated by lichens and feather moss. Can. J. For. Res. 1995, 25, 713–723. [Google Scholar] [CrossRef]
- Jarvis, P.; Linder, S. Botany: Constraints to growth of boreal forests. Nature 2000, 405, 904–905. [Google Scholar] [CrossRef]
- Tamm, C.O. Nitrogen in Terrestrial Ecosystems. Questions of Productivity, Vegetational Changes, and Ecosystem Stability; Springer Verlag: Berlin, Germany, 1991. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Axelsson, E.P.; Lundmark, T.; Högberg, P.; Nordin, A. Belowground Competition Directs Spatial Patterns of Seedling Growth in Boreal Pine Forests in Fennoscandia. Forests 2014, 5, 2106-2121. https://doi.org/10.3390/f5092106
Axelsson EP, Lundmark T, Högberg P, Nordin A. Belowground Competition Directs Spatial Patterns of Seedling Growth in Boreal Pine Forests in Fennoscandia. Forests. 2014; 5(9):2106-2121. https://doi.org/10.3390/f5092106
Chicago/Turabian StyleAxelsson, E. Petter, Tomas Lundmark, Peter Högberg, and Annika Nordin. 2014. "Belowground Competition Directs Spatial Patterns of Seedling Growth in Boreal Pine Forests in Fennoscandia" Forests 5, no. 9: 2106-2121. https://doi.org/10.3390/f5092106