
Seasonal and Depth Dynamics of Soil Moisture Affect Trees on the Tibetan Plateau
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Tree-Ring Isotope Data
2.3. Meteorological Data
2.4. The SM Data
2.5. Data Analysis
3. Results
3.1. The Spatial and Temporal Patterns of the δ18OTR
3.2. The SM Dynamics at Different Depth Gradients
3.3. The Response of δ18OTR to Climatic and SM Factors
3.4. The Correlation between δ18OTR and Key Factors during the Growing Season
3.5. The Spatial Patterns of δ18OTR Response to SM with Soil Depths
4. Discussion
4.1. The δ18OTR Records Soil Water Use by Tree Growth on the TP
4.2. Seasonal Use Strategies of SM for Trees across Wet and Dry Gradients
4.3. Depth Use Strategies of SM for Trees across Wet and Dry Gradients
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Keyimu, M.; Li, Z.; Liu, G.; Fu, B.; Fan, Z.; Wang, X.; Halik, U. Tree-ring based minimum temperature reconstruction on the southeastern Tibetan Plateau. Quat. Sci. Rev. 2021, 251, 106712. [Google Scholar] [CrossRef]
- Wang, J.; Taylor, A.R.; D’Orangeville, L. Warming-induced tree growth may help offset increasing disturbance across the Canadian boreal forest. Proc. Natl. Acad. Sci. USA 2023, 120, e2212780120. [Google Scholar] [CrossRef] [PubMed]
- Seddon, A.W.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of global terrestrial ecosystems to climate variability. Nature 2023, 531, 229–232. [Google Scholar] [CrossRef]
- Priyadarshini, K.V.R.; Prins, H.H.; Bie, S.; Heitkönig, I.M.; Woodborne, S.; Gort, G.; Kroon, H. Seasonality of hydraulic redistribution by trees to grasses and changes in their water-source use that change tree-grass interactions. Ecohydrology 2016, 9, 218–228. [Google Scholar] [CrossRef]
- Amer, A.; Franceschi, E.; Hjazin, A. Structure and Ecosystem Services of Three Common Urban Tree Species in an Arid Climate City. Forests 2023, 14, 671. [Google Scholar] [CrossRef]
- Eslaminejad, P.; Heydari, M.; Kakhki, F.V.; Mirab-balou, M.; Omidipour, R.; Muñoz-Rojas, M.; Lucas-Borja, M.E. Plant species and season influence soil physicochemical properties and microbial function in a semi-arid woodland ecosystem. Plant Soil 2020, 456, 43–59. [Google Scholar] [CrossRef]
- Ma, J.; Han, J.; Zhang, Y.; Dong, Q.; Lei, N.; Liu, Z.; Du, Y. Temporal stability of soil water content on slope during the rainy season in gully regulation watershed. Environ. Earth Sci. 2020, 79, 173. [Google Scholar] [CrossRef]
- Bertrand, G.; Masini, J.; Goldscheider, N.; Meeks, J.; Lavastre, V.; Celle-Jeanton, H.; Hunkeler, D. Determination of spatiotemporal variability of tree water uptake using stable isotopes (δ18O, δ2H) in an alluvial system supplied by a high-altitude watershed, Pfyn forest, Switzerland. Ecohydrology 2014, 7, 319–333. [Google Scholar] [CrossRef]
- Wang, H.; Tetzlaff, D.; Soulsby, C. Modelling the effects of land cover and climate change on soil water partitioning in a boreal headwater catchment. J. Hydrol. 2018, 558, 520–531. [Google Scholar] [CrossRef]
- Zeng, Q.; Rossi, S.; Yang, B.; Qin, C.; Li, G. Environmental drivers for cambial reactivation of Qilian junipers (Juniperus przewalskii) in a semi-arid region of northwestern China. Atmosphere 2020, 11, 232. [Google Scholar] [CrossRef]
- Zhang, F.; Jia, W.; Zhu, G.; Shi, Y.; Zhang, M. Using stable isotopes to investigate differences of plant water sources in subalpine habitats. Hydrol. Process. 2022, 36, e14518. [Google Scholar] [CrossRef]
- Gao, S.; Liu, R.; Zhou, T.; Fang, W.; Yi, C.; Lu, R.; Luo, H. Dynamic responses of tree-ring growth to multiple dimensions of drought. Glob. Change Biol. 2018, 24, 5380–5390. [Google Scholar] [CrossRef] [PubMed]
- Bose, T.; Sengupta, S.; Chakraborty, S.; Borgaonkar, H. Reconstruction of soil water oxygen isotope values from tree ring cellulose and its implications for paleoclimate studies. Quat. Int. 2016, 425, 387–398. [Google Scholar] [CrossRef]
- Xu, Q.; Li, H.; Chen, J.; Cheng, X.; Liu, S.; An, S. Water use patterns of three species in subalpine forest, Southwest China: The deuterium isotope approach. Ecohydrology 2011, 4, 236–244. [Google Scholar] [CrossRef]
- Roden, J.S.; Lin, G.; Ehleringe, J.R. A mechanistic model for interpretation of hydrogen and oxygen isotope ratios in tree-ring cellulose. Geochim. Cosmochim. Acta 2000, 64, 21–35. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M. Least-cost input mixtures of water and nitrogen for photosynthesis. Am. Nat. 2003, 161, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Deng, W.; Wei, G.; Chen, J.; Zheng, X.; Shi, T.; Zeng, T. A pilot study on zinc isotopic compositions in shallow-water coral skeletons. Geochem. Geophys. Geosyst. 2020, 21, e2020GC009430. [Google Scholar] [CrossRef]
- Deurwaerder, H.; Hervé-Fernández, P.; Stahl, C.; Burban, B.; Petronelli, P.; Hoffman, B.; Verbeeck, H. Liana and tree below-ground water competition-evidence for water resource partitioning during the dry season. Tree Physiol. 2018, 38, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Gessler, A.; Ferrio, J.P.; Hommel, R.; Treydte, K.; Werner, R.A.; Monson, R.K. Stable isotopes in tree rings: Towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 2014, 34, 796–818. [Google Scholar] [CrossRef]
- Hahm, W.J.; Dralle, D.N.; Rempe, D.M.; Bryk, A.B.; Thompson, S.E.; Dawson, T.E.; Dietrich, W.E. Low subsurface water storage capacity relative to annual rainfall decouples Mediterranean plant productivity and water use from rainfall variability. Geophys. Res. Lett. 2019, 46, 6544–6553. [Google Scholar] [CrossRef]
- Lind, M.; Zhang, L.; Falste, D.; Franklin, O.; Brännström, A. Plant diversity and drought: The role of deep roots. Ecol. Modell. 2014, 290, 85–93. [Google Scholar] [CrossRef]
- Baker, J.C.A.; Gloor, M.; Spracklen, D.V.; Arnold, S.R.; Tindall, J.C.; Clerici, S.J.; Brienen, R.J.W. What drives interannual variation in tree ring oxygen isotopes in the Amazon. Geophys. Res. Lett. 2016, 43, 11–831. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.; Leavit, S.; Cresse, E.L.; Quine, T.A.; Shi, J.; Shi, S. Tree-ring δ18O identifies similarity in timing but differences in depth of soil water uptake by trees in mesic and arid climates. Agric. For. Meteorol. 2021, 308, 108569. [Google Scholar] [CrossRef]
- Yang, B.; Wen, X.; Sun, X. Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region. Agric. For. Meteorol. 2015, 201, 218–228. [Google Scholar] [CrossRef]
- Sun, J.; Ye, C.; Liu, M.; Wang, Y.; Chen, J.; Wang, S.; Tsubo, M. Response of net reduction rate in vegetation carbon uptake to climate change across a unique gradient zone on the Tibetan Plateau. Environ. Res. 2022, 203, 111894. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Fong, A.; McVicar, N.; Smith, S.; Giambattista, J.; Wells, D.; Alexander, A. Comparing deep learning-based auto-segmentation of organs at risk and clinical target volumes to expert inter-observer variability in radiotherapy planning. Radiother. Oncol. 2020, 144, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Fang, O.; Zhang, Q.B. Tree resilience to drought increases in the Tibetan Plateau. Glob. Change Biol. 2019, 25, 245–253. [Google Scholar] [CrossRef]
- Ge, F.; Sielmann, F.; Zhu, X.; Fraedrich, K.; Zhi, X.; Peng, T.; Wang, L. The link between Tibetan Plateau monsoon and Indian summer precipitation: A linear diagnostic perspective. Clim. Dyn. 2017, 49, 4201–4215. [Google Scholar] [CrossRef]
- Tan, L.; Cai, Y.; Edwards, L.R.; Lan, J.; Zhang, H.; Gao, Y. High resolution monsoon precipitation changes on southeastern Tibetan Plateau over the past 2300 years. Quat. Sci. Rev. 2018, 195, 122–132. [Google Scholar] [CrossRef]
- Grießinger, J.; Bräuning, A.; Helle, G.; Schleser, G.H.; Hochreuther, P.; Meier, W.J.H.; Zhu, H. A dual stable isotope approach unravels common climate signals and species-specific responses to environmental change stored in multi-century tree-ring series from the Tibetan plateau. Geosciences 2019, 9, 151. [Google Scholar] [CrossRef]
- Xu, C.; Zhu, H.; Nakatsuka, T.; Sano, M.; Li, Z.; Shi, F.; Guo, Z. Sampling strategy and climatic implication of tree-ring cellulose oxygen isotopes of Hippophae tibetana and Abies georgei on the southeastern Tibetan Plateau. Int. J. Biometeorol. 2018, 63, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Sano, M.; Dimri, A.P.; Ramesh, R. Moisture source signals preserved in a 242-year tree-ring δ18O chronology in the western Himalaya. Glob. Planet. Change 2017, 157, 73–82. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agric. For. Meteorol. 2018, 260, 165–175. [Google Scholar] [CrossRef]
- Shi, P.J.; Huang, J.G.; Hui, C.; Grissino-Mayer, H.D.; Tardif, J.; Zhai, L.H.; Li, B.L. Capturing spiral radial growth of conifers using the superellipse to model tree-ring geometric shape. Front. Plant Sci. 2015, 6, 856. [Google Scholar] [CrossRef] [PubMed]
- Wernicke, J.; Grießinger, J.; Hochreuther, P.; Bräuning, A. Variability of summer humidity during the past 800 years on the eastern Tibetan Plateau inferred from δ18O of tree-ring cellulose. Clim. Past. 2014, 10, 3327–3356. [Google Scholar]
- Sano, M.; Tshering, P.; Komori, J. May-September precipitation in the Bhutan Himalaya since 1743 as reconstructed from tree ring cellulose δ18O. J. Geophys. Res. Atmos. 2013, 118, 8399–8410. [Google Scholar] [CrossRef]
- Treydte, K.S.; Schleser, G.H.; Helle, G. The twentieth century was the wettest period in northern Pakistan over the past millennium. Nature 2006, 440, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Qin, C.; Bräuning, A. Long-term decrease in Asian monsoon rainfall and abrupt climate change events over the past 6,700 years. Proc. Natl. Acad. Sci. USA 2021, 118, e2102007118. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Li, X.; Evans, M.N.; Wang, W.; An, W.; Xu, G.; Wu, G. Seasonal incursion of Indian Monsoon humidity and precipitation into the southeastern Qinghai-Tibetan Plateau inferred from tree ring δ18O values with intra-seasonal resolution. Earth Planet. Sci. Lett. 2016, 443, 9–19. [Google Scholar] [CrossRef]
- Murray, F.W. On the Computation of Saturation Vapor Pressure; Rand Corporation: Santa Monica, CA, USA, 1966. [Google Scholar]
- New, M.; Hulme, M.; Jones, P. Representing twentieth-century space-time climate variability. Part I: Development of a 1961-90 mean monthly terrestrial climatology. J. Clim. 1999, 12, 829–856. [Google Scholar] [CrossRef]
- Duan, A.; Xiao, Z. Does the climate warming hiatus exist over the Tibetan Plateau? Sci. Rep. 2015, 5, 13711. [Google Scholar] [CrossRef]
- Jiao, L.; Jiang, Y.; Zhang, W.; Wang, M.; Wang, S.; Liu, X. Assessing the stability of radial growth responses to climate change by two dominant conifer trees species in the Tianshan Mountains, northwest China. For. Ecol. Manag. 2019, 433, 667–677. [Google Scholar] [CrossRef]
- Li, Y.; Peng, S.; Liu, H.; Zhang, X.; Ye, W.; Han, Q.; Li, Y. Westerly jet stream controlled climate change mode since the Last Glacial Maximum in the northern Qinghai-Tibet Plateau. Earth Planet. Sci. Lett. 2020, 549, 116529. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, T.; Wu, B. Impact of developing ENSO on Tibetan Plateau summer rainfall. J. Clim. 2021, 34, 3385–3400. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.; Nakatsuka, T.; Fang, K.; Song, H.; Liu, R.; Wang, K. East Asian Summer Monsoon moisture sustains summer relative humidity in the southwestern Gobi Desert, China: Evidence from δ18O of tree rings. Clim. Dyn. 2019, 52, 6321–6337. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Lloyd, J. Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere. In Stable Isotopes and Plant Carbon-Water Relations; Academic Press: Cambridge, MA, USA, 1993; pp. 47–70. [Google Scholar]
- Mirfenderesgi, G.; Bohrer, G.; Matheny, A.M.; Fatichi, S.; Schäfer, K.V. Tree level hydrodynamic approach for resolving aboveground water storage and stomatal conductance and modeling the effects of tree hydraulic strategy. J. Geophys. Res. Biogeosci. 2016, 121, 1792–1813. [Google Scholar] [CrossRef]
- Zhan, N.; Wang, Z.; Xie, Y.; Shang, X.; Liu, G.; Wu, Z. Expression patterns and regulation of non-coding RNAs during synthesis of cellulose in Eucalyptus grandis Hill. Forests 2021, 12, 1565. [Google Scholar] [CrossRef]
- Giraldo, J.A.; Valle, J.I.; González-Caro, S.; Sierra, C.A. Intra-annual isotope variations in tree rings reveal growth rhythms within the least rainy season of an ever-wet tropical forest. Trees 2022, 36, 1039–1052. [Google Scholar] [CrossRef]
- Rao, W.; Chen, X.; Meredith, K.T.; Tan, H.; Gao, M.; Liu, J. Water uptake of riparian plants in the lower Lhasa River Basin, South Tibetan Plateau using stable water isotopes. Hydrol. Process. 2020, 34, 3492–3505. [Google Scholar] [CrossRef]
- Van, T.; Verheyen, K.; Kint, V.; Van, E.; Muys, B. Plasticity of tree architecture through interspecific and intraspecific competition in a young experimental plantation. For. Ecol. Manag. 2017, 385, 1–9. [Google Scholar]
- Schwendenmann, L.; Pendall, E.; Sanchez-Bragado, R.; Kunert, N.; Hölscher, D. Tree water uptake in a tropical plantation varying in tree diversity: Interspecific differences, seasonal shifts and complementarity. Ecohydrology 2015, 8, 1–12. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Alexander, M.R.; Xia, J.; Wang, F.; Zhang, F.; Pederson, N. Drought limits wood production of Juniperus przewalskii even as growing seasons lengthens in a cold and arid environment. Catena 2021, 196, 104936. [Google Scholar] [CrossRef]
- Brienen, R.J.; Hietz, P.; Wanek, W.; Gloor, M. Oxygen isotopes in tree rings record variation in precipitation δ18O and amount effects in the south of Mexico. J. Geophys. Res. Biogeosci. 2013, 118, 1604–1615. [Google Scholar] [CrossRef]
- Guo, X.; Tian, L. Spatial patterns and possible mechanisms of precipitation changes in recent decades over and around the Tibetan Plateau in the context of intense warming and weakening winds. Clim. Dyn. 2022, 59, 2081–2102. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Li, Q. Asian summer monsoon-related relative humidity recorded by tree ring δ18O during last 205 years. J. Geophys. Res. Atmos. 2019, 124, 9824–9838. [Google Scholar] [CrossRef]
- Li, F.F.; Lu, H.L.; Wang, G.Q.; Yao, Z.Y.; Li, Q.; Qiu, J. Zoning of precipitation regimes on the Qinghai-Tibet Plateau and its surrounding areas responded by the vegetation distribution. Sci. Total Environ. 2022, 838, 155844. [Google Scholar] [CrossRef]
- Holdo, R.M.; Nippert, J.B.; Mack, M.C. Rooting depth varies differentially in trees and grasses as a function of mean annual rainfall in an African savanna. Oecologia 2018, 186, 269–280. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, S.; Song, W.; Lu, Q.; Lin, G. Water-use strategies of two desert plants along a precipitation gradient in northwestern China. J. Plant Ecol. 2011, 35, 789. [Google Scholar] [CrossRef]
- Sun, L.; Yang, L.; Chen, L.; Zhao, F.; Li, S. Short-term changing patterns of stem water isotopes in shallow soils underlain by fractured bedrock. Hydrol. Res. 2019, 50, 577–588. [Google Scholar] [CrossRef]
- Grossiord, C.; Sevanto, S.; Dawson, T.E.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; McDowell, N.G. Warming combined with more extreme precipitation regimes modifies the water sources used by trees. New Phytol. 2017, 213, 584–596. [Google Scholar] [CrossRef]
- Oerter, E.; Finstad, K.; Schaefer, J.; Goldsmith, G.R.; Dawson, T.; Amundson, R. Oxygen isotope fractionation effects in soil water via interaction with cations (Mg, Ca, K, Na) adsorbed to phyllosilicate clay minerals. J. Hydrol. 2014, 515, 1–9. [Google Scholar] [CrossRef]
- Dai, Y.; Zheng, X.J.; Tang, L.S.; Li, Y. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Plant Soil 2015, 389, 73–87. [Google Scholar] [CrossRef]
- Tron, S.; Bodner, G.; Laio, F. Can diversity in root architecture explain plant water use efficiency? A modeling study. Ecol. Modell. 2015, 312, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Jiao, L.; Zhang, P.; Du, D.; Wu, X.; Wei, M.; Li, Q.; Wang, X.; Qi, C. The key role of ecological resilience in radial growth processes of conifers under drought stress in the subalpine zone of marginal deserts. Sci. Total Environ. 2023, 903, 166864. [Google Scholar] [CrossRef]
- Yao, Y.; Liu, Y.; Zhou, S.; Son, J.; Fu, B. Soil moisture determines the recovery time of ecosystems from drought. Glob. Change Biol. 2013, 29, 3562–3574. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
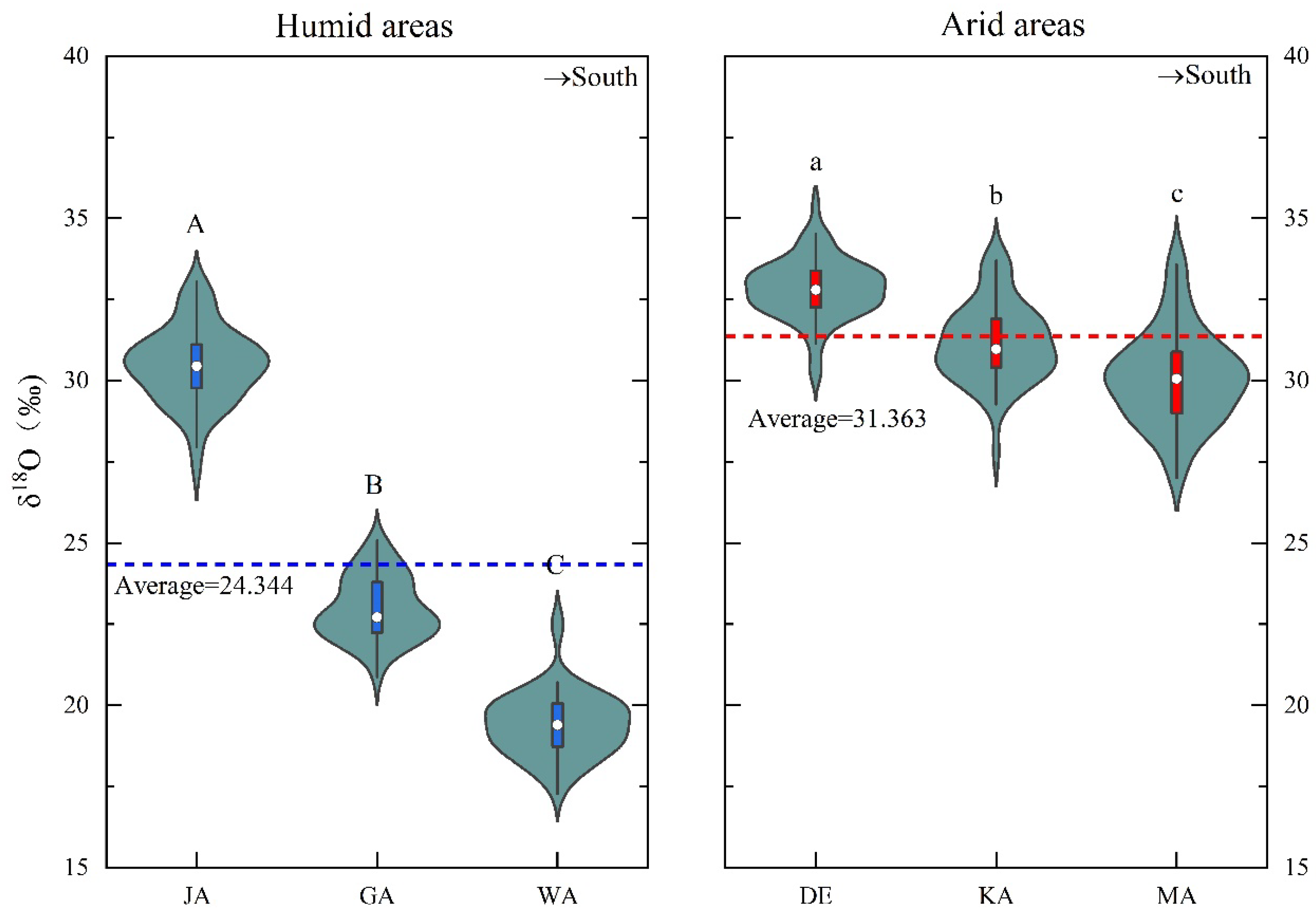{kind=link}
{kind=link}
{kind=link}
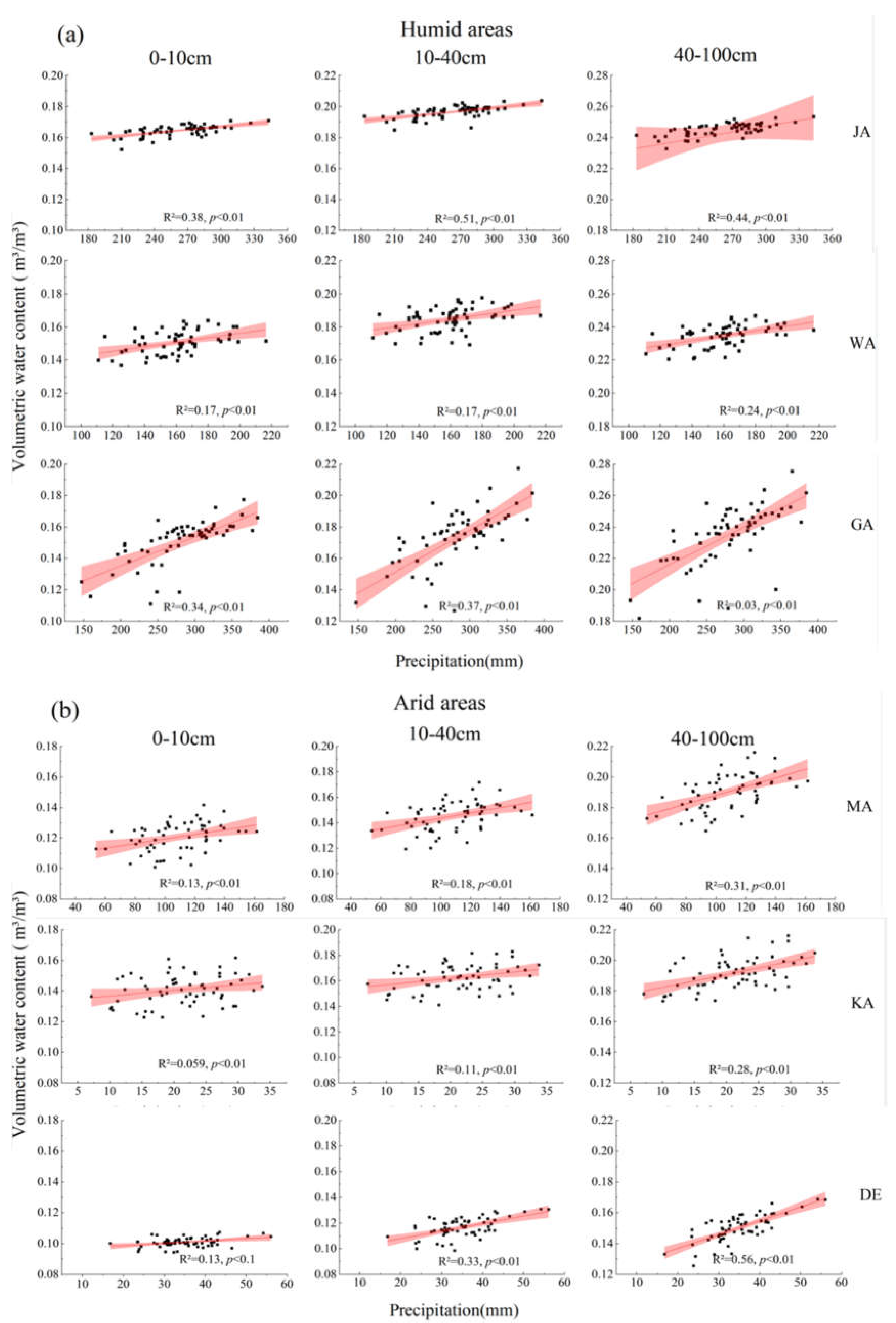{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Location | Elevation (m) | Tree Species | Time Span | Annual Total Precipitation (mm) | Date Source |
|---|---|---|---|---|---|---|
| JA | 29°38′ N, 79°51′ E | 1870 | Cedrus deodara | 1621–2008 | 1505 | Xu et al. (2018) [31] |
| WA | 27°59′ N, 90° E | 3500 | Larix griffithii | 1743–2011 | 1330 | Sano et al. (2013) [36] |
| GA | 28°10′ N, 85°11′ E | 3500 | Abies spectabilis | 1801–2000 | 958 | Xu et al. (2018) [31] |
| MA | 32°13′ N, 77°13′ E | 2700 | Abies pindrow | 1768–2008 | 650 | Sano et al. (2017) [32] |
| KA | 35°54′ N, 74°56′ E | 2900 | Juniperus excelsa | 1900–1998 | 329 | Treydte et al. (2006) [37] |
| DE | 37°48′ N, 97°78′ E | 3500 | Juniperus przewalskii | 1168–2011 | 197 | Yang et al. (2021) [38] |
| Control Variables | Correlations with δ18OTR | JA | WA | GA | MA | KA | DE |
|---|---|---|---|---|---|---|---|
| VPD | VMC0–10 | −0.396 ** | −0.364 * | −0.409 * | −0.459 * | −0.211 | −0.145 |
| VMC10–40 | −0.265 | −0.360 * | −0.383 * | −0.472 * | −0.277 | −0.286 | |
| VMC40–100 | −0.210 | −0.225 | −0.241 | −0.416 * | −0.356* | −0.385 * | |
| PDSI | −0.161 | −0.106 | −0.255 | −0.264 | −0.278 | −0.585 ** | |
| RH | 0.270 | 0.179 | 0.423 * | 0.151 | −0.064 | −0.163 | |
| PDSI | VMC0–10 | −0.251 | −0.459 * | −0.365 * | −0.527 ** | −0.004 | 0.173 |
| VMC10–40 | −0.213 | −0.451 * | −0.317 | −0.542 ** | −0.118 | 0.164 | |
| VMC40–100 | −0.14 | −0.156 | −0.343 | −0.500 ** | −0.239 | 0.119 | |
| VPD | 0.057 | 0.432 | −0.141 | −0.314 | 0.003 | −0.021 | |
| RH | 0.072 | −0.396 * | 0.251 | −0.028 | −0.042 | −0.022 | |
| RH | VMC0–10 | −0.356 ** | −0.377 * | −0.392 * | −0.53 ** | −0.213 | −0.130 |
| VMC10–40 | −0.289 | −0.380 * | −0.365 * | −0.542 ** | −0.276 | −0.274 | |
| VMC40–100 | −0.214 | −0.225 | −0.280 | −0.499 * | −0.352 * | −0.365 * | |
| VPD | 0.296 | 0.259 | 0.367 * | 0.159 | −0.042 | −0.008 | |
| PDSI | −0.208 | −0.113 | −0.242 | −0.315 | −0.277 | −0.570 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Jiao, L.; Xue, R.; Che, X.; Zhang, P.; Wang, X.; Yuan, X. Seasonal and Depth Dynamics of Soil Moisture Affect Trees on the Tibetan Plateau. Forests 2024, 15, 752. https://doi.org/10.3390/f15050752
Li Q, Jiao L, Xue R, Che X, Zhang P, Wang X, Yuan X. Seasonal and Depth Dynamics of Soil Moisture Affect Trees on the Tibetan Plateau. Forests. 2024; 15(5):752. https://doi.org/10.3390/f15050752
Chicago/Turabian StyleLi, Qian, Liang Jiao, Ruhong Xue, Xichen Che, Peng Zhang, Xuge Wang, and Xin Yuan. 2024. "Seasonal and Depth Dynamics of Soil Moisture Affect Trees on the Tibetan Plateau" Forests 15, no. 5: 752. https://doi.org/10.3390/f15050752